

Staying Dry and Clean: An Insect’s Guide to Hydrophobicity

Abstract
:Simple Summary
Abstract
1. Fundamentals of Wettability in Nature
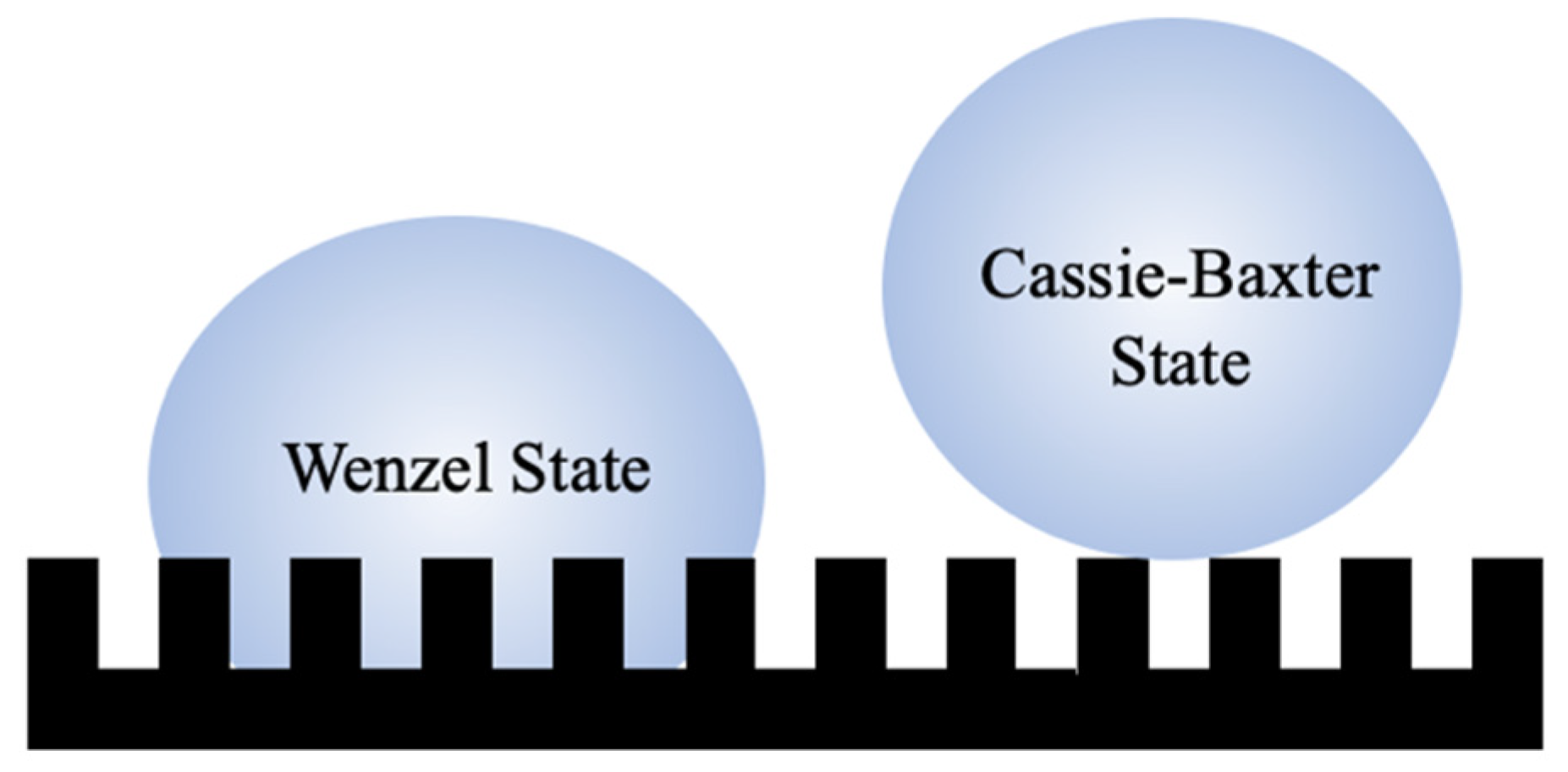
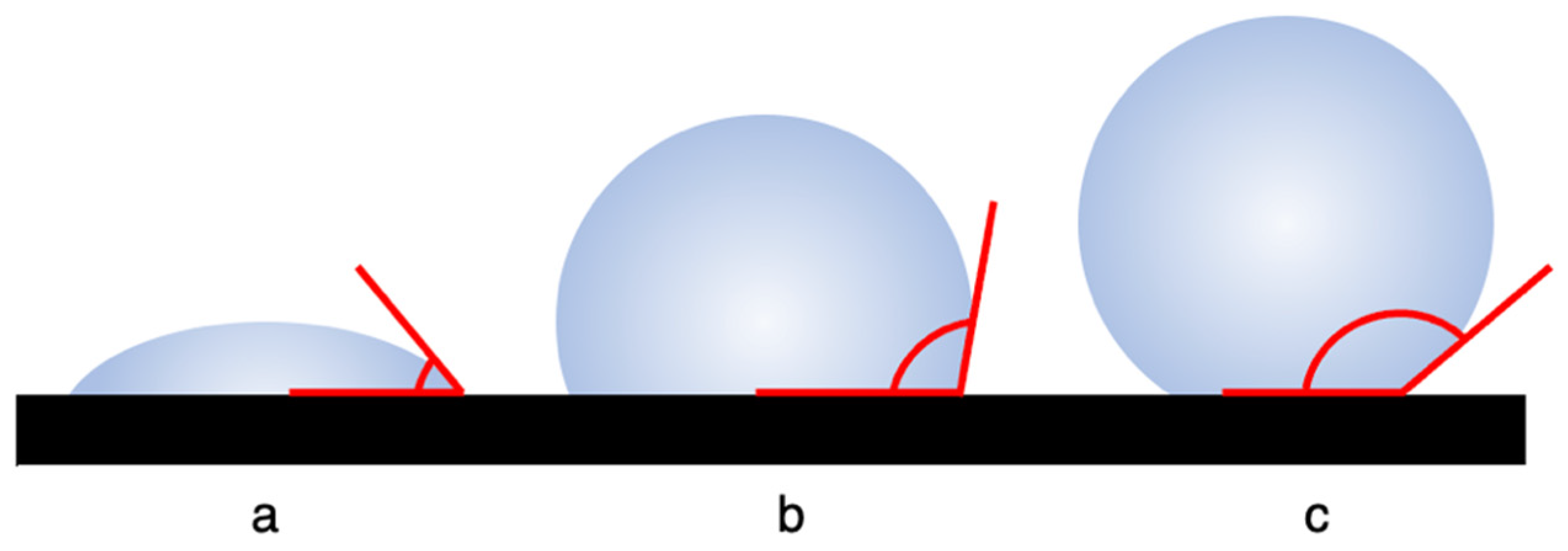
1.1. Wettability
1.2. Hydrophobic Mechanisms in Nature
1.3. Hydrophobic Mechanisms in Insects
2. Terminology and Methodology
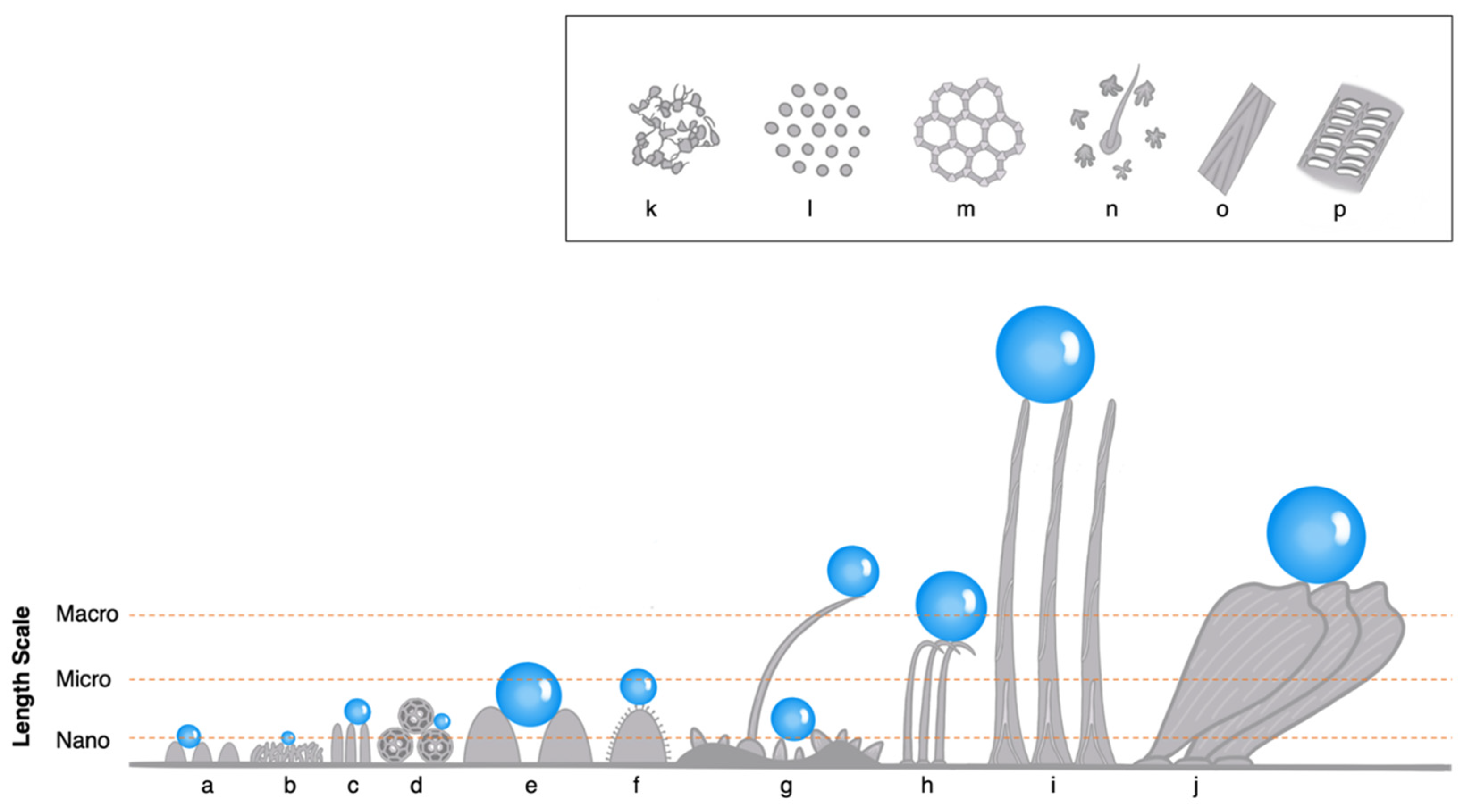
2.1. Overview of Length Scale and Cuticular Terminology
2.2. Literature Review Methodology
3. Hydrophobic Cuticular Structures Found in Insects
3.1. Nanoscale Hydrophobic Structures of Insects
3.2. Microscale Hydrophobic Structures of Insects
3.3. Macroscale Hydrophobic Structures of Insects
3.4. Hierarchical Hydrophobic Structures of Insects
3.4.1. Blattodea
3.4.2. Coleoptera
3.4.3. Collembola
3.4.4. Diptera
3.4.5. Hemiptera
3.4.6. Hymenoptera
3.4.7. Lepidoptera
3.4.8. Orthoptera
3.4.9. Neuroptera
3.4.10. Mecoptera, Megaloptera, and Trichoptera
4. Discussion and Bioinspired Design Implications
5. Outlook
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schroeder, T.B.H.; Houghtaling, J.; Wilts, B.D.; Mayer, M. It’s Not a Bug, It’s a Feature: Functional Materials in Insects. Adv. Mater. 2018, 30, 1705322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishino, T.; Meguro, M.; Nakamae, K.; Matsushita, M.; Ueda, Y. The Lowest Surface Free Energy Based on −CF 3 Alignment. Langmuir 1999, 15, 4321–4323. [Google Scholar] [CrossRef]
- Bico, J.; Marzolin, C.; Quéré, D. Pearl Drops. EPL 1999, 47, 220. [Google Scholar] [CrossRef]
- Herminghaus, S. Roughness-Induced Non-Wetting. EPL 2000, 52, 165. [Google Scholar] [CrossRef]
- Lafuma, A.; Quéré, D. Superhydrophobic States. Nat. Mater. 2003, 2, 457–460. [Google Scholar] [CrossRef]
- Yoshimitsu, Z.; Nakajima, A.; Watanabe, T.; Hashimoto, K. Effects of Surface Structure on the Hydrophobicity and Sliding Behavior of Water Droplets. Langmuir 2002, 18, 5818–5822. [Google Scholar] [CrossRef]
- Cassie, A.B.D.; Baxter, S. Wettability of Porous Surfaces. Trans. Faraday Soc. 1944, 40, 546–551. [Google Scholar] [CrossRef]
- Wenzel, R.N. Resistance of Solid Surfaces to Wetting by Water. Ind. Eng. Chem. 1936, 28, 988–994. [Google Scholar] [CrossRef]
- Bhushan, B.; Nosonovsky, M. The Rose Petal Effect and the Modes of Superhydrophobicity. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2010, 368, 4713–4728. [Google Scholar] [CrossRef]
- Bico, J.; Thiele, U.; Quéré, D. Wetting of Textured Surfaces. Colloids Surf. A Physicochem. Eng. Asp. 2002, 206, 41–46. [Google Scholar] [CrossRef]
- Quéré, D. Wetting and Roughness. Annu. Rev. Mater. Res. 2008, 38, 71–99. [Google Scholar] [CrossRef]
- Barthlott, W.; Mail, M.; Bhushan, B.; Koch, K. Plant Surfaces: Structures and Functions for Biomimetic Innovations. Nano-Micro Lett. 2017, 9, 1265–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimoto, S.; Bhushan, B. Bioinspired Self-Cleaning Surfaces with Superhydrophobicity, Superoleophobicity, and Superhydrophilicity. RSC Adv. 2012, 3, 671–690. [Google Scholar] [CrossRef]
- Watson, G.S.; Green, D.W.; Cribb, B.W.; Brown, C.L.; Meritt, C.R.; Tobin, M.J.; Vongsvivut, J.; Sun, M.; Liang, A.-P.; Watson, J.A. Insect Analogue to the Lotus Leaf: A Planthopper Wing Membrane Incorporating a Low-Adhesion, Nonwetting, Superhydrophobic, Bactericidal, and Biocompatible Surface. ACS Appl. Mater. Interfaces 2017, 9, 24381–24392. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Liu, W. Biomimic from the Superhydrophobic Plant Leaves in Nature: Binary Structure and Unitary Structure. Plant Sci. 2007, 172, 1103–1112. [Google Scholar] [CrossRef]
- Hsu, C.-P.; Lin, Y.-M.; Chen, P.-Y. Hierarchical Structure and Multifunctional Surface Properties of Carnivorous Pitcher Plants Nepenthes. JOM 2015, 67, 744–753. [Google Scholar] [CrossRef]
- Li, Z.; Guo, Z. Bioinspired Surfaces with Wettability for Antifouling Application. Nanoscale 2019, 11, 22636–22663. [Google Scholar] [CrossRef]
- Wong, T.-S.; Kang, S.H.; Tang, S.K.Y.; Smythe, E.J.; Hatton, B.D.; Grinthal, A.; Aizenberg, J. Bioinspired Self-Repairing Slippery Surfaces with Pressure-Stable Omniphobicity. Nature 2011, 477, 443–447. [Google Scholar] [CrossRef]
- Bormashenko, E.; Bormashenko, Y.; Stein, T.; Whyman, G.; Bormashenko, E. Why do pigeon feathers repel water? Hydrophobicity of pennae, Cassie–Baxter wetting hypothesis and Cassie–Wenzel capillarity-induced wetting transition. J. Colloid Interf. Sci. 2007, 311, 212–216. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, X.; Xin, J.H. Hydrophobic duck feathers and their simulation on textile substrates for water repellent treatment. Bioinspir. Biomim. 2008, 3, 046007. [Google Scholar] [CrossRef]
- Darmanin, T.; Guittard, F. Superhydrophobic and Superoleophobic Properties in Nature. Mater. Today 2015, 18, 273–285. [Google Scholar] [CrossRef]
- Böcher, J.; Nachman, G. Temperature and Humidity Responses of the Arctic-Alpine Seed Bug Nysius Groenlandicus. Entomol. Exp. Appl. 2001, 99, 319–330. [Google Scholar] [CrossRef]
- Worland, M.R.; Block, W.L.; Grubor-Lajsic, G.O. Survival of Heleomyza Borealis (Diptera, Heleomyzidae) Larvae down to −60 °C. Physiol. Entomol. 2000, 25, 1–5. [Google Scholar] [CrossRef]
- Bartholomew, A.; El Moghrabi, J. Seasonal Preference of Darkling Beetles (Tenebrionidae) for Shrub Vegetation Due to High Temperatures, Not Predation or Food Availability. J. Arid Environ. 2018, 156, 34–40. [Google Scholar] [CrossRef]
- Raś, M.; Kamiński, M.J.; Iwan, D. Fossoriality in Desert-Adapted Tenebrionid (Coleoptera) Larvae. Sci. Rep. 2022, 12, 13233. [Google Scholar] [CrossRef]
- Birrell, J.H.; Shah, A.A.; Hotaling, S.; Giersch, J.J.; Williamson, C.E.; Jacobsen, D.; Woods, H.A. Insects in High-Elevation Streams: Life in Extreme Environments Imperiled by Climate Change. Glob. Chang. Biol. 2020, 26, 6667–6684. [Google Scholar] [CrossRef]
- Kuitunen, K.; Kovalev, A.; Gorb, S.N. Sex-Related Effects in the Superhydrophobic Properties of Damselfly Wings in Young and Old Calopteryx Splendens. PLoS ONE 2014, 9, e88627. [Google Scholar] [CrossRef] [Green Version]
- Watson, G.S.; Watson, J.A.; Cribb, B.W. Diversity of Cuticular Micro- and Nanostructures on Insects: Properties, Functions, and Potential Applications. Annu. Rev. Entomol. 2017, 62, 185–205. [Google Scholar] [CrossRef] [Green Version]
- Andersen, S.O.; Peter, M.G.; Roepstorff, P. Cuticular Sclerotization in Insects. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1996, 113, 689–705. [Google Scholar] [CrossRef]
- Andersen, S.O. Insect Cuticular Sclerotization: A Review. Insect Biochem. Mol. Biol. 2010, 40, 166–178. [Google Scholar] [CrossRef]
- Hopkins, T.L.; Kramer, K.J. Insect Cuticle Sclerotization. Annu. Rev. Entomol. 1992, 37, 273–302. [Google Scholar] [CrossRef]
- Gorb, S. Attachment Devices of Insect Cuticle; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; ISBN 978-0-7923-7153-3. [Google Scholar]
- Wei, L.; Reiter, K.E.; McElrath, T.; Alleyne, M.; Dunn, A.C. Diffraction Gratings Alter the Surface Friction of Iridescent Beetle Cuticle against Fibrous Surfaces. Biotribology 2019, 20, 100108. [Google Scholar] [CrossRef]
- Jaggessar, A.; Shahali, H.; Mathew, A.; Yarlagadda, P.K.D.V. Bio-Mimicking Nano and Micro-Structured Surface Fabrication for Antibacterial Properties in Medical Implants. J. Nanobiotechnol. 2017, 15, 64. [Google Scholar] [CrossRef] [Green Version]
- Hinton, H.E. Plastron Respiration in Bugs and Beetles. J. Insect Physiol. 1976, 22, 1529–1550. [Google Scholar] [CrossRef]
- Wan, Q.; Li, H.; Zhang, S.; Wang, C.; Su, S.; Long, S.; Pan, B. Combination of Active Behaviors and Passive Structures Contributes to the Cleanliness of Housefly Wing Surfaces: A New Insight for the Design of Cleaning Materials. Colloids Surf. B Biointerfaces 2019, 180, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Ghiradella, H.; Radigan, W. Collembolan Cuticle: Wax Layer and Antiwetting Properties. J. Insect Physiol. 1974, 20, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Ghiradella, H. Chapter 4—Insect Cuticular Surface Modifications: Scales and Other Structural Formations. In Advances in Insect Physiology; Casas, J., Simpson, S.J., Eds.; Advances in Insect Physiology: Insect Integument and Colour; Academic Press: Cambridge, MA, USA, 2010; Volume 38, pp. 135–180. [Google Scholar]
- Gao, X.; Jiang, L. Water-Repellent Legs of Water Striders. Nature 2004, 432, 36. [Google Scholar] [CrossRef]
- Mail, M.; Klein, A.; Bleckmann, H.; Schmitz, A.; Scherer, T.; Rühr, P.T.; Lovric, G.; Fröhlingsdorf, R.; Gorb, S.N.; Barthlott, W. A New Bioinspired Method for Pressure and Flow Sensing Based on the Underwater Air-Retaining Surface of the Backswimmer Notonecta. Beilstein J. Nanotechnol. 2018, 9, 3039–3047. [Google Scholar] [CrossRef] [Green Version]
- Parker, A.R.; Lawrence, C.R. Water Capture by a Desert Beetle. Nature 2001, 414, 33–34. [Google Scholar] [CrossRef]
- Enright, R.; Miljkovic, N.; Alvarado, J.L.; Kim, K.; Rose, J.W. Dropwise Condensation on Micro- and Nanostructured Surfaces. Nanoscale Microscale Thermophys. Eng. 2014, 18, 223–250. [Google Scholar] [CrossRef]
- Miljkovic, N.; Enright, R.; Wang, E.N. Modeling and Optimization of Superhydrophobic Condensation. J. Heat Transf. 2013, 135, 111004. [Google Scholar] [CrossRef]
- Zhao, D.; Tian, Q.; Wang, M.; Jin, Y. Study on the Hydrophobic Property of Shark-Skin-Inspired Micro-Riblets. J. Bionic Eng. 2014, 11, 296–302. [Google Scholar] [CrossRef]
- Bharathidasan, T.; Kumar, S.V.; Bobji, M.S.; Chakradhar, R.P.S.; Basu, B.J. Effect of Wettability and Surface Roughness on Ice-Adhesion Strength of Hydrophilic, Hydrophobic and Superhydrophobic Surfaces. Appl. Surf. Sci. 2014, 314, 241–250. [Google Scholar] [CrossRef]
- Koszut, J.; Boyina, K.; Popovic, G.; Carpenter, J.; Wang, S.; Miljkovic, N. Superhydrophobic Heat Exchangers Delay Frost Formation and Reduce Defrost Energy Input of Aircraft Environmental Control Systems. Int. J. Heat Mass Transf. 2022, 189, 122669. [Google Scholar] [CrossRef]
- Mei, H.; Luo, D.; Guo, P.; Song, C.; Liu, C.; Zheng, Y.; Jiang, L. Multi-Level Micro-/Nanostructures of Butterfly Wings Adapt at Low Temperature to Water Repellency. Soft Matter 2011, 7, 10569–10573. [Google Scholar] [CrossRef]
- Nguyen, S.H.; Webb, H.K.; Mahon, P.J.; Crawford, R.J.; Ivanova, E.P. Natural Insect and Plant Micro-/Nanostructsured Surfaces: An Excellent Selection of Valuable Templates with Superhydrophobic and Self-Cleaning Properties. Molecules 2014, 19, 13614–13630. [Google Scholar] [CrossRef] [Green Version]
- Deparis, O.; Mouchet, S.; Dellieu, L.; Colomer, J.-F.; Sarrazin, M. Nanostructured Surfaces: Bioinspiration for Transparency, Coloration and Wettability. Mater. Today Proc. 2014, 1, 122–129. [Google Scholar] [CrossRef]
- Richards, A.G.; Richards, P.A. The Cuticular Protuberances of Insects. Int. J. Insect Morphol. Embryol. 1979, 8, 143–157. [Google Scholar] [CrossRef]
- Nguyen, S.H.; Webb, H.K.; Crawford, R.J.; Ivanova, E.P. Natural Antibacterial Surfaces. In Antibacterial Surfaces; Springer: Cham, Switzerland, 2015; pp. 9–26. ISBN 978-3-319-18594-1. [Google Scholar]
- Watson, G.S.; Cribb, B.W.; Watson, J.A. How Micro/Nanoarchitecture Facilitates Anti-Wetting: An Elegant Hierarchical Design on the Termite Wing. ACS Nano 2010, 4, 129–136. [Google Scholar] [CrossRef]
- Hasan, J.; Roy, A.; Chatterjee, K.; Yarlagadda, P.K.D.V. Mimicking Insect Wings: The Roadmap to Bioinspiration. ACS Biomater. Sci. Eng. 2019, 5, 3139–3160. [Google Scholar] [CrossRef]
- Byun, D.; Hong, J.; Saputra; Ko, J.H.; Lee, Y.J.; Park, H.C.; Byun, B.-K.; Lukes, J.R. Wetting Characteristics of Insect Wing Surfaces. J. Bionic Eng. 2009, 6, 63–70. [Google Scholar] [CrossRef]
- Wagner, T.; Neinhuis, C.; Barthlott, W. Wettability and Contaminability of Insect Wings as a Function of Their Surface Sculptures. Acta Zool. 1996, 77, 213–225. [Google Scholar] [CrossRef]
- Xu, Z.; Kong, L. A Study of Functional Micro/Nano Structural Surfaces in Bionic Applications. In Proceedings of the 2017 IEEE International Conference on Manipulation, Manufacturing and Measurement on the Nanoscale (3M-NANO), Shanghai, China, 7–11 August 2017; pp. 206–209. [Google Scholar]
- Hosoda, N.; Gorb, S.N. Underwater Locomotion in a Terrestrial Beetle: Combination of Surface de-Wetting and Capillary Forces. Proc. R. Soc. B Biol. Sci. 2012, 279, 4236–4242. [Google Scholar] [CrossRef] [Green Version]
- Balmert, A.; Bohn, H.F.; Ditsche-Kuru, P.; Barthlott, W. Dry under Water: Comparative Morphology and Functional Aspects of Air-Retaining Insect Surfaces. J. Morphol. 2011, 272, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, W.J.; Seely, M.K. Fog Basking by the Namib Desert Beetle, Onymacris Unguicularis. Nature 1976, 262, 284–285. [Google Scholar] [CrossRef]
- Sun, M.; Chen, Y.; Zheng, Y.; Zhen, M.; Shu, C.; Dai, Z.; Liang, A.; Gorb, S.N. Wettability Gradient on the Elytra in the Aquatic Beetle Cybister Chinensis and Its Role in Angular Position of the Beetle at Water-Air Interface. Acta Biomater. 2017, 51, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Montreuil, O.; Candet, C.; Bonaccorso, A.; Szczepanski, C.R.; Orange, F.; Godeau, R.-P.; Guittard, F.; Darmanin, T.; Godeau, G. Micro- and Nanoscopic Observations of Sexual Dimorphisms in Mecynorhina Polyphemus Confluens (Kraatz, 1890) (Coleoptera, Cetoniidae, Goliathini) and Consequences for Surface Wettability. Arthropod Struct. Dev. 2019, 49, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Filippov, A.E.; Kovalev, A.; Gorb, S.N. Numerical Simulation of the Pattern Formation of the Springtail Cuticle Nanostructures. J. R. Soc. Interface 2018, 15, 20180217. [Google Scholar] [CrossRef] [Green Version]
- Schmüser, L.; Zhang, W.; Marx, M.T.; Encinas, N.; Vollmer, D.; Gorb, S.; Baio, J.E.; Räder, H.J.; Weidner, T. Role of Surface Chemistry in the Superhydrophobicity of the Springtail Orchesella Cincta (Insecta:Collembola). ACS Appl. Mater. Interfaces 2020, 12, 12294–12304. [Google Scholar] [CrossRef]
- Gundersen, H.; Leinaas, H.P.; Thaulow, C. Surface structure and wetting characteristics of Collembola cuticles. PLoS ONE 2014, 9, e86783. [Google Scholar] [CrossRef]
- Hensel, R.; Neinhuis, C.; Werner, C. The springtail cuticle as a blueprint for omniphobic surfaces. Chem. Soc. Rev. 2016, 45, 323–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.-M.S.; Watson, G.S.; Cribb, B.W.; Watson, J.A. Non-Wetting Wings and Legs of the Cranefly Aided by Fine Structures of the Cuticle. J. Exp. Biol. 2011, 214, 915–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Monge, A.; Rodríguez Arrieta, J.; Jiménez-Chavarría, M.; Retana-Salazar, A. Observations on the Ultrastructure and Hydrophobicity of the Wings of Thirteen Neotropical Families of Diptera (Insecta) with Comments on Their Flight. Acta Microsc. 2015, 24, 111–117. [Google Scholar]
- Liu, K.; Jiang, L. Bio-Inspired Design of Multiscale Structures for Function Integration. Nano Today 2011, 6, 155–175. [Google Scholar] [CrossRef]
- Gao, X.; Yan, X.; Yao, X.; Xu, L.; Zhang, K.; Zhang, J.; Yang, B.; Jiang, L. The Dry-Style Antifogging Properties of Mosquito Compound Eyes and Artificial Analogues Prepared by Soft Lithography. Adv. Mater. 2007, 19, 2213–2217. [Google Scholar] [CrossRef]
- Sun, Z.; Liao, T.; Liu, K.; Jiang, L.; Kim, J.H.; Dou, S.X. Fly-Eye Inspired Superhydrophobic Anti-Fogging Inorganic Nanostructures. Small 2014, 10, 3001–3006. [Google Scholar] [CrossRef]
- Lee, S.C.; Kim, J.H.; Lee, S.J. Floating of the Lobes of Mosquito (Aedes Togoi) Larva for Respiration. Sci. Rep. 2017, 7, 43050. [Google Scholar] [CrossRef] [Green Version]
- Van Breugel, F.; Dickinson, M.H.; Meinwald, J. Superhydrophobic Diving Flies (Ephydra Hians) and the Hypersaline Waters of Mono Lake. Proc. Natl. Acad. Sci. USA 2017, 114, 13483–13488. [Google Scholar] [CrossRef] [Green Version]
- Neumann, D.; Woermann, D. Physical Conditions for Trapping Air by a Microtrichia-Covered Insect Cuticle during Temporary Submersion. Naturwissenschaften 2009, 96, 933–941. [Google Scholar] [CrossRef]
- Goodwyn, P.P.; Fujisaki, K.; De Souza, E.; Gorb, S. Moulding technique demonstrates the contribution of surface geometry to the super-hydrophobic properties of the surface of a water strider. Acta Biomater. 2008, 4, 766–770. [Google Scholar] [CrossRef]
- Orozco, F.; Alfaro-González, B.; Ureña, Y.C.; Villalobos, K.; Sanchez, A.; Bravo, F.; Vega, J.R.; Argüello-Miranda, O. Nanobiodiversity: The Potential of Extracellular Nanostructures. J. Renew. Mater. 2017, 5, 199–207. [Google Scholar] [CrossRef]
- Tripathy, A.; Sreedharan, S.; Bhaskarla, C.; Majumdar, S.; Peneti, S.K.; Nandi, D.; Sen, P. Enhancing the Bactericidal Efficacy of Nanostructured Multifunctional Surface Using an Ultrathin Metal Coating. Langmuir 2017, 33, 12569–12579. [Google Scholar] [CrossRef] [PubMed]
- Webb, H.K.; Hasan, J.; Truong, V.K.; Crawford, R.J.; Ivanova, E.P. Nature Inspired Structured Surfaces for Biomedical Applications. Curr. Med. Chem. 2011, 18, 3367–3375. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Liang, A.; Watson, G.S.; Watson, J.A.; Zheng, Y.; Ju, J.; Jiang, L. Influence of Cuticle Nanostructuring on the Wetting Behaviour/States on Cicada Wings. PLoS ONE 2012, 7, e35056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisdom, K.M.; Watson, J.A.; Qu, X.; Liu, F.; Watson, G.S.; Chen, C.-H. Self-Cleaning of Superhydrophobic Surfaces by Self-Propelled Jumping Condensate. Proc. Natl. Acad. Sci. USA 2013, 110, 7992–7997. [Google Scholar] [CrossRef] [Green Version]
- Dellieu, L.; Sarrazin, M.; Simonis, P.; Deparis, O.; Vigneron, J.P. A two-in-one superhydrophobic and anti-reflective nanodevice in the grey cicada Cicada orni (Hemiptera). J. Appl. Phys. 2014, 116, 024701. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.-M.; Watson, J.A.; Cribb, B.W.; Watson, G.S. Fouling of Nanostructured Insect Cuticle: Adhesion of Natural and Artificial Contaminants. Biofouling 2011, 27, 1125–1137. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.; Dana, C.E.; Hong, S.; Román, J.K.; Jo, K.D.; Hong, J.W.; Nguyen, J.; Cropek, D.M.; Alleyne, M.; Miljkovic, N. Exploring the Role of Habitat on the Wettability of Cicada Wings. ACS Appl. Mater. Interfaces 2017, 9, 27173–27184. [Google Scholar] [CrossRef]
- Oh, J.; Hoffman, J.B.; Hong, S.; Jo, K.D.; Román-Kustas, J.; Reed, J.H.; Dana, C.E.; Cropek, D.M.; Alleyne, M.; Miljkovic, N. Dissolvable Template Nanoimprint Lithography: A Facile and Versatile Nanoscale Replication Technique. Nano Lett. 2020, 20, 6989–6997. [Google Scholar] [CrossRef]
- Román, J.K.; Walsh, C.M.; Oh, J.; Dana, C.E.; Hong, S.; Jo, K.D.; Alleyne, M.; Miljkovic, N.; Cropek, D.M. Spatially Resolved Chemical Analysis of Cicada Wings Using Laser-Ablation Electrospray Ionization (LAESI) Imaging Mass Spectrometry (IMS). Anal. Bioanal. Chem. 2018, 410, 1911–1921. [Google Scholar] [CrossRef]
- Román-Kustas, J.; Hoffman, J.B.; Reed, J.H.; Gonsalves, A.E.; Oh, J.; Li, L.; Hong, S.; Jo, K.D.; Dana, C.E.; Miljkovic, N.; et al. Molecular and Topographical Organization: Influence on Cicada Wing Wettability and Bactericidal Properties. Adv. Mater. Interfaces 2020, 7, 2000112. [Google Scholar] [CrossRef]
- Ditsche-Kuru, P.; Schneider, E.S.; Melskotte, J.-E.; Brede, M.; Leder, A.; Barthlott, W. Superhydrophobic Surfaces of the Water Bug Notonecta Glauca: A Model for Friction Reduction and Air Retention. Beilstein J. Nanotechnol. 2011, 2, 137–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seymour, R.S.; Matthews, P.G.D. Physical Gills in Diving Insects and Spiders: Theory and Experiment. J. Exp. Biol. 2013, 216, 164–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynn, M.R.; Bush, J.W.M. Underwater Breathing: The Mechanics of Plastron Respiration. J. Fluid Mech. 2008, 608, 275–296. [Google Scholar] [CrossRef]
- Rakitov, R.; Gorb, S.N. Brochosomal Coats Turn Leafhopper (Insecta, Hemiptera, Cicadellidae) Integument to Superhydrophobic State. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakitov, R.A. Brochosomal Coatings of the Integument of Leafhoppers (Hemiptera, Cicadellidae). In Functional Surfaces in Biology; Springer: Dordrecht, The Netherlands, 2009; Volume 1, pp. 113–137. ISBN 978-1-4020-6694-8. [Google Scholar]
- Pike, N.; Richard, D.; Foster, W.; Mahadevan, L. How Aphids Lose Their Marbles. Proc. Biol. Sci. 2002, 269, 1211–1215. [Google Scholar] [CrossRef] [Green Version]
- Boevé, J.-L.; Voigt, D.; Gorb, S.N. Integument and Defence in Larva and Prepupa of a Sawfly Living on a Semi-Aquatic Plant. Naturwissenschaften 2013, 100, 107–110. [Google Scholar] [CrossRef]
- Voigt, D.; Gorb, S.; Boevé, J.-L. Superhydrophobic Cuticle with a “ Pinning Effect” in the Larvae of the Iris Sawfly, Rhadinoceraea Micans (Hymenoptera, Tenthredinidae). Zoology 2011, 114, 265–271. [Google Scholar] [CrossRef]
- Wei, J.; Liang, Y.; Chen, X.; Gorb, S.N.; Wu, Z.; Li, H.; Wu, J. Enhanced Flexibility of the Segmented Honey Bee Tongue with Hydrophobic Tongue Hairs. ACS Appl. Mater. Interfaces 2022, 14, 12911–12919. [Google Scholar] [CrossRef]
- Rodriguez, L.J.; Young, F.; Rasplus, J.-Y.; Kjellberg, F.; Compton, S.G. Constraints on Convergence: Hydrophobic Hind Legs Allow Some Male Pollinator Fig Wasps Early Access to Submerged Females. J. Nat. Hist. 2017, 51, 761–782. [Google Scholar] [CrossRef] [Green Version]
- Mlot, N.J.; Tovey, C.A.; Hu, D.L. Fire Ants Self-Assemble into Waterproof Rafts to Survive Floods. Proc. Natl. Acad. Sci. USA 2011, 108, 7669–7673. [Google Scholar] [CrossRef] [PubMed]
- Kovalev, A.; Rebora, M.; Salerno, G.; Gorb, S. Air-Entrapping Capacity in the Hair Coverage of Malacosoma Castrensis (Lasiocampidae: Lepidoptera) Caterpillar: A Case Study. J. Exp. Biol. 2020, 223, jeb225029. [Google Scholar] [CrossRef]
- Sackey, J.; Sone, B.T.; Dompreh, K.A.; Maaza, M. Wettability Property in Natural Systems: A Case of Flying Insects. MRS Adv. 2018, 3, 2697–2703. [Google Scholar] [CrossRef]
- Sato, O.; Kubo, S.; Gu, Z.Z. Structural Color Films with Lotus Effects, Superhydrophilicity, and Tunable Stop-Bands. Acc. Chem. Res. 2009, 42, 1–10. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B. Depilation Increases While Hairiness Decreases the Risk of Drowning: A Hitherto Unappreciated Survival Role of Setae in Woolly Bear Caterpillars of the Moth Lemyra Imparilis (Lepidoptera: Noctuoidea: Erebidae). Eur. J. Entomol. 2016, 113, 130–134. [Google Scholar] [CrossRef] [Green Version]
- Cheeseman, S.; Owen, S.; Truong, V.K.; Meyer, D.; Ng, S.H.; Vongsvivut, J.; Linklater, D.; Tobin, M.J.; Werner, M.; Baulin, V.A.; et al. Pillars of Life: Is There a Relationship between Lifestyle Factors and the Surface Characteristics of Dragonfly Wings? ACS Omega 2018, 3, 6039–6046. [Google Scholar] [CrossRef]
- Nguyen, S.H.T.; Webb, H.K.; Hasan, J.; Tobin, M.J.; Crawford, R.J.; Ivanova, E.P. Dual Role of Outer Epicuticular Lipids in Determining the Wettability of Dragonfly Wings. Colloids Surf. B Biointerfaces 2013, 106, 126–134. [Google Scholar] [CrossRef]
- Gao, C.-Y.; Meng, G.-X.; Li, X.; Wu, M.; Liu, Y.; Li, X.-Y.; Zhao, X.; Lee, I.; Feng, X. Wettability of Dragonfly Wings: The Structure Detection and Theoretical Modeling: Wettability, Modeling, and Simulation of Dragonfly Wings. Surf. Interface Anal. 2013, 45, 650–655. [Google Scholar] [CrossRef]
- Wan, Y.; Cong, Q.; Wang, X.; Yan, Z. The Wettability and Mechanism of Geometric Non-Smooth Structure of Dragonfly Wing Surface. J. Bionic Eng. 2008, 5, 40–45. [Google Scholar] [CrossRef]
- Aideo, S.N.; Mohanta, D. Limiting hydrophobic behavior and reflectance response of dragonfly and damselfly wings. Appl. Surf. Sci. 2016, 387, 609–616. [Google Scholar] [CrossRef]
- Gorb, S.N.; Tynkkynen, K.; Kotiaho, J.S. Crystalline wax coverage of the imaginal cuticle in Calopteryx splendens (Odonata: Calopterygidae). Inter. J. Odonatol. 2009, 12, 205–221. [Google Scholar] [CrossRef]
- Watson, G.S.; Cribb, B.W.; Watson, J.A. The Role of Micro/Nano Channel Structuring in Repelling Water on Cuticle Arrays of the Lacewing. J. Struct. Biol. 2010, 171, 44–51. [Google Scholar] [CrossRef]
- Watson, G.S.; Myhra, S.; Cribb, B.W.; Watson, J.A. Putative Functions and Functional Efficiency of Ordered Cuticular Nanoarrays on Insect Wings. Biophys. J. 2008, 94, 3352–3360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakitov, R.A. Powdering of Egg Nests with Brochosomes and Related Sexual Dimorphism in Leafhoppers (Hemiptera: Cicadellidae). Zool. J. Linn. Soc. 2004, 140, 353–381. [Google Scholar] [CrossRef] [Green Version]
- Rakitov, R. The Covering Formed by Brochosomes on the Cuticle of Leafhoppers (Homoptera, Cicadellidae). Entomol. Rev. 1995, 74, 90–103. [Google Scholar]
- Watson, G.S.; Cribb, B.W.; Watson, J.A. Contrasting Micro/Nano Architecture on Termite Wings: Two Divergent Strategies for Optimising Success of Colonisation Flights. PLoS ONE 2011, 6, e24368. [Google Scholar] [CrossRef] [Green Version]
- Gould, J.; Valdez, J.W. Locomotion with a Twist: Aquatic Beetle Walks Upside down on the Underside of the Water’s Surface. Ethology 2021, 127, 669–673. [Google Scholar] [CrossRef]
- Keilin, D.; Tate, P.; Vincent, M. The Perispiracular Glands of Mosquito Larvae. Parasitology 1935, 27, 257–262. [Google Scholar] [CrossRef]
- Park, K.-C.; Kim, P.; Grinthal, A.; He, N.; Fox, D.; Weaver, J.C.; Aizenberg, J. Condensation on Slippery Asymmetric Bumps. Nature 2016, 531, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Nowlin, K.; Lajeunesse, D.R. Fabrication of Hierarchical Biomimetic Polymeric Nanostructured Surfaces. Mol. Syst. Des. Eng. 2017, 2, 201–213. [Google Scholar] [CrossRef]
- Agonafer, D.D.; Lee, H.; Vasquez, P.A.; Won, Y.; Jung, K.W.; Lingamneni, S.; Ma, B.; Shan, L.; Shuai, S.; Du, Z.; et al. Porous Micropillar Structures for Retaining Low Surface Tension Liquids. J. Colloid Interface Sci. 2018, 514, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Bhadra, C.M.; Dang, T.H.Y.; Buividas, R.; Wang, J.; Crawford, R.J.; Ivanova, E.P.; Juodkazis, S. A Bactericidal Microfluidic Device Constructed Using Nano-Textured Black Silicon. RSC Adv. 2016, 6, 26300–26306. [Google Scholar] [CrossRef]
- Sharma, D.K.; Baghel, V.; Kumar, R.; Avasthi, D.K.; Sikarwar, B.S. Recent Developments in Fabrication of Super-Hydrophobic Surfaces: A Review. Lect. Notes Mech. Eng. 2019, 127–140. [Google Scholar] [CrossRef]
- Khodakarami, S.; Fazle Rabbi, K.; Suh, Y.; Won, Y.; Miljkovic, N. Machine Learning Enabled Condensation Heat Transfer Measurement. Int. J. Heat Mass Transf. 2022, 194, 123016. [Google Scholar] [CrossRef]
- Kwon, B.; Foulkes, T.; Yang, T.; Miljkovic, N.; King, W.P. Additively Manufactured Impinging Air Jet Cooler for High-Power Electronic Devices. In Proceedings of the 2019 18th IEEE Intersociety Conference on Thermal and Thermomechanical Phenomena in Electronic Systems (ITherm), Las Vegas, NV, USA, 28–31 May 2019; pp. 941–945. [Google Scholar]
- Kim, S.; Wu, Z.; Esmaili, E.; Dombroskie, J.J.; Jung, S. How a Raindrop Gets Shattered on Biological Surfaces. Proc. Natl. Acad. Sci. USA 2020, 117, 13901–13907. [Google Scholar] [CrossRef]
- Wang, L.; Wang, R.; Wang, J.; Wong, T.S. Compact nanoscale textures reduce contact time of bouncing droplets. Sci. Adv. 2020, 6, eabb2307. Available online: http://www.science.org/doi/10.1126/sciadv.abb2307 (accessed on 1 November 2022). [CrossRef]
- Geyer, F.; D’Acunzi, M.; Sharifi-Aghili, A.; Saal, A.; Gao, N.; Kaltbeitzel, A.; Sloot, T.-F.; Berger, R.; Butt, H.-J.; Vollmer, D. When and How Self-Cleaning of Superhydrophobic Surfaces Works. Sci. Adv. 2020, 6, eaaw9727. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Ji, B.; Feng, L.; Wang, X.; Yang, D.; Rabbi, K.F.; Peng, Q.; Hoque, M.J.; Jin, P.; Bello, E.; et al. Particulate–Droplet Coalescence and Self-Transport on Superhydrophobic Surfaces. ACS Nano 2022, 16, 12910–12921. [Google Scholar] [CrossRef]
- Stork, N.E. How Many Species of Insects and Other Terrestrial Arthropods Are There on Earth? Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Adopted Term | Synonym |
|---|---|
| Hierarchical | Multi-scale, multi-layer, layered |
| Cuticle | Exoskeleton, integument |
| Sculpturing | Textured, pattern, topography |
| Ordered | Homogenous, non-random, organized |
| Disordered | Inhomogeneous, nonhomogeneous, heterogeneous, random, disorganized |
| Pillar | Nipple, nipple-array, nipple-like, tapered rod, wax rod, wax needle, conical protrusion, conical protuberance, protrusion, protuberance, projection, denticle |
| Dome | Bump, hemisphere, protrusion, protuberance, projection, papillae, denticle, ridge |
| Hair | Setae, sensillum, trichia, bristle, spine |
| Wax | Wax crystals, wax rods, wax needles, wax particles, wax powder |
| Structure | Feature, sculpture |
| Groove | Cavity, trough |
| Pattern | Array |
| Scale | Sternite |
| Particle | Granule |
| Brochosome | N/A |
| Micraster | N/A |
| Ground Scale | N/A |
| Cover Scale | N/A |
| Cross-rib | N/A |
| Order | Insect | Structure | Length Scale | References |
|---|---|---|---|---|
| Blattodea | Tree termite (Nasutitermes sp.) | Macrotrichia with troughs Micrasters Hairs | Hierarchical (nano, micro, macro) | [51,52] |
| Termite (Microcerotermes sp.) | ||||
| Termite wings | Microsetae with nanogrooves Micrasters Nanobumps Microbumps Micropillars Denticles | Hierarchical (nano, micro) | [28,48,53] | |
| Tree termite (Nasutitermes walkeri) | ||||
| Coleoptera | Namib desert beetle (unidentified species) | Hydrophilic microbumps with hydrophobic troughs Wax Hexagonal array of flattened micro-hemispheres Setae Alternating hydrophobic and wax-coated, and hydrophilic and non-waxy, regions Macro near-random array of bumps Flattened hemispheres microstructure (regular hexagonal array) | Hierarchical (micro, macro) | [1,21,28,41,48,54,55,56] |
| Desert toktokkie beetle (Physaterna cribripes) | ||||
| Terrestrial leaf beetle (Gastrophysa viridula) | Setae | [57] | ||
| Troutstream beetle (Amphizoa sinica) | [48] | |||
| Darkling beetle (Stenocara sp.) | [54,58] | |||
| Water lily leaf beetle (Galerucela myphaea) | Physical gill setae | Micro | [1,55,59] | |
| Sun beetle (Pachnoda marginata) | Setae | |||
| Darkling beetle (Lagria hirta) | ||||
| Darkling beetle (Zophobas morio) | ||||
| Flower chafer beetle (Mimela testaceipes) | ||||
| Poplar leaf beetle (Chrysomela populi) | ||||
| Aquatic beetle (Stegoelmis sp.) | Plastron scales Plastron sternites Plastron hairs | Hierarchical (nano, micro, macro) | [35] | |
| Tyletelmis mila | ||||
| Elsianus isus | ||||
| Elmis maugei | ||||
| Limnius volckmari | ||||
| Austrolimnus waterhousei | ||||
| Austrolimnus formosus | ||||
| Riffle beetle (Hexacylloepus nunezi) | ||||
| Pilelmis halia | ||||
| Riffle beetle (Cylloepus barberi) | ||||
| Riffle beetle (Cylloepus caicus) | ||||
| Portlemis nevermanni | ||||
| Lixellus haldemani | ||||
| Leaf beetle (Macroplea mutica) | ||||
| Horsetail weevil (Grypus equiseti) | ||||
| Weevil (Bagous americanus) | ||||
| Weevil (Bagous limosus) | ||||
| Golden edge diving beetle (Cybister chinensis) | Ordered submicron-scaled pits along polygonal edges and pores | Micro | [60] | |
| Dung beetle elytra (Geotrupes stercorarius) | Cuticle structure Cement Wax layer | Hierarchical (nano, micro) | ||
| Polyphemus beetle (Mecynorhina polyphemus confluens) | Vertically aligned needles (tomentose section on the elytra) | [61] | ||
| Collembola | Orchesella cincta | Multiscaled rough structure Nanocavities Surface chemistry: lipids with a diverse carbon number triacylglycerols Nanometer-thin triacylglycerol-containing wax layer at the cuticle surface Nanoscopic, comb-like structures | Hierarchical (nano, micro) | [62] [63] |
| Entomobrya intermedia | Setae Hexagonal or rhombic comb-like patterns | [1,21,28] | ||
| Pogonognathellus flavescens | ||||
| Vertagopus arboreus | ||||
| Isotoma viridus | ||||
| Kalaphorura burmeisteri | ||||
| Stenaphoruella quadrispina | ||||
| Dicyrtomina ornata | ||||
| Arrhopalites pygmaeus | ||||
| Tetrodontophora bielanensis | ||||
| Tomocerus fivescens | Hexagonal sculpturing Primary (small) granules with ridges connecting primary structures Secondary (large) granules | [37,64,65] | ||
| Diptera | Tiger crane fly (Nephrotoma australasiae) | Four hair types, some with nanogrooves | Hierarchical (nano, micro) | [66] |
| March flies (Bibionidae) | Microtrichia | Micro | [67] | |
| Thick-headed flies (Conopidae) | ||||
| Soldier flies (Stratiomyidae) | ||||
| Mosquito compound eye | Hexagonally close-packed micro-ommatidia Hexagonally non-close-packed nano-nipples | Hierarchical (nano, micro) | [1,21,68] | |
| Northern house mosquito compound eye (Culex pipiens) | Microhemispheres (ommatidia) Hexagonally non-close-packed nanonipples on ommatidia | [69] | ||
| Green bottle fly eye (Lucilia sericata) | Hexagonally close-packed nanonipples on ommatidia | [70] | ||
| Marsh crane fly (Tipula oleracea) | Nanoscopic and microscopic hairs, some with nanogrooves, on their wings | [54] [1,55] | ||
| Pale giant horse fly (Tabanus bovinus) | ||||
| Marmalade hoverfly (Episyrphus balteatus) | ||||
| Drone fly (Eristalis tenax) | ||||
| Noon fly (Mesembrina meridiana) | ||||
| Horse fly (Tabanus chrysurus | ||||
| Coastal rock pool mosquito larvae (Aedes togoi) | Hydrofuge lobe (snorkel-like breathing apparatus) Oil secretions Lipids | [1,71] | ||
| House fly (Musca domestica) | Microtrichia with nanoscale grooves | Hierarchical (nano, micro, macro) | [36] | |
| Alkali fly (Ephydra hians) | Setae | Micro | [72] | |
| Intertidal midge (Clunio marinus) | Microtrichia | [73] | ||
| Ephemeroptera | Blue-winged olive fly (Ephemerella ignita) | Fractal | Nano | [55] |
| Burrowing mayfly (Ephoron virgo) | ||||
| Mayfly (Ephemera sp.) | [54] | |||
| Ephemera vulgata | [55] | |||
| Hemiptera | Water strider (Aquarius paludum) | Microtrichia Setae | Hierarchical (nano, micro) | [74] |
| Cicada wings (Megapomponia intermedia) | Nanopillars Nanostructures in orderly manner Nipple-like protuberances (cone like base with spherical cap) Denticle Cuticular nanoarrays Nanodomes Disordered inhomogeneous surface Nanostructured conically shaped protrusions | Nano | [21,34,48,49,53,55,68,75,76,77] | |
| Milky cicada (Ayuthia spectabile) | ||||
| Claripennis aguila | ||||
| Pomponia scitula | ||||
| Meimuna conica | ||||
| Meimuna durga | ||||
| Aola bindusara | Nanostructure (protrusion) | [21,34,48,49,53,55,68,75,76,77,78] | ||
| Meimuna mongolica | ||||
| Platylomia radha | ||||
| Dundubia vaginata | ||||
| Dundubia rasingna | ||||
| Meimuna opalifer | [21,34,48,49,53,54,55,68,75,76,77] | |||
| Terpnosia vacua | ||||
| Terpnosia jingpingensis | ||||
| Cryptotympana atrata | [21,34,48,49,53,55,68,75,76,77,78] | |||
| Clear-wing cicada (Psaltoda claripennis) | [21,34,48,49,51,53,55,68,75,76,77,79] | |||
| Chremistica maculata | ||||
| Meimuna microdon | ||||
| Zamara smaragdina | ||||
| Grey cicada (Cicadia orni) | Nipple array Nanocone array | [17,21,34,48,49,53,55,68,75,76,77,80] | ||
| Tettigia orni | ||||
| Leptopsalta bifurcata | Nanostructure (protrusion) | [78] | ||
| Wattle cicada (Cicadetta oldfieldi) | Hexagonally packed spherically capped conical protuberances Clear membrane: similar well-ordered structure size, shape, and periodicity Black membrane: less-ordered surface with individual diamond-shaped structures Relatively large-sized curved projections (bumps), flat (low in height) and spaced many hundreds of nanometers apart | [81] | ||
| Black cicada (Gudanga sp. nr adamsi) | ||||
| Bladder cicada (Cystosoma schemltzi) | ||||
| Scissor grinder cicada (Neotibicen pruinosus) | Nanopillars | [82,83,84,85] | ||
| Dog day cicada (Neotibicen tibicen) | [82] | |||
| Bush cicada (Megatibicen dorsatus) | ||||
| Pharaoh cicada (Magicicada septendecium) | ||||
| Dward periodical cicada (Magicicada cassinii) | [85] | |||
| Backswimmer (Notonecta glauca) | Physical gill setae and microtricia Large sparse setae Small dense microtrichia Sharp-tipped setae Tapered-rod protective wing covers | Hierarchical (nano, micro) | [1,58,86] | |
| Backwimmer (Anisops sp.) | Compressible gas gill | [40,87,88] | ||
| Backswimmer (Notonecta sp.) | Setae (clubs and pins) | Flynn and Bush, 2008 Mail et al., 2018 | ||
| Common water strider (Gerris remigis) | Numerous oriented needle-sharped microsetae with elaborate nanogrooves Papillae Spindly microsetae with nanoscale grooves | Hierarchical (nano, micro, macro) | [39] | |
| Common pond skater (Gerris lacustris) | Physical gill microtrichia Setae | [1,58] | ||
| Water strider leg | Microsetae with nanogrooves Micrasters Nanobumps Microbumps Micropillars Microhairs Microtrichia | Hierarchical (nano, micro) | [21,53,68] | |
| Water-treader (Mesovelia sp.) | Two-tiered hair layer Microtrichia | Micro | [88] | |
| Leafhoppers (Cicadellidae) | Brochosomes with truncated icosahedral geometry Chemistry: protein and lipids Honeycomb-shaped hexagonal and pentagonal structures with re-entrant curvatures | Nano | [1,21,28,89,90] | |
| Planthopper (Desudaba danae) | Microprojections Nano-protuberances Micropillars | Hierarchical (nano, micro) | [1] [14] | |
| River bug (Aphelocheirus aestivalis) | Physical gill cuticular hairs Plastron hairs | [1,35,87] | ||
| Coreidae dock bug (Coreus marginatus) | [55] | |||
| Pentatomidae gorse shield bug (Piezodorus lituratus) | Cuticular hairs | |||
| Pentatomidae stink bug (Carbula putoni) | [54] | |||
| Lantern bug (Limois emelianovi) | ||||
| Naurocoridae saucer bug (Ilyocoris cimicoides) | Setae | Micro | [58] | |
| Naurcoridae (Idiocarus minor) | Plastron hairs | Hierarchical (nano, micro, macro) | [35] | |
| Naurcoridae (Cataractocoris marginiventris) | ||||
| Naurcoridae (Heleocoris mexicanus) | ||||
| Naucoridae water bug (Cryphocricos mexicanus) | Leaf-like setae | |||
| Helotrephidae (Neotrephes usingeri) | Plastron sternites with protuberances | |||
| Lesser water boatman (Corixa punctata) | Setae Microtrichia | Micro | [58] | |
| Agraptocorixa eurynome | Compressible gas gill | [87] | ||
| Poplar spiral gall aphid (Pemphigus spyrothecae) | Powdery wax (long chain esters) Wax needles | [91] | ||
| Hymenoptera | Flower wasp (Scolia soror) | Nanostructures Relatively large-sized curved projections (bumps), flat (low in height), and spaced many hundreds of nanometers apart | Nano | [81] |
| Sawfly larvae (Rhadinoceraea micans) | Complex sculptures Wax crystals Hill-shaped sculptures with radial ridges and crater-like tips | Hierarchical (nano, micro) | [92,93] | |
| Pollinator eyes | Ommatidia cuticular geometry | Nano | [75] | |
| Wasp (Vespa sp.) | Setae | Hierarchical (nano, micro) | [48] | |
| Yellow hornet (Vespa simillima xanthoptera) | ||||
| Black hornet (Vespa dybowskii) | [48] | |||
| European honey bee tongue (Apis Mellifera) | Dense hairs | Micro | [55,94] | |
| German wasp (Paravespula germanica) | [55] | |||
| Red wasp (Vespula rufa schrenckii) | [54] | |||
| Lesser paper wasp (Parapoly varia) | ||||
| Elm sawfly (Arge captiva) | ||||
| Male pollinator fig wasp hind legs (Ceratosolen corneri) | Setae Microtrichia | Hierarchical (nano, micro) | [95] | |
| Red imported fire ant rafts (Solenopsis invicta) | [96] | |||
| Lepidoptera | Giant blue morpho (Morpho didius) | Scales with aligned micro-grooves | Hierarchical (micro, macro) | [34] |
| Moth wing (Prasinocyma albicosta) | Scales with typical overlaying tile type arrangement | Hierarchical (nano, micro) | [81] | |
| Ground Lackey caterpillar (Malacosoma castrensis) | Hair (setae) Microtrichia | [97] | ||
| Malabar tree nymph (Idea malabarica) | Complicated composition of nano- and microstructures Nanostructures of cross-ribs separated by longitudinal ridges | [98] | ||
| Citrus swallowtail (Papilio xuthus) | [51] | |||
| Dark green fritillary (Speyeria aglaja) | Cover scales Nano-/microfeatures Layered cuticle Ommatidia nanonipples | [21,28,48,53,54,55,68,77] | ||
| Menelause blue morpho (Morpho menelaus) | Cover scales Nano-/microfeatures Layered cuticle Ommatidia nanonipples Ordered microstructure Ground scales Wing scales | [21,28,48,53,54,55,68,77,99] | ||
| Indian cabbage white (Artogeia canidia) | Cover scales Nano-/microfeatures Layered cuticle Ommatidia nanonipples | [21,28,48,53,54,55,68,77] | ||
| Aega morpho (Morpho aega) | Cover scales Nano-/microfeatures Layered cuticle | |||
| Horse-chestnut leaf miner (Cameraria ohridella) | ||||
| Boarmia ribeata | ||||
| Wood carpet moth (Cidaria rivata) | ||||
| Chinese tussar moth (Autographa pernyi) | ||||
| Small skipper (Thymelicus sylvestris) | ||||
| Cabbage white (Pieris brassicae) | ||||
| Small tortoiseshell butterfly (Aglais urticae) | ||||
| Marbled white (Melanargia galathea) | ||||
| Mulberry tiger moth (Lemyra imparilis) | Setae with nanogrooves Macrotrichia with longitudinal ridges and troughs | Hierarchical (nano, micro, macro) | [100] | |
| Mecoptera | Meadow scorpionfly (Panorpa vulgaris) | Setae with nanogrooves Macrotrichia with longitudinal ridges and troughs | Hierarchical (nano, micro, macro) | [55] |
| Megaloptera | Alderfly (Sialis lutaria) | Setae with nanogrooves Macrotrichia with longitudinal ridges and troughs | Hierarchical (nano, micro, macro) | [55] |
| Odonata | Vagrant darter dragonfly (Sympetrum vulgatum) | Randomly oriented nanopillars at various scales | Nano | [34,48,53,54,55,77] |
| Red-veined darter dragonfly (Sympetrum fonscolombii) | Waxy epicuticular layer [with] unique surface nanoarchitecture that consists of irregular arrays of nanoscale pillars | [101] | ||
| Dragonfly wings | Clear C-H stretching bands, prevalence of methylene bands which indicate long-chain aliphatic hydrocarbons | [76] | ||
| Yellow-striped flutterer dragonfly (Rhyothemis phyllis chloe) | Rod-like structures | [81] | ||
| Spread-winged damselfly (Lestes sponsa) | Wax rods Nanostructures Nanopillar array | [17,102] | ||
| Australian emperor dragonfly (Hemianex papuensis) | Randomly oriented nanopillars Fractal Nanostructures Nanospikes Nanomembrane surface appeared as a nanoscale mesh with rough spikes | [51] | ||
| Blue-tailed damselfly (Ischnura elegans) | [55] | |||
| Common skimmer dragonfly (Orthetrum albistylum speciosum) | [103,104] | |||
| Wandering glider dragonfly (Pantala flavescens) | Waxy covering and geometric non-smooth structure-column papillae Long-chain hydrocarbons, fatty acids, alcohols, and esters | |||
| Tau emerald dragonfly (Hemicordulia tau) | Nanostructures Nanoscale pillars Epicuticular lipids (primarily aliphatic hydrocarbons, especially n-alkanes with even-number chain lengths between C19 and C26, and relatively small proportion of palmitic acid) | [101] | ||
| Brown darner dragonfly (Gynacantha dravida) | Oblate-shaped (chitin) nano-fibrils | [105] | ||
| Blue riverdamsel (Pseudagrion microcephalum) | ||||
| Swamp flat-tail dragonfly (Austrothemis nigrescens) | Waxy epicuticular layer with unique surface nanoarchitecture that consists of irregular arrays of nanoscale pillars Clear C-H stretching bands, prevalence of methylene bands which indicate long-chain aliphatic hydrocarbons | [101] | ||
| Epaulet skimmer dragonfly (Orthetrum chrysostigma) | ||||
| Violet dropwing dragonfly (Trithemis annulata) | ||||
| Lesser emperor dragonfly (Anax parthenope) | ||||
| Emperor dragonfly (Anax imperator) | ||||
| Green-eyed hook-tail dragonfly (Onychogomphus forcipatus) | ||||
| Wandering percher dragonfly (Diplacodes bipunctata) | Nanopillars Cuticular waxes: hydroxyl, alkyl hydrocarbons, ester carbonyl, amide groups, long-chain aliphatic hydrocarbons | [48] | ||
| Black-faced percher (Diplacodes melanopsis) | ||||
| Common bluetail damselfly (Ischnura heterosticta) | ||||
| Red and blue damselfly (Xanthagrion erythroneurum) | ||||
| Banded demoiselle damselfly (Calopteryx splendens) | Long wax rods on wing veins Wax crystals of various shapes | Hierarchical (nano, micro) | [106,27] | |
| Orthoptera | Chinese grasshopper (Acrida cinerea cinerea) | Denticle | Hierarchical (nano, micro) | [48,54] |
| Long-headed grasshopper (Atractomorpha latta) | ||||
| Sickle-bearing bush-cricket (Phaneroptera falcata) | [55] | |||
| Common field grasshopper (Chorthippus brunneus) | ||||
| Oriental migratory locust (Locusta migratoria) | ||||
| Mottled grasshopper (Myrmeleotettix maculatus) | ||||
| Neuroptera | Banded lacewing (Glenoleon pulchellus) | Nanostructures Wing membrane: interconnected ridges forming a dense netting on the cuticle surface Vein regions: array of macrotrichia | Hierarchical (nano, micro, macro) | [48] |
| Common green lacewing (Chrysoperla carnea) | [55] | |||
| Ant lion (Grocus bore) | [54] | |||
| Mantid fly (Mantispa sp.) | ||||
| Glenuroides japonicus | ||||
| Green lacewing (Chrysopa oculata) | Mactrotrichia on wing veins (angled toward cells) with longitudinal ridges and troughs along hair shaft Dense netting of nanopillars on wing membrane | [107] | ||
| Mantid lacewing (Ditaxis biseriata) | ||||
| Australian blue eyes lacewing (Nymphes myrmeleonides) | ||||
| Trichoptera | Net-spinning caddisfly (Hydropsyche pellucidula) | Mactrotrichia on wing veins (angled toward cells) with longitudinal ridges and troughs along hair shaft Dense netting of nanopillars on wing membrane | Hierarchical (nano, micro, macro) | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bello, E.; Chen, Y.; Alleyne, M. Staying Dry and Clean: An Insect’s Guide to Hydrophobicity. Insects 2023, 14, 42. https://doi.org/10.3390/insects14010042
Bello E, Chen Y, Alleyne M. Staying Dry and Clean: An Insect’s Guide to Hydrophobicity. Insects. 2023; 14(1):42. https://doi.org/10.3390/insects14010042
Chicago/Turabian StyleBello, Elizabeth, Yutao Chen, and Marianne Alleyne. 2023. "Staying Dry and Clean: An Insect’s Guide to Hydrophobicity" Insects 14, no. 1: 42. https://doi.org/10.3390/insects14010042