

Direct and Indirect Competitive Interactions between Ooencyrtus nezarae and Paratelenomus saccharalis Parasitizing Megacopta cribraria Egg Patches
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants
2.2. Insect Rearing
2.3. Physiological Condition of Insects
2.4. Experiment I: Timing of Adult Arrival at the Competition
2.5. Experiment II: Characterizing Aggressive Behavior of Parasitoids
2.6. Experiment III: Intraguild Predation of Parasitoid Larvae
3. Data and Statistical Analysis
4. Results
4.1. Experiment I: The Timing of Adult Arrival at the Competition
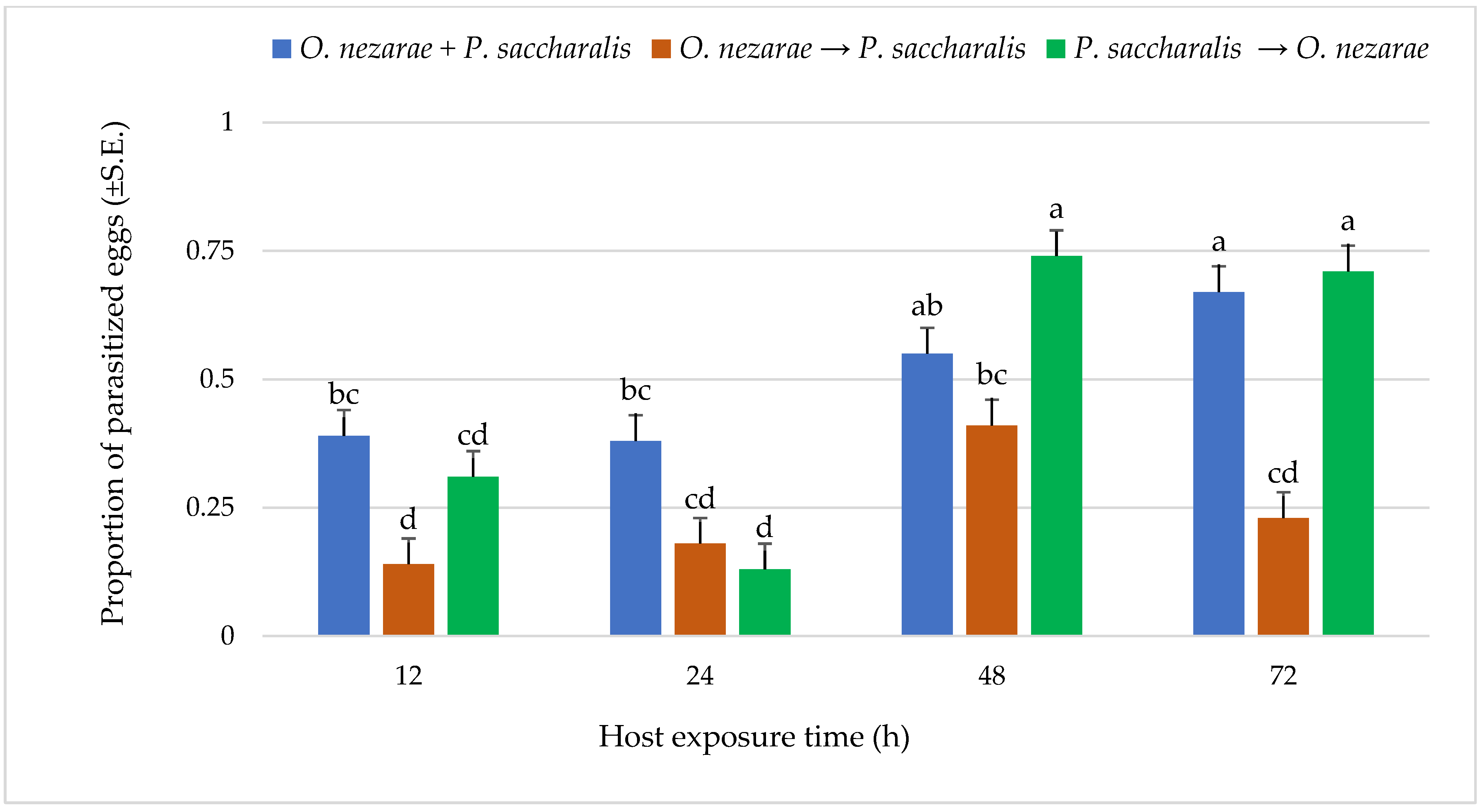
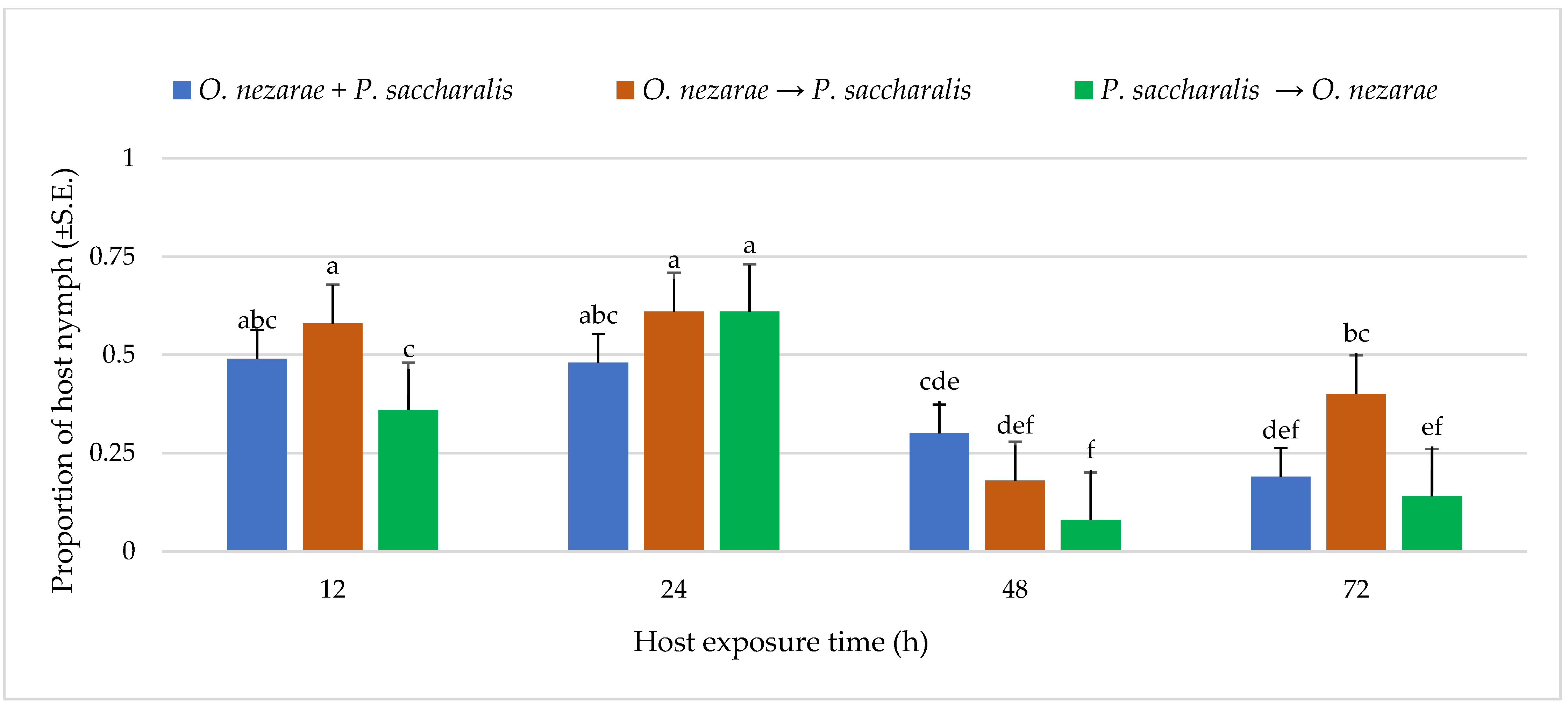
4.2. Experiment II: Characterizing Aggressive Behavior
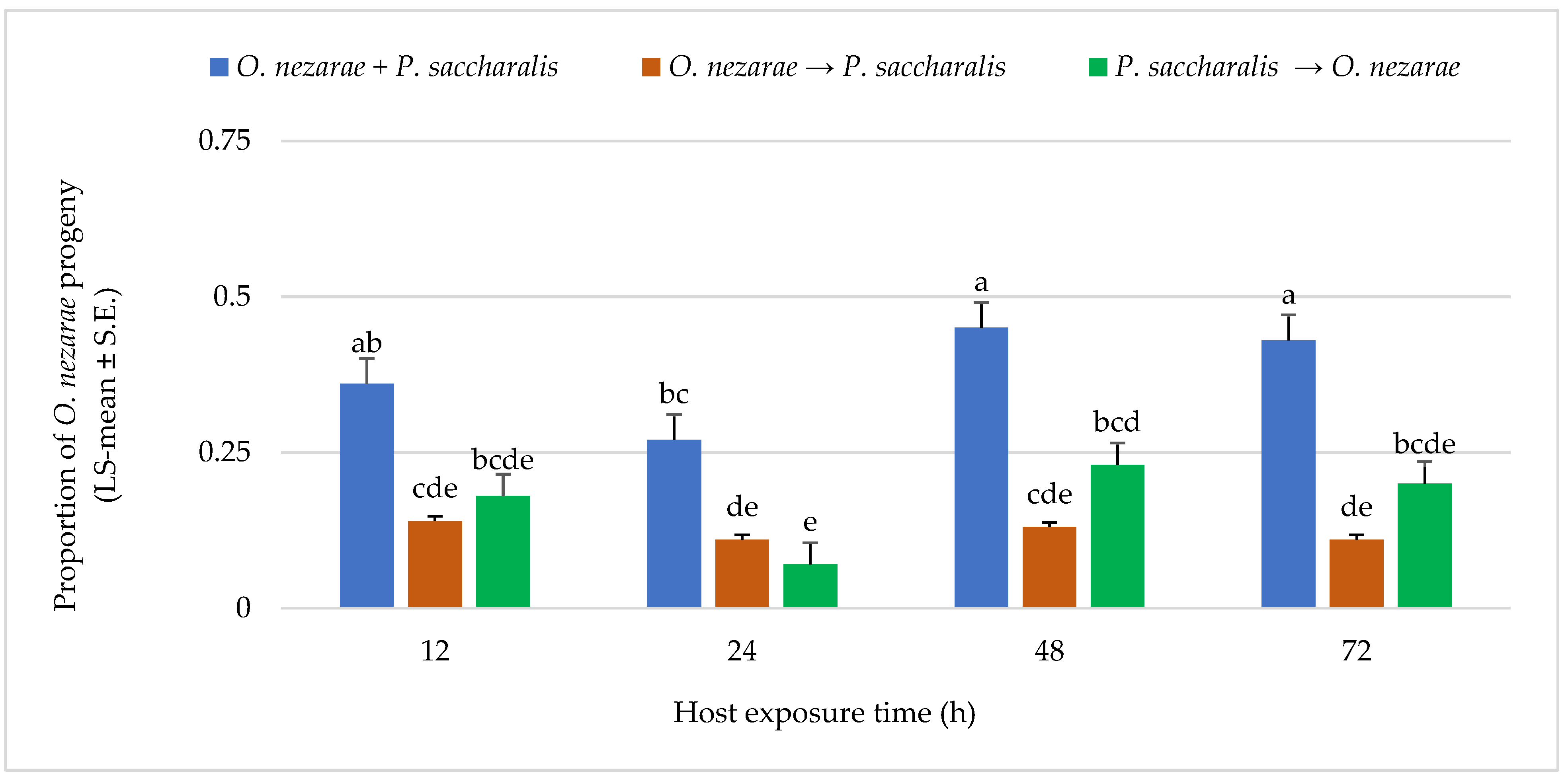
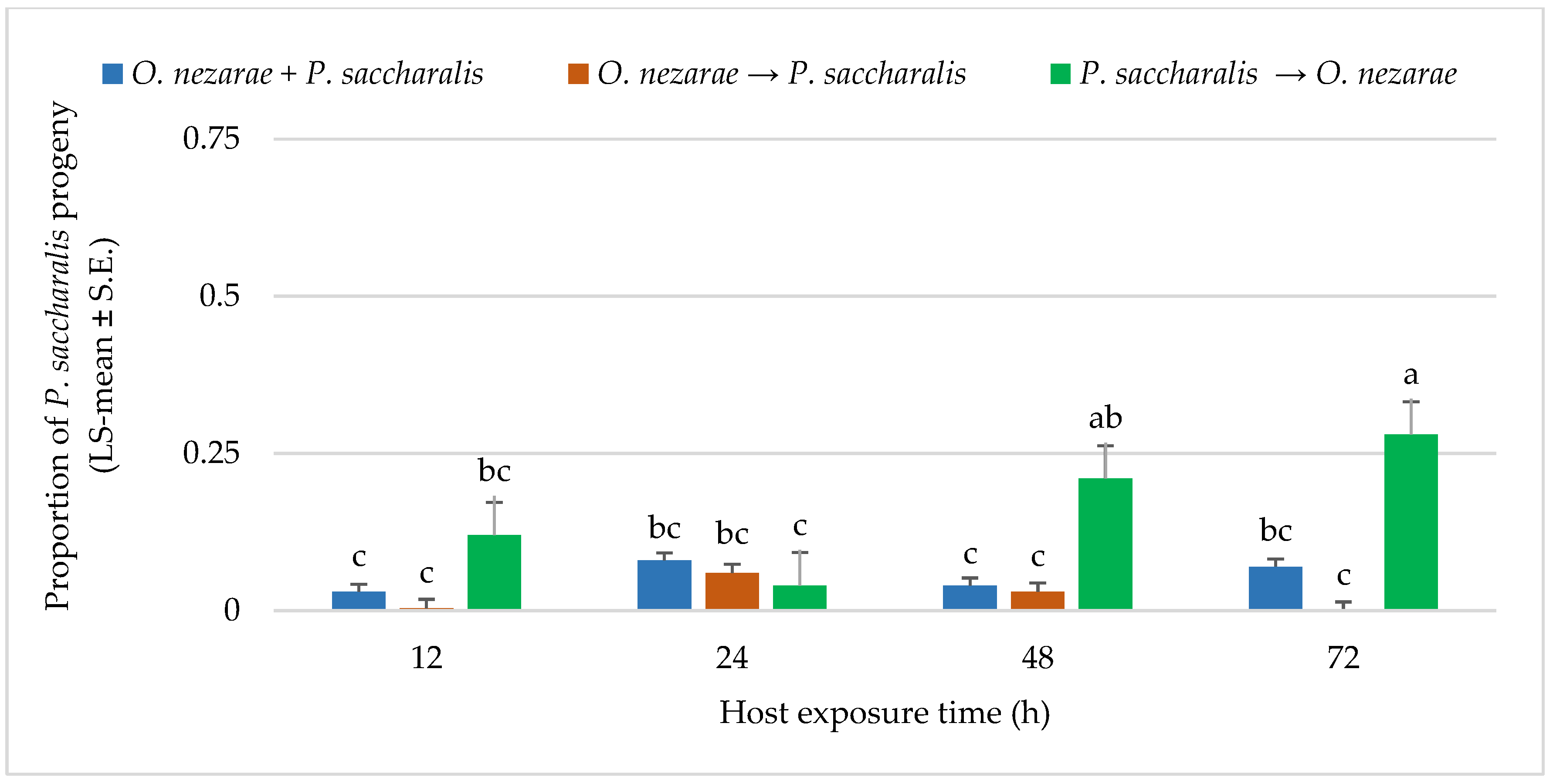
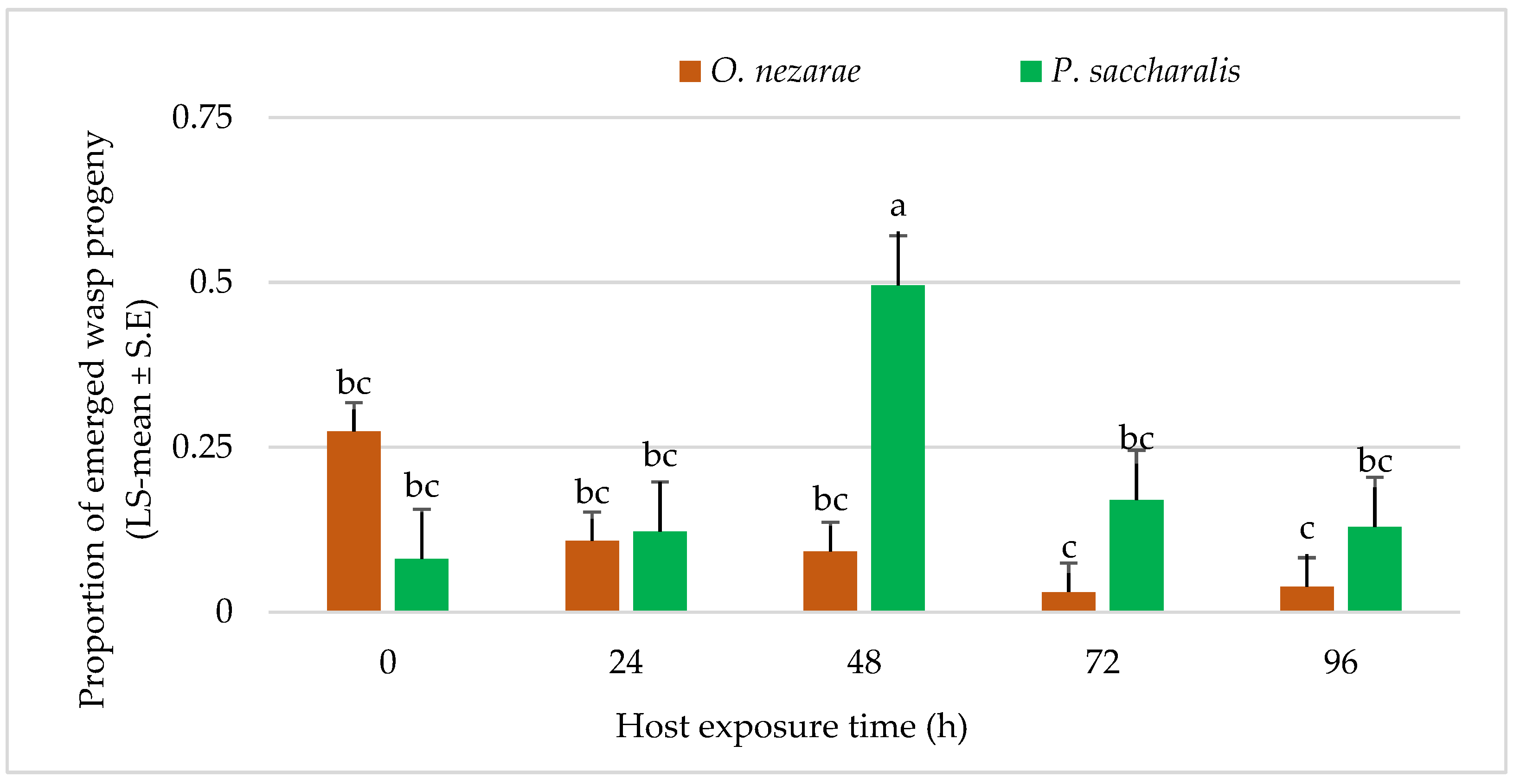
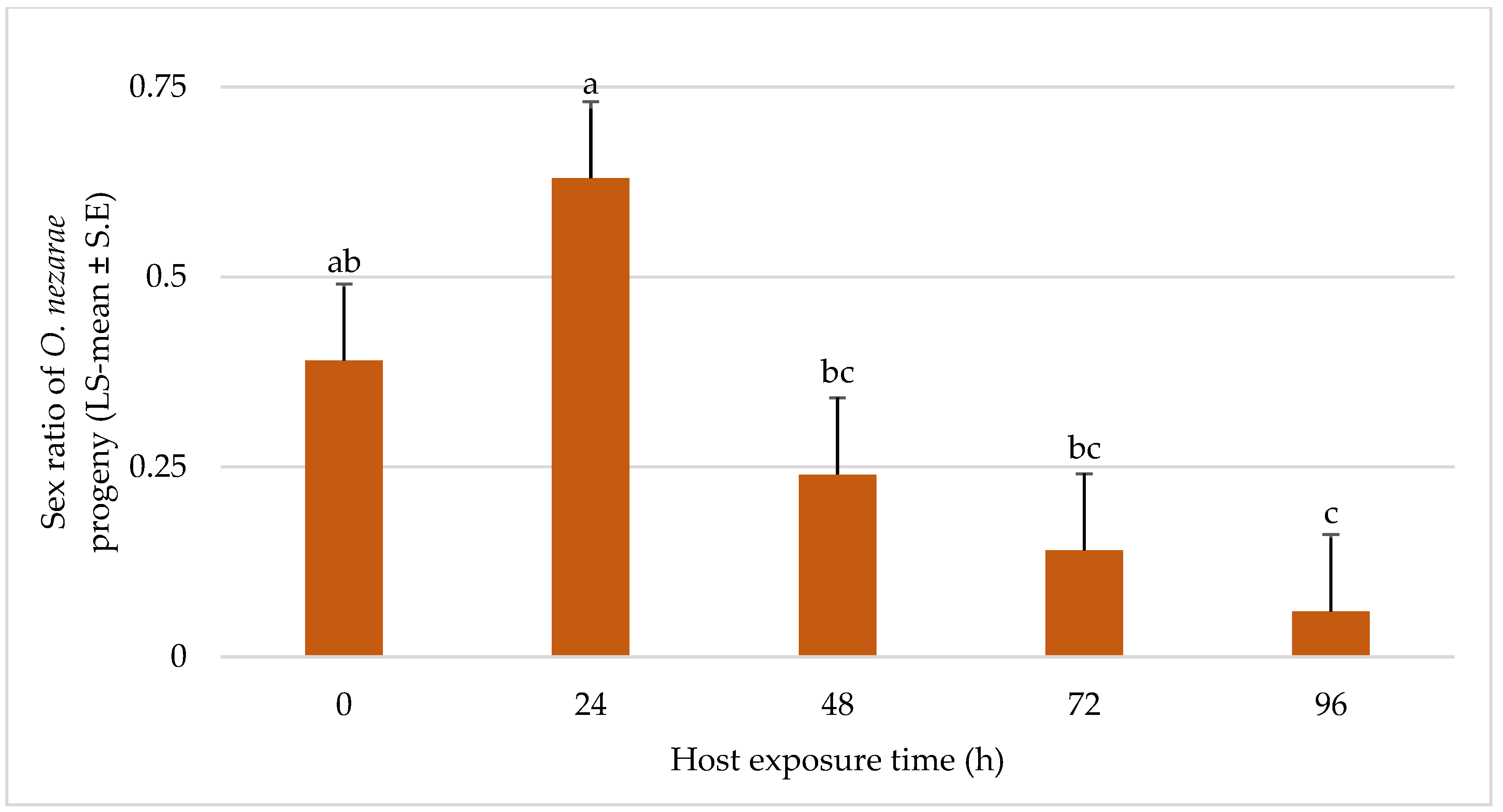
4.3. Experiment III: Intraguild Predation of Parasitoid Larvae
5. Discussion
5.1. Experiment I: The Timing of Adult Arrival at the Competition
5.2. Experiment II: Characterizing Aggressive Behavior
5.3. Experiment III: Intraguild Predation of Larvae
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harvey, J.A.; Poelman, E.H.; Tanaka, T. Intrinsic inter-and intraspecific competition in parasitoid wasps. Annu. Rev. Entomol. 2013, 58, 333–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, P.W. Parasitiods utilizing the same host: Adaptive nature of differences in size and form. Ecology 1972, 53, 190–195. [Google Scholar] [CrossRef]
- Bográn, C.E.; Heinz, K.M.; Ciomperlik, M.A. Interspecific competition among insect parasitoids: Field experiments with whiteflies as hosts in cotton. Ecology 2002, 83, 653–668. [Google Scholar] [CrossRef]
- Polis, G.A.; Strong, D.R. Food web complexity and community dynamics. Am. Nat. 1996, 147, 813–846. [Google Scholar] [CrossRef]
- Harvey, J.A.; Wagenaar, R.; Martijn Bezemer, T. Life-history traits in closely related secondary parasitoids sharing the same primary parasitoid host: Evolutionary opportunities and constraints. Entomol. Exp. Appl. 2009, 132, 155–164. [Google Scholar] [CrossRef]
- Brodeur, J.; Rosenheim, J.A. Intraguild interactions in aphid parasitoids. Entomol. Exp. Appl. 2000, 97, 93–108. [Google Scholar] [CrossRef] [Green Version]
- Cusumano, A.; Peri, E.; Bradleigh, V.S.; Colazza, S. Interspecific extrinsic and intrinsic competitive interactions in egg parasitoids. BioControl 2012, 57, 719–734. [Google Scholar] [CrossRef]
- Mohammadpour, M.; Jalali, M.A.; Michaud, J.P.; Ziaaddini, M.; Hashemirad, H. Multiparasitism of stink bug eggs: Competitive interactions between Ooencyrtus pityocampae and Trissolcus agriope. BioControl 2014, 59, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Frago, E. Interactions between parasitoids and higher order natural enemies: Intraguild predation and hyperparasitoids. Curr. Opin. Insect. Sci. 2016, 14, 81–86. [Google Scholar] [CrossRef]
- Van Veen, F.J.; Rajkumar, A.; Muller, C.B.; Godfray, H.C. Increased reproduction by pea aphids in the presence of secondary parasitoids. Ecol. Entomol. 2001, 26, 425–429. [Google Scholar] [CrossRef]
- Hassell, M.P.; Varley, G.C. New inductive population model for insect parasites and its bearing on biological control. Nature 1969, 223, 1133–1137. [Google Scholar] [CrossRef]
- Hassell, M.P. Mutual interference between searching insect parasites. J. Anim. Ecol. 1971, 40, 473–486. [Google Scholar] [CrossRef]
- Boivin, G.; Brodeur, J. Intra-and interspecific interactions among parasitoids: Mechanisms, outcomes and biological control. In Trophic and Guild in Biological Interactions Control; Springer: Dordrecht, The Netherlands, 2006; pp. 123–144. [Google Scholar] [CrossRef]
- Zwolfer, H. The structure and effect of parasite complexes attacking phytophagous host insects. In Dynamics of Populations, Proceedings of the Advanced Study Institute on ‘Dynamics and Numbers in Populations’, Oosterbeck, The Netherlands, 7–18 September 1970; den Boer, P.J., Gradwell, G.R., Eds.; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1971; pp. 405–418. [Google Scholar]
- Rosenheim, J.A.; Kaya, H.K.; Ehler, L.E.; Marois, J.J.; Jaffee, B.A. Intraguild predation among biological-control agents: Theory and evidence. Biol. Control 1985, 5, 303–335. [Google Scholar] [CrossRef]
- De Moraes, C.M.; Cortesero, A.M.; Stapel, J.O.; Lewis., W.J. Intrinsic and extrinsic competitive interactions between two larval parasitoids of Heliothis virescens. Ecol. Entomol. 1999, 24, 402–410. [Google Scholar] [CrossRef]
- Cusumano, A.; Peri, E.; Colazza, S. Interspecific competition/facilitation among insect parasitoids. Curr. Opin. Insect Sci. 2016, 14, 12–16. [Google Scholar] [CrossRef]
- Overington, S.E.; Dubois, F.; Lefebvre, L. Food unpredictability drives both generalism and social foraging: A game theoretical model. Behav. Ecol. 2008, 19, 836–841. [Google Scholar] [CrossRef] [Green Version]
- Dorn, S.; Beckage, N.E. Superparasitism in gregarious hymenopteran parasitoids: Ecological, behavioural and physiological perspectives. Physiol. Entomol. 2007, 32, 199–211. [Google Scholar] [CrossRef]
- Gandon, S.; Rivero, A.; Varaldi, J. Superparasitism evolution: Adaptation or manipulation? Am. Nat. 2006, 167, E1–E22. [Google Scholar] [CrossRef] [Green Version]
- Brodeur, J. Host specificity and trophic relationships of hyperparasitoids. In Parasitoid Population Biology; Hochberg, M.E., Ives, A.R., Eds.; Princeton University Press: Princeton, NJ, USA, 2000; pp. 163–183. [Google Scholar]
- Hunter, M.S.; Woolley, J.B. Evolution and behavioral ecology of heteronomous aphelinid parasitoids. Annu. Rev. Entomol. 2001, 46, 251–290. [Google Scholar] [CrossRef]
- Tillman, P.G.; Powell, J.E. Interspecific discrimination and larval competition among Microplitis croceipes, Microplitis demolitor, Cotesia kazak (Hym, Braconidae), and Hyposoter didymator (Hym, Ichneumonidae), parasitoids of Heliothis virescens (Lep, Noctuidae). Entomophaga 1992, 37, 439–451. [Google Scholar] [CrossRef]
- Uka, D.; Hiraoka, T.; Iwabuchi, K. Physiological suppression of the larval parasitoid Glyptapanteles pallipes by the polyembryonic parasitoid Copidosoma floridanum. J. Insect Physiol. 2006, 52, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Laing, J.E.; Corrigan, J.E. Intrinsic competition between the gregarious parasite, Cotesia glomeratus and the solitary parasite Cotesia rubecula [Hymenoptera: Braconidae] for their host Artogeia rapae [Lepidoptera: Pieridae]. Entomophaga 1987, 32, 493–501. [Google Scholar] [CrossRef]
- Kfir, R.; Van Hamburg, H. Interspecific competition between Telenomus ullyetti (Hymenoptera:Scelionidae) and Trichogrammatoidea lutea (Hymenoptera: Trichogrammatidae) parasitizing eggs of the cotton bollworm Heliothis armiger in the laboratory. Environ. Entomol. 1988, 17, 664–670. [Google Scholar] [CrossRef]
- Wang, X.G.; Messing, R.H. Intra-and interspecific competition by Fopius arisanus and Diachasmimorpha tryoni (Hymenoptera: Braconidae), parasitoids of tephritid fruit flies. Biol. Control 2003, 27, 251–259. [Google Scholar] [CrossRef]
- Takasu, K.; Hirose, Y. Seasonal egg parasitism of phytophagous stink bugs in a soybean field in Fukuoka. In Proc. Assoc. Pl. Prot. Kyushu 1985, 31, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Takasu, K.; Hirose, Y. Kudzu-vine community as a breeding site of Ooencyrtus nezarae Ishi (Hymenoptera: Encyrtidae), an egg parasitoid of bugs attacking soybean. Jpn. J. Appl. Entomol. Zool. 1986, 30, 302–304. [Google Scholar] [CrossRef]
- Ademokoya, B.; Balusu, R.; Morawo, T.; Fadamiro, H. Attraction of Paratelenomus saccharalis (Hymenoptera: Platygastridae), an egg parasitoid of Megacopta cribraria (Hemiptera: Plataspidae), to host associated olfactory cues. J. Entomol. Sci. 2017, 52, 323–331. [Google Scholar] [CrossRef]
- Hoshino, K.; Adati, T.; Olson, D.M.; Takasu, K. Seasonal occurrence and interspecific interactions of egg parasitoids of Megacopta cribraria (heteroptera: Plataspidae) in Japan. Environ. Entomol. 2017, 46, 487–493. [Google Scholar] [CrossRef]
- Alim, M.A.; Lim, U. Ecotoxicological effect of insecticides on Ooencyrtus nezarae (Hymenoptera: Encyrtidae) reared from refrigerated and unrefrigerated Riptortus pedestris (Hemiptera: Alydidae) host. Biocontrol Sci. Technol. 2014, 24, 133–144. [Google Scholar] [CrossRef]
- Knight, I.A.; Roberts, P.M.; Gardner, W.A.; Oliver, K.M.; Reay-Jones, F.P.; Reisig, D.D.; Toews, M.D. Spatial distribution of Megacopta cribraria (Hemiptera: Plataspidae) adults, eggs and parasitism by Paratelenomus saccharalis (Hymenoptera: Platygastridae) in soybean. Environ. Entomol. 2017, 46, 1292–1298. [Google Scholar] [CrossRef]
- Johnson, N.F. Revision of world species of Paratelenomus Dodd (Hymenoptera: Scelionidae). Can. Entomol. 1996, 128, 273–291. [Google Scholar] [CrossRef] [Green Version]
- Eger, J.E., Jr.; Pitcher, A.J.; Halbert, S.E.; Penca, C.; Hodges, A.C. First report of Brachyplatys subaeneus (Westwood) (Hemiptera: Heteroptera: Plataspidae) in the United States. Insecta Mundi 2020, 0814, 1–6. Available online: https://journals.flvc.org/mundi/article/download/126889/126532 (accessed on 26 June 2022).
- Hirose, Y.; Takasu, K.; Takagi, M. Egg parasitoids of phytophagous bugs in soybean: Mobile natural enemies as naturally occurring biological control agents of mobile pests. Biol. Control 1996, 7, 84–94. [Google Scholar] [CrossRef]
- Mizutani, N.; Wada, T.; Higuchi, H.; Ono, M.; Leal, S.W. A component of a synthetic aggregation pheromone of Riptortus clavatus (Thunberg) (Heteroptera: Alydidae), that attracts an egg parasitoid, Ooencyrtus nezarae Ishii (Hymenoptera: Encyrtidae). Appl. Entomol. Zool. 1997, 32, 504–507. [Google Scholar] [CrossRef] [Green Version]
- Ademokoya, B.; Balusu, R.; Ray, C.; Mottern, J.; Fadamiro, H. The first record of Ooencyrtus nezarae (Hymenoptera: Encyrtidae) on kudzu bug (Hemiptera: Plataspidae) in North America. J. Insect Sci. 2018, 18, 8. [Google Scholar] [CrossRef] [Green Version]
- Takasu, K.; Hirose, Y. The parasitoid Ooencyrtus nezarae (Hymenoptera: Encyrtidae) prefers hosts parasitized by conspecifics over unparasitized hosts. Oecologia 1991, 87, 319–323. [Google Scholar] [CrossRef]
- Chicas-Mosier, A.M.; Balusu, R.R.; Ajayi, O.S.; Kafle, B.D.; Morawo, T.; Mertoglu, G.; Smith, C.M.; Warsi, S.; Fadamiro, H.Y. Kudzu Bug (Megacopta cribraria) and associated egg parasitoids emergence rates in Alabama are predicted by weather indices. Environ. Entomol. 2022, 51, 1113–1119. [Google Scholar] [CrossRef]
- Potting, R.P.J.; Snellen, H.M.; Vet, L.E.M. Fitness consequences of superparasitism and mechanism of host discrimination in the stemborer parasitoid Cotesia flavipes. Entomol. Exp. Appl. 1997, 82, 341–348. [Google Scholar] [CrossRef]
- Singer, M.S.; Stireman, J., III. O. Does anti-parasitoid defense explain host-plant selection by a polyphagous caterpillar? Oikos 2003, 100, 554–562. [Google Scholar] [CrossRef]
- Dubois, F.; Giraldeau, L.A.; Grant, J.W. Resource defense in a group-foraging context. Behav. Ecol. 2003, 14, 2–9. [Google Scholar] [CrossRef]
- Benelli, G. Should I fight or should I flight? How studying insect aggression can help integrated pest management. Pest Manag. Sci. 2015, 71, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Christopher, Y. Agonistic behavior. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T., Eds.; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Takasu, K.; Hirose, Y. The number of larval instars in Ooencyrtus species (Hymenoptera, Encyrtidae). Jpn. J. Entomol. 1989, 57, 398–401. [Google Scholar]
- Cusumano, A.; Peri, E.; Vinson, S.B.; Colazza, S. Intraguild interactions between two egg parasitoids exploring host patches. BioControl 2011, 56, 173–184. [Google Scholar] [CrossRef]
- Britt, K.E. An Ecological Study of the Kudzu Bug in East Tennessee: Life History, Seasonality, and Phenology. Master’s Thesis, University of Tennessee, Knoxville, TN, USA, 2016. [Google Scholar]
- Gardner, W.A.; Blount, J.L.; Golec, J.R.; Jones, W.A.; Hu, X.P.; Talamas, E.J.; Evans, R.M.; Dong, X.; Ray, C.H., Jr.; Buntin, G.D.; et al. Discovery of Paratelenomus saccharalis (Dodd) (Hymenoptera: Platygastridae), an egg parasitoid of Megacopta cribraria F. (Hemiptera: Plataspidae) in its expanded North American range. J. Entomol. Sci. 2013, 48, 355–359. [Google Scholar] [CrossRef] [Green Version]
- Aung, K.S.D.; Takagi, M.; Ueno, T. Effect of female’s age on the progeny production and sex ratio of Ooencyrtus nezarae, an egg parasitoid of the bean bug Riptortus clavatus. J. Fac. Agr. Kyushu Univ. 2010, 55, 83–85. [Google Scholar] [CrossRef]
- Takano, S.I.; Takasu, K. Food deprivation increases reproductive effort in a parasitoid wasp. Biol. Control 2019, 133, 75–80. [Google Scholar] [CrossRef]
- Ganjisaffar, F.; Perring, T.M. Life history evaluation of Ooencyrtus lucidus, a newly described egg parasitoid of Bagrada hilaris. Insects 2020, 11, 292. [Google Scholar] [CrossRef]
- Benelli, G.; Canale, A. Aggressive behavior in olive fruit fly females: Oviposition site guarding against parasitic wasps. J. Insect Behav. 2016, 29, 680–688. [Google Scholar] [CrossRef]
- Krasylenko, Y.; Těšitel, J.; Ceccantini, G.; Oliveira-da-Silva, M.; Dvořák, V.; Steele, D.; Sosnovsky, Y.; Piwowarczyk, R.; Watson, D.M.; Teixeira-Costa, L. Parasites on parasites: Hyper-, epi-, and autoparasitism among flowering plants. Am. J. Bot. 2021, 108, 8–21. [Google Scholar] [CrossRef]
- Tian, S.P.; Zhang, J.H.; Yan, Y.H.; Wang, C.Z. Interspecific competition between the ichneumonid Campoletis chlorideae and the braconid Microplitis mediator in their host Helicoverpa armigera. Entomol. Exp. Appl. 2008, 127, 10–19. [Google Scholar] [CrossRef]
- Bernays, E.A. Neural limitations in phytophagous insects: Implications for diet breadth and evolution of host affiliation. Annu. Rev. Entomol. 2001, 46, 703–727. [Google Scholar] [CrossRef] [Green Version]
- Kidd, D.; Amarasekare, P. The role of transient dynamics in biological pest control: Insights from a host parasitoid community. J. Anim. Ecol. 2012, 81, 47–57. [Google Scholar] [CrossRef]
- Vinson, S.B. Competition and host discrimination between two species of tobacco budworm parasitoids. Ann. Entomol. Soc. Am. 1972, 65, 229–236. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
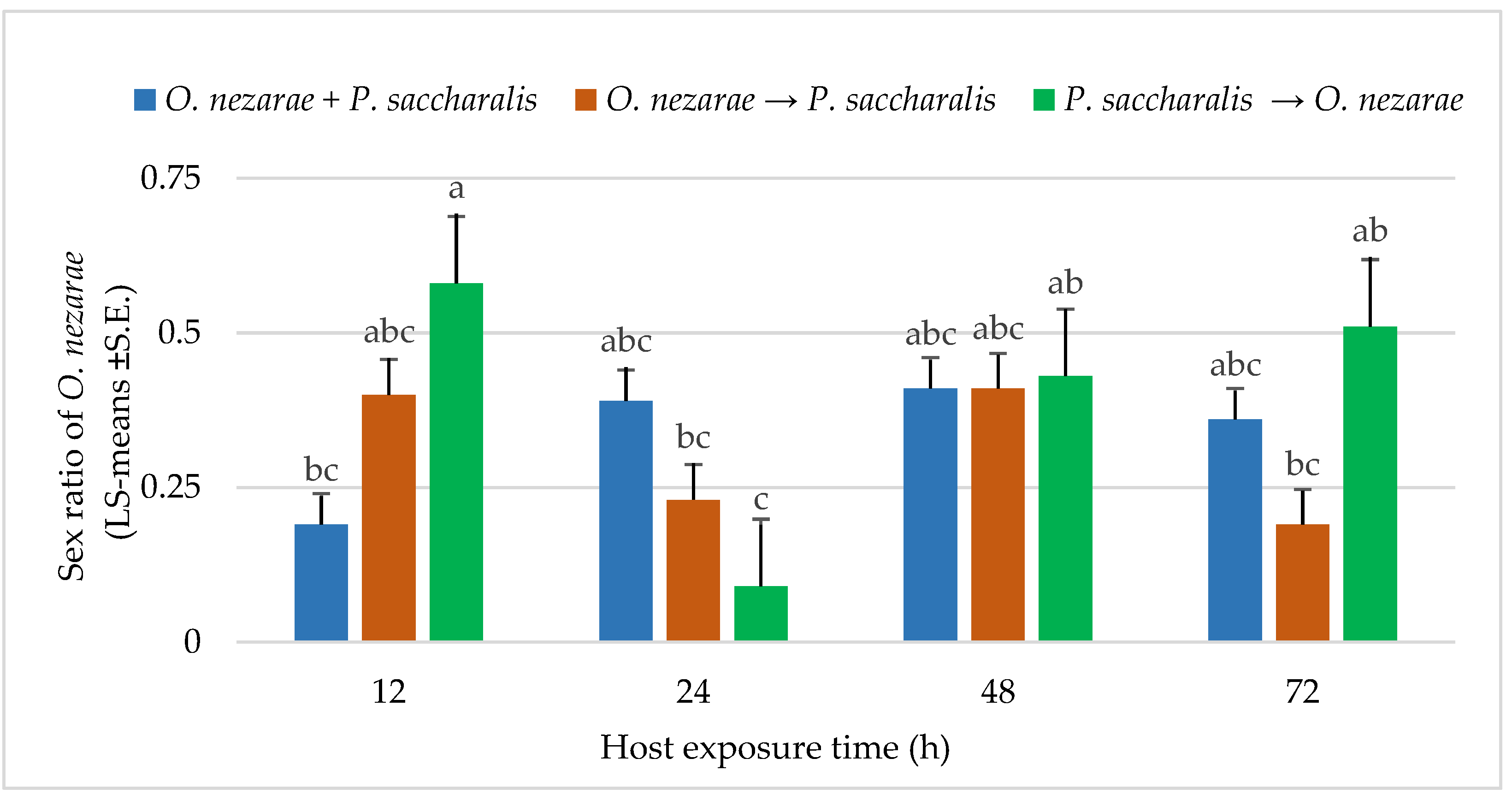{kind=link}
{kind=link}
{kind=link}
| (Proportion ± SE) | ||
|---|---|---|
| Treatment | M. cribraria Parasitized Eggs | Emerged Wasp |
| Simultaneous release | 0.50 ± 0.025 a | 0.43 ± 0.02 a |
| Sequential release with O. nezarae first | 0.24 ± 0.025 b | 0.15 ± 0.02 c |
| Sequential release with P. saccharalis first | 0.47 ± 0.025 a | 0.34 ± 0.02 b |
| ANOVA for the main effect | F = 19.91; df = 2, 237; p < 0.0001 | F = 32.32; df = 2, 237; p < 0.0001 |
| (Proportion ± S.E) | ||
|---|---|---|
| Host Exposure Time (h) | M. cribraria Parasitized Eggs | Emerged Wasp |
| 0 | 0.38 ± 0.07 ab | 0.35 ± 0.06 ab |
| 24 | 0.35 ± 0.07 ab | 0.23 ± 0.06 b |
| 48 | 0.59 ± 0.07 a | 0.58 ± 0.06 a |
| 72 | 0.23 ± 0.07 b | 0.20 ± 0.06 b |
| 96 | 0.19 ± 0.07 b | 0.16 ± 0.06 b |
| ANOVA for main effect | F = 5.21; df = 4, 95; p = 0.0008 | F = 7.90; df = 4, 95; p < 0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Warsi, S.; Chicas-Mosier, A.M.; Balusu, R.R.; Jacobson, A.L.; Fadamiro, H.Y. Direct and Indirect Competitive Interactions between Ooencyrtus nezarae and Paratelenomus saccharalis Parasitizing Megacopta cribraria Egg Patches. Insects 2023, 14, 35. https://doi.org/10.3390/insects14010035
Warsi S, Chicas-Mosier AM, Balusu RR, Jacobson AL, Fadamiro HY. Direct and Indirect Competitive Interactions between Ooencyrtus nezarae and Paratelenomus saccharalis Parasitizing Megacopta cribraria Egg Patches. Insects. 2023; 14(1):35. https://doi.org/10.3390/insects14010035
Chicago/Turabian StyleWarsi, Sanower, Ana M. Chicas-Mosier, Rammohan R. Balusu, Alana L. Jacobson, and Henry Y. Fadamiro. 2023. "Direct and Indirect Competitive Interactions between Ooencyrtus nezarae and Paratelenomus saccharalis Parasitizing Megacopta cribraria Egg Patches" Insects 14, no. 1: 35. https://doi.org/10.3390/insects14010035