

Development of a Kairomone-Based Attractant as a Monitoring Tool for the Cocoa Pod Borer, Conopomorpha cramerella (Snellen) (Lepidoptera: Gracillariidae)
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials Tested
2.2. Experimental Sites
2.3. Experimental Design
2.4. Field Experiments
2.5. Statistics
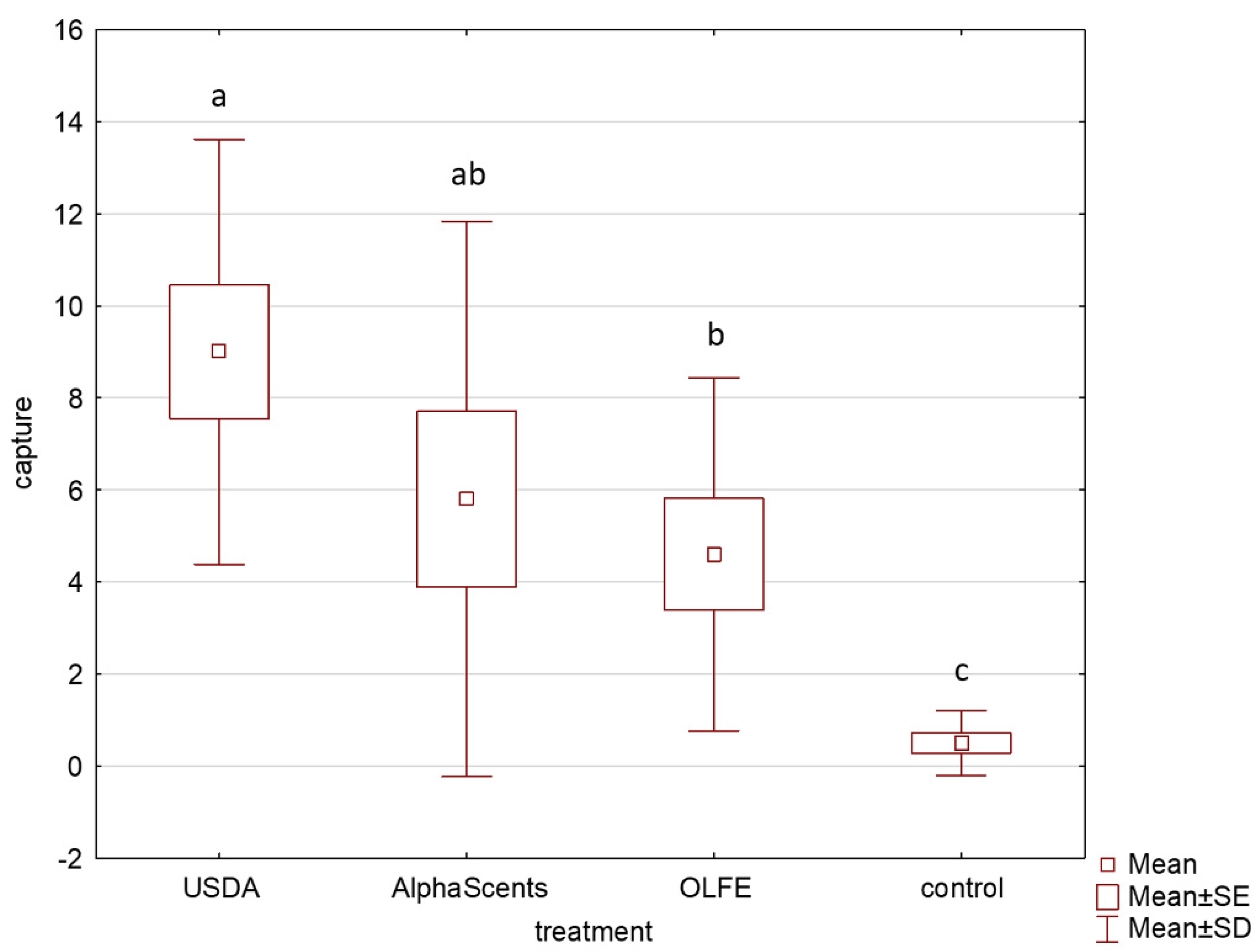
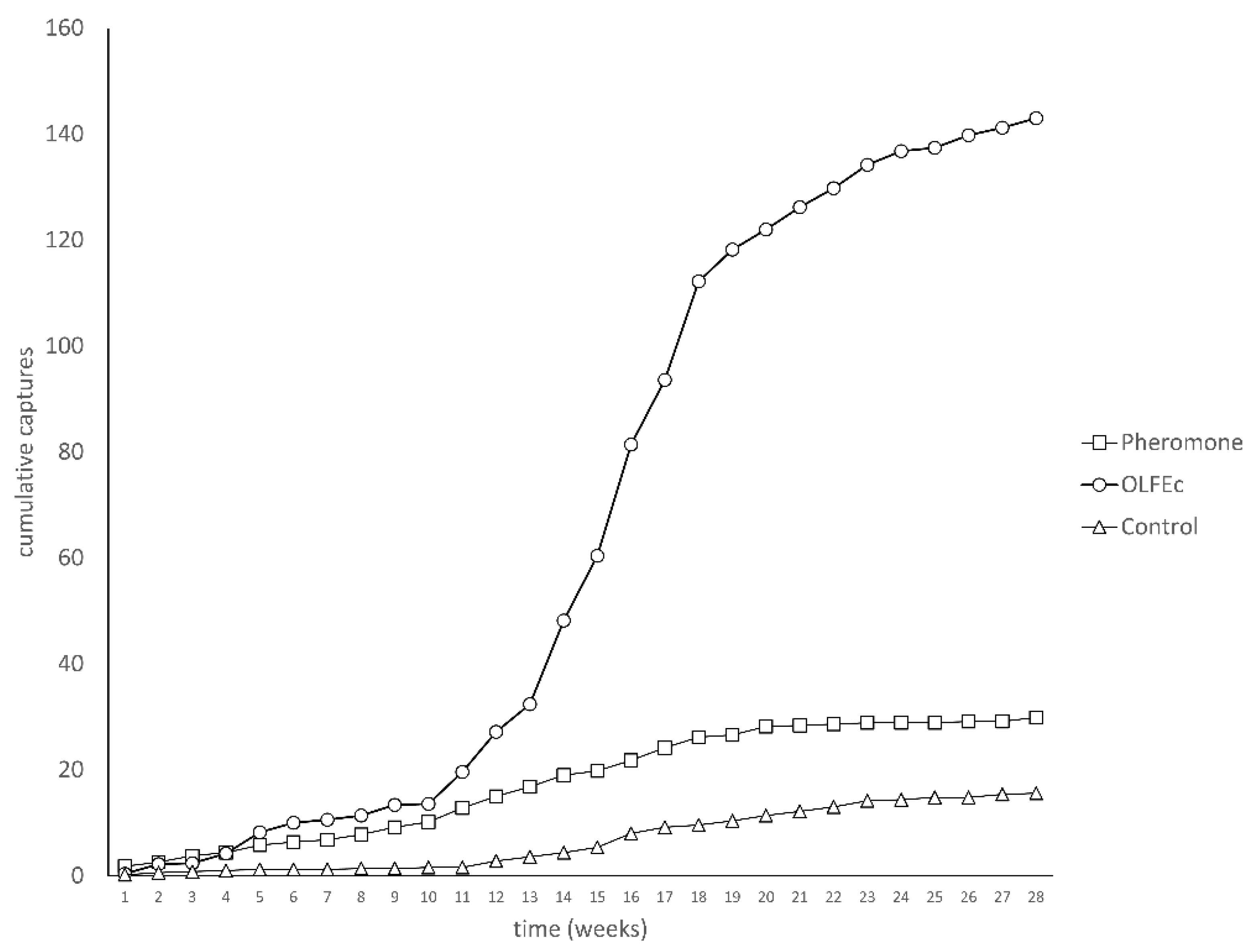
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Cocoa Organization (ICCO). Pest and Diseases. Cocoa Pod Borer (C. cramerella). 2015. Available online: http://www.icco.org/about/pest.aspx (accessed on 21 July 2019).
- Vos, J.G.M.; Ritchie, B.J.; Flood, J. Discovery Learning about Cacao: An Inspirational Guide for Training Facilitators; CABI Bioscience: Surrey, UK, 2003. [Google Scholar]
- Rosmana, A.; Daha, L.; Ridayani, A.; Gassa, A.; Urling, A. Persistency and penetration of entomopathogenic nematode Steinernema carpocapsae on surface of cocoa pod and its infectivity to cocoa pod borer Conopomorpha cramerella (Lepidoptera, Gracillariidae). Agritrop 2009, 28, 161–168. [Google Scholar]
- Lim, G.T.; Pan, K.Y. Observations on the Sexual Activity and Egg Production of Cocoa Pod Borer Conopomorpha Cramerella (Snellen) in the Laboratory; Ann Res Report; Department of Agriculture: Kota Kinabalu, Malaysia, 1986. [Google Scholar]
- See, Y.A.; Koo, K.C. Influence of Dolichoderus thoracicus (Hymenoptera: Formicidae) on cocoa pod damage by Conopomorpha cramerella (Lepidoptera: Gracillariidae) in Malaysia. Bull. Entomol. Res. 1996, 86, 467–474. [Google Scholar] [CrossRef]
- Vanialingam, T.; Easaw, P.T.; Irshad, M.; Manals, J.; Cruz, P. Early work in the control of the cocoa pod borer Acrocercops cramerella in the Philippines. In Proceedings of the 8th International Cocoa Research Conference, Cartagena, Colombia, 18–23 October 1981; pp. 345–351. [Google Scholar]
- Youdeowei, A. Review of Cocoa Pests in Indonesia and the Philippines and the development of research strategies for cocoa pest management. In Proceedings of the Developing Bulk Cocoa in Indonesia, Report on a Visit, Mimeo, Jakarta, Indonesia, 7 February–14 March 1980; pp. 1–64. [Google Scholar]
- Lim, G.T. Biology, ecology and control of cocoa pod borer Conopomorpha cramerella (Snellen). In Cocoa Pest and Disease Management in Southeast Asia and Australia; FAO Plant Production and Protection Paper; Keane, P.J., Putter, C.A.J., Eds.; FAO: Rome, Italy, 1992; Volume 22, pp. 85–100. [Google Scholar]
- Wood, B.J.; Fee, C.G.; Cheong, S.S.; Foh, C.C. Trials on control of the cocoa pod borer Conopomorpha cramerella (Snellen) in Sabah by regular complete harvesting. Trop. Pest Manag. 1992, 38, 271–278. [Google Scholar] [CrossRef]
- Beevor, P.S.; Mumford, J.D.; Shah, S.; Day, R.K.; Hall, D.R. Observations on pheromone-baited mass trapping for control of cocoa pod borer, Conopomorpha cramerella, in Sabah, East Malaysia. Crop Prot. 1993, 12, 134–140. [Google Scholar] [CrossRef]
- Teh, C.-L.; Panga, J.T.-Y.; Ho, C.-T. Variation of the response of clonal cocoa to attack by cocoa pod borer Conopomorpha cramerella (Lepidoptera: Gracillariidae) in Sabah. Crop Prot. 2006, 25, 712–717. [Google Scholar] [CrossRef]
- Matlick, B.K. Machete Technology: What Small Cocoa Farmers Need! Review of Pod Borer Control Methods by Small Farmers in Indonesia. In International Workshop on Sustainable Cocoa Growing; Smithsonian Tropical Research Institute: Panama, Republic of Panama, 29 March to 3 April 1998; Available online: https://nationalzoo.si.edu/scbi/migratorybirds/research/cacao/matlick.cfm (accessed on 21 July 2019).
- Zhang, A.; Kuang, L.F.; Maisin, N.; Karmuru, B.; Hall, D.R.; Virdiana, I.; Amber, S.; Purung, H.B.; Wang, S.; Hebbar, P. Activity of cocoa pod borer sex pheromone in cocoa fields. Environ. Entomol. 2008, 37, 719–724. [Google Scholar] [CrossRef]
- Niogret, J.; Ekayant, I.A.; Zhang, A. Sex pheromone of cocoa pod borer, Conopomorpha cramerella: Field activity evaluation of pheromone formulations in an Indonesian plantation. Insects 2022, 13, 663. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.J.A.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; Loon, J.J.A.; Dicke, M. Insect Plant Biology; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Cha, D.H.; Linn, C.E.; Teal, P.E.A.; Zhang, A.; Roelofs, W.L.; Loeb, G.M. Eavesdropping on plant volatiles by a specialist moth: Significance of ratio and concentration. PLoS ONE 2011, 6, e17033. [Google Scholar] [CrossRef]
- Preti, M.; Knight, A.L.; Favaro, R.; Basoalto, E.; Tasin, M.; Angeli, S. Comparison of New Kairomone-based lures for Cydia pomonella (Lepidoptera: Tortricidae) in Italy and USA. Insects 2021, 12, 72. [Google Scholar] [CrossRef]
- Knight, A.L.; Preti, M.; Basoalto, E.; Mujica, M.V.; Favaro, R.; Angeli, S. Combining female removal with mating disruption for management of Cydia pomonella in apple. Entomol. Gen. 2022, 42, 309–321. [Google Scholar] [CrossRef]
- Kendra, P.E.; Montgomery, W.S.; Deyrup, M.A.; Wakarchuk, D. Improved lure for redbay ambrosia beetle developed by enrichment of α-copaene content. J. Pest Sci. 2016, 89, 427–438. [Google Scholar] [CrossRef]
- Torto, B.; Suazo, A.; Alborn, H.; Tumlinson, J.; Teal, P.E.A. Response of the small hive beetle (Aethina tumida) to a blend of chemicals identified from honeybee (Apis mellifera) volatiles. Apidology 2005, 36, 523–532. [Google Scholar] [CrossRef]
- Torto, B.; Arbogast, R.T.; Alborn, H.; Suazo, A.; Engelsdrop, D.V.; Boucias, D.; Tumlinson, J.H.; Teal, P.E.A. Composition of volatiles from fermenting pollen dough and attractiveness to the small hive beetle Aethina tumida, a parasite of the honeybee Apis mellifera. Apidology 2007, 38, 380–389. [Google Scholar] [CrossRef]
- Torto, B.; Boucias, D.G.; Arbogast, R.T.; Tumlinson, J.H.; Teal, P.E. Multitrophic interaction facilitates parasite-host relationship between an invasive beetle and the honeybee. Proc. Natl. Acad. Sci. USA 2007, 104, 8374–8378. [Google Scholar] [CrossRef]
- Thwaite, W.G.; Mooney, A.M.; Eslick, M.A.; Nichol, H.I. Evaluating pear ester lures for monitoring Cydia pomonella (L.) (Lepidoptera: Tortricidae) in Granny Smith apples under mating disruption. Gen. Appl. Entomol. 2004, 33, 55–60. [Google Scholar]
- Kutinkova, H.; Subchev, M.; Light, S.; Lingren, B. Interactive effects of ethyl (2E,4Z)-2,4-decadienoate and sex pheromone lures to codling moth: Apple orchard investigations in Bulgaria. J. Plant Prot. Res. 2005, 45, 49–52. [Google Scholar]
- Landolt, P.J.; Suckling, D.M.; Judd, G.J.R. Positive interaction of a feeding attractant and a host kairomone for trapping the codling moth, Cydia pomonella (L.). J. Chem. Ecol. 2007, 33, 2236–2244. [Google Scholar] [CrossRef]
- Niogret, J.; Ekayanti, A.; Kendra, P.E.; Ingram, K.; Lambert, S.; Espky, N.D.; Marelli, J.-P. Host preferences of the cocoa pod borer, Conopomorpha cramerella, the main threat to cocoa production in Southeast Asia. Entomol. Exp. Applicata 2020, 168, 221–227. [Google Scholar] [CrossRef]
- Vanhove, W.; Vanhoudt, N.; Bhanu, K.R.M.; Abubeker, S.; Feng, Y.; Yu, M.; Van Damme, P.; Zhang, A. Geometric isomers of sex pheromone components do not affect attractancy of Conopomorpha cramerella in cocoa plantations. J. Appl. Entomol. 2015, 139, 660–668. [Google Scholar] [CrossRef]
- Bradley, J.D. Identify of the South-East Asian cocoa moth, Conopomorpha cramerella (Snellen) (Lepidoptera: Gracillariidae), with descriptions of three allied new species. Bull. Entomol. Res. 1986, 76, 41–51. [Google Scholar] [CrossRef]
- Knight, A.L.; Croft, B.A. Modeling and prediction technology. In Tortricid Pests; Van der Geest, L.P.S., Evenhuis, H.H., Eds.; Elsevier: Amsterdam, The Netherlands, 1991; pp. 301–312. [Google Scholar]
- Niogret, J.; Ekayanti, A.; Ingram, K.; Lambert, S.; Kendra, P.E.; Alborn, H.; Epsky, N.D. Development and behavioral ecology of Conopomorpha cramerella (Lepidoptera: Gracillariidae). Fla ENT 2019, 102, 382–387. [Google Scholar] [CrossRef]
- Leblanc, L.; Rubinoff, D.; Vargas, R.I. Attraction of non-target species to fruit fly (Diptera: Tephritidae) male lures and decaying fruit flies in traps in Hawaii. Environ. Entomol. 2009, 38, 1146–1461. [Google Scholar] [CrossRef] [Green Version]
- Beyaert, I.; Hilker, M. Plant odour plumes as mediators of plant-insect interactions. Biol. Rev. 2014, 89, 68–81. [Google Scholar] [CrossRef]
- Barrozo, R.B.; Gadenne, C.; Anton, S. Switching attraction to inhibition: Mating-induced reversed role of sex pheromone in an insect. J. Exp. Biol. 2010, 213, 2933–2939. [Google Scholar] [CrossRef]
- Barrozo, R.B.; Jarriault, D.; Deisig, N.; Gemeno, C.; Monsempes, C.; Lucas, P.; Gadenne, C.; Anton, S. Mating-induced differential coding of plant odour and sex pheromone in a male moth. Eur. J. Neurosci. 2011, 33, 1841–1850. [Google Scholar] [CrossRef]
- Kromann, S.H.; Saveer, A.M.; Binyameen, M.; Bengtsson, M.; Birgersson, G.; Hansson, B.S.; Schlyter, F.; Witzgall, P.; Ignell, R.; Becher, P.G. Concurrent modulation of neuronal and behavioural olfactory responses to sex and host plant cues in a male moth. Proc. R. Soc. B 2015, 282, 20141884. [Google Scholar] [CrossRef] [PubMed]
- Landolt, P.J.; Phillips, T.W. Host plant influences on sex pheromone behavior of phytophagous insects. Annu. Rev. Entomol. 1997, 42, 371–391. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Guerrero, A. Interactions of insect pheromones and plant semiochemicals. Trends Plant Sci. 2004, 9, 253–261. [Google Scholar] [CrossRef]
- Dickens, J.C.; Smith, J.W.; Light, D.M. Green leaf volatiles enhance sex attractant pheromone of the tobacco budworm, Heliothis virescens (Lepidoptera: Noctuidae). Chemoecology 1993, 4, 175–177. [Google Scholar] [CrossRef]
- Light, D.M.; Flath, R.A.; Buttery, R.G.; Zalom, F.G.; Rice, R.E.; Dickens, J.C.; Jang, E.B. Host-plant green-leaf volatiles synergize the synthetic sex pheromones of the corn earworm and codling moth (Lepidoptera). Chemoecol 1993, 4, 145–152. [Google Scholar] [CrossRef]
- Schmidt-Büsser, D.; Von Arx, M.; Guerin, P.M. Host plant volatiles serve to increase the response of male European grape berry moths, Eupoecilia ambiguella, to their sex pheromone. J. Comp. Physiol. A 2009, 195, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Varela, N.; Avilla, J.; Anton, S.; Gemeno, C. Synergism of pheromone and host plant volatile blends in the attraction of Grapholita molesta males. Entomol. Exp. Appl. 2011, 141, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Von Arx, M.; Schmidt-Busser, D.; Guerin, P.M. Plant volatiles enhance behavioral responses of grapevine moth males, Lobesia botrana to sex pheromone. J. Chem. Ecol. 2012, 38, 222–225. [Google Scholar] [CrossRef]
- Niogret, J.; Mars Wrigley, Smithfield, QLD, Australia. Unpublished work. 2019.




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niogret, J.; Kendra, P.E.; Ekayanti, A.; Zhang, A.; Marelli, J.-P.; Tabanca, N.; Epsky, N. Development of a Kairomone-Based Attractant as a Monitoring Tool for the Cocoa Pod Borer, Conopomorpha cramerella (Snellen) (Lepidoptera: Gracillariidae). Insects 2022, 13, 813. https://doi.org/10.3390/insects13090813
Niogret J, Kendra PE, Ekayanti A, Zhang A, Marelli J-P, Tabanca N, Epsky N. Development of a Kairomone-Based Attractant as a Monitoring Tool for the Cocoa Pod Borer, Conopomorpha cramerella (Snellen) (Lepidoptera: Gracillariidae). Insects. 2022; 13(9):813. https://doi.org/10.3390/insects13090813
Chicago/Turabian StyleNiogret, Jerome, Paul E. Kendra, Arni Ekayanti, Aijun Zhang, Jean-Philippe Marelli, Nurhayat Tabanca, and Nancy Epsky. 2022. "Development of a Kairomone-Based Attractant as a Monitoring Tool for the Cocoa Pod Borer, Conopomorpha cramerella (Snellen) (Lepidoptera: Gracillariidae)" Insects 13, no. 9: 813. https://doi.org/10.3390/insects13090813