
Genomic-Phenomic Reciprocal Illumination: Desyopone hereon gen. et sp. nov., an Exceptional Aneuretine-like Fossil Ant from Ethiopian Amber (Hymenoptera: Formicidae: Ponerinae) †
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Geological Setting
2.2. Materials and Specimen Preparation
2.3. Microphotography
2.4. Synchrotron Micro-Computed Tomography
2.5. Data Segmentation and Rendering
2.6. Conceptual
2.7. Measurements and Indices
| BL | Body length: the total body length from the anterior margin of the head excluding mandibles to the apex of the abdomen, measured in dorsal view. |
| HL | Head length: the length of the head capsule excluding the mandibles; measured in full-face view in a straight line from a line that spans the anteriormost points of the clypeal lobes to the level of a line that spans the posterior corners of the head capsule. |
| HW | Head width: the maximum width of the head immediately behind the eyes, measured in full-face view. |
| HWE * | Head width, eyes: the maximum width of the head, including the compound eyes. |
| SL | Scape length: the maximum straight-line length of the scape, excluding the basal constriction or neck that occurs just distad the condylar bulb. |
| EL | Eye length: in profile, the maximum measurable length of the compound eye. |
| OLL * | Ocellus length, lateral: the maximum length of the lateral ocellus, measured in full-face view. |
| OIL * | Inter-ocellus length: the minimum distance between the lateral ocelli, measured in full-face view. |
| WL | Weber’s length: the diagonal length of the mesosoma in profile, from the angle at which the pronotum meets the cervix to the posterior basal angle of the metapleuron. |
| ML * | Mesoscutum length: the maximum length of the mesoscutum, measured in dorsal view. |
| MW * | Mesoscutum width: the maximum width of the mesoscutum, measured in dorsal view. |
| FWL * | Forewing length: the maximum length of the forewing from the apices of the axillary sclerites to the wing apex. |
| PH | Petiole height: the maximum height of petiole (abdominal segment 2), measured in profile view. |
| PL | Petiole length: the maximum length of petiole (abdominal segment 2), measured in dorsal view. |
| PW | Petiole width: the maximum width of petiole (abdominal segment 2), measured in dorsal view. |
| GL | Gaster length: the maximum length of gaster (abdominal tergites 3 to 7), measured in dorsal view. |
| GW | Gaster width: the maximum width of gaster, measured in dorsal view. |
| CI | Cephalic index: HW/HL X 100. |
| HWI * | Head width index: HW/HWE X 100. |
| SI | Scape index: SL/HW X 100. |
| OI | Ocular index: EL/HL X 100. |
| OCI * | Ocellar index: OLL/OIL X 100. |
| EPI | Eye Position Index: in full-face view, the distance from a horizontal line that spans the anterior clypeal margin to one that spans the anterior margins of the eyes, divided by the distance from a horizontal line that spans the posterior margins of the eyes to one that spans the posterior corners of the head, X 100. |
| MI* | Mesoscutum index: MW/ML X 100. |
| PI | Petiolar index: PL/PH X 100. |
2.8. Repositories
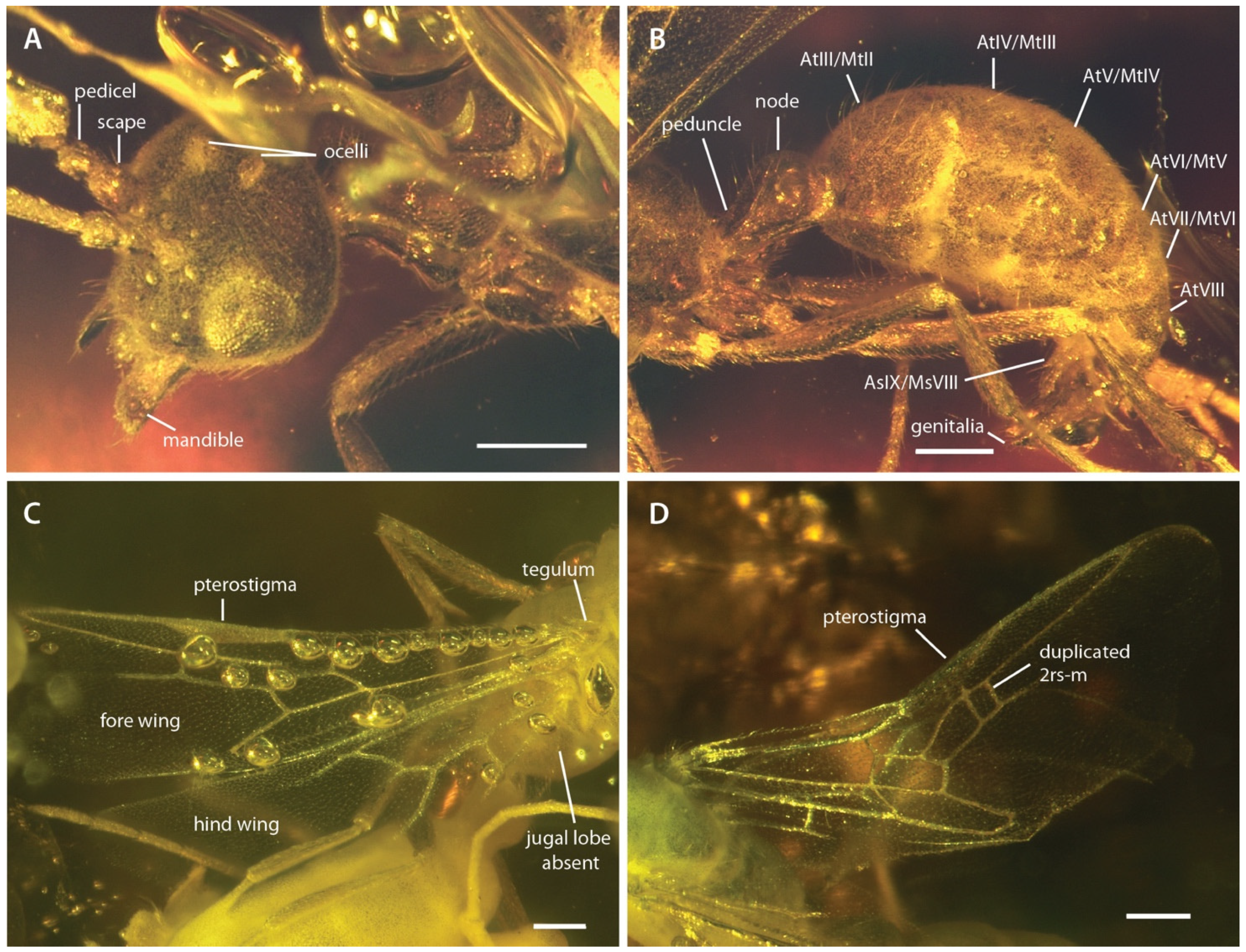
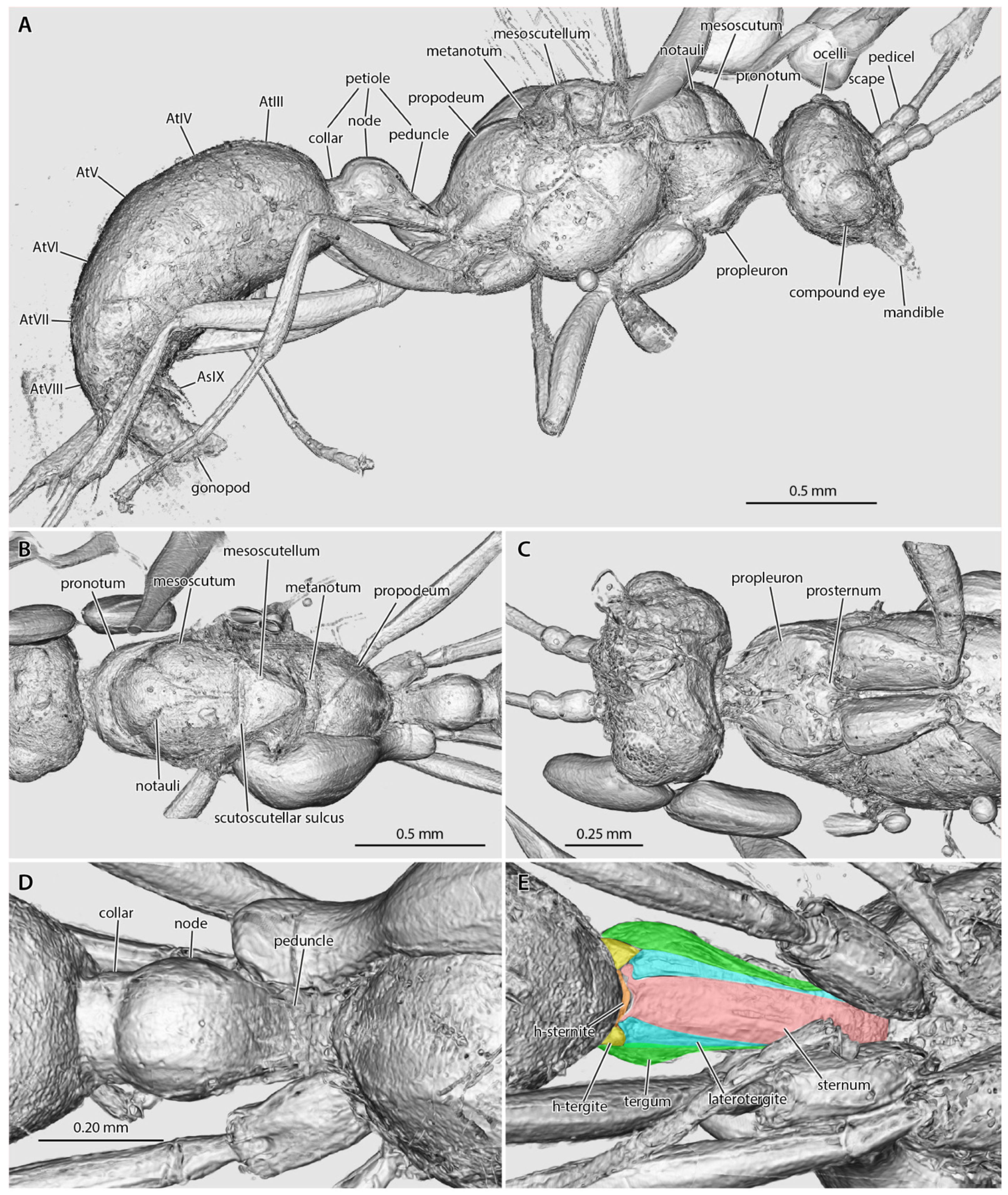
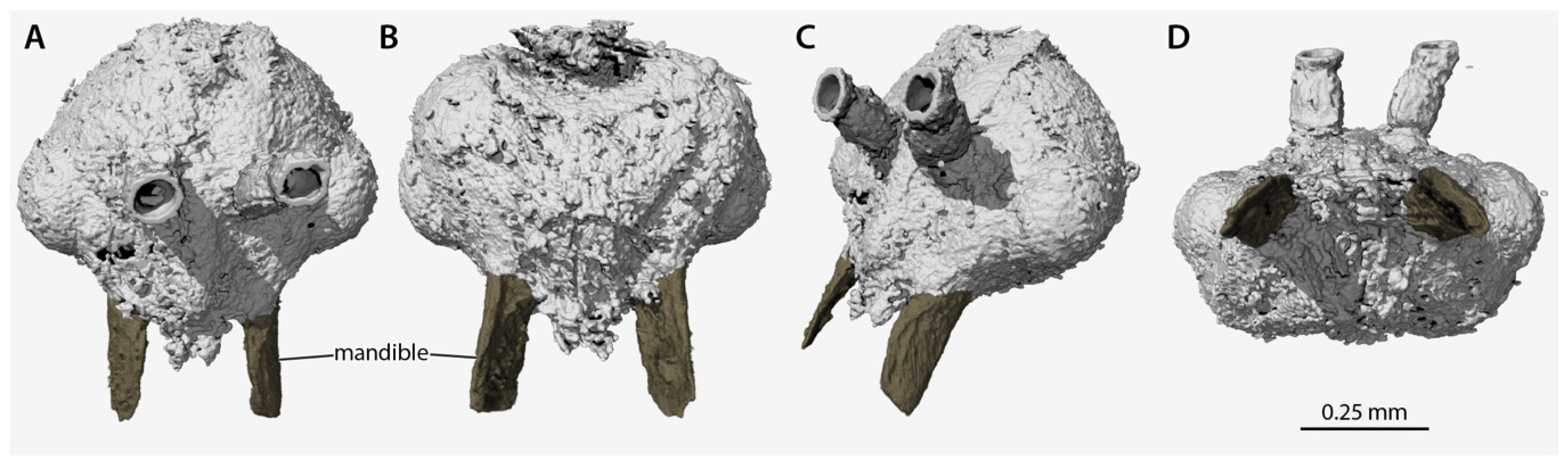
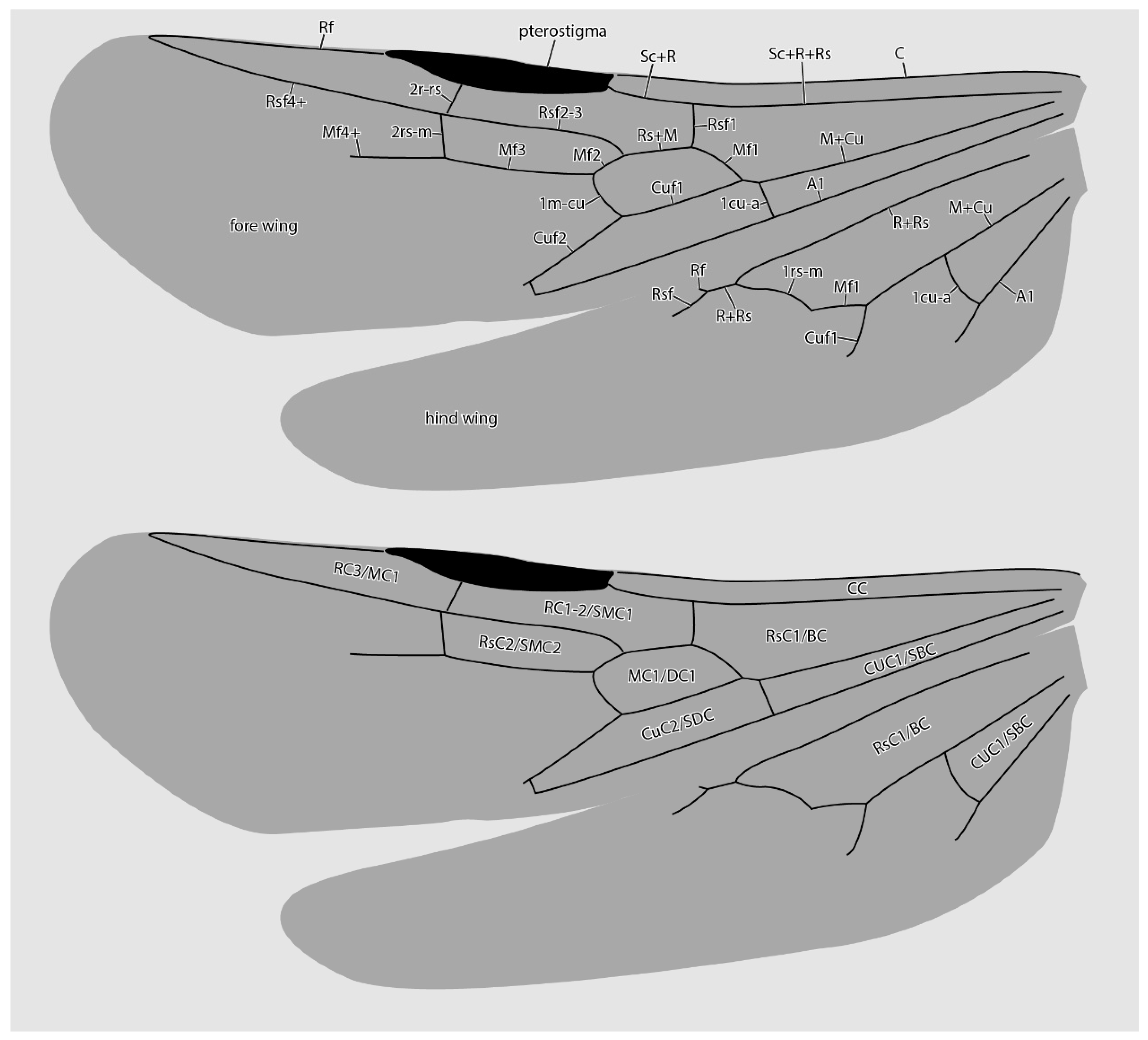
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bolton, B. AntCat, an Online Catalog of the Ants of the World by Barry Bolton. Available online: https://antcat.org (accessed on 28 June 2022).
- Barden, P. Fossil ants (Hymenoptera: Formicidae): Ancient diversity and the rise of modern lineages. Myrmecol. News 2017, 24, 1–30. [Google Scholar]
- LaPolla, J.S.; Dlussky, G.M.; Perrichot, V. Ants and the fossil record. Ann. Rev. Entomol. 2013, 58, 609–630. [Google Scholar] [CrossRef] [PubMed]
- Boudinot, B.E.; Perrichot, V.; Chaul, J.C.M. † Camelosphecia gen. nov., lost ant-wasp intermediates from the mid-Cretaceous (Hymenoptera, Formicoidea). ZooKeys 2020, 1005, 21–55. [Google Scholar] [CrossRef] [PubMed]
- Heer, O. Die Insektenfauna der Tertiärgebilde von Oeningen und von Radoboj in Croatien. Zweiter Theil: Heuschrecken, Florfliegen, Aderflüger, Schmetterlinge und Fliegen; W. Engelmann Publisher: Leipzig, Germany, 1849; p. 264. [Google Scholar]
- Mayr, G. Die Ameisen des baltischen Bernsteins. Beitr. Naturk. Preuss 1868, 1, 1–102. [Google Scholar]
- Wheeler, W.M. The ants of the Baltic Amber. Schrift. Physikal. Ökonom. Gesell. Königsberg 1915, 55, 1–142. [Google Scholar]
- Carpenter, F.M. The fossil ants of North America. Bull. Mus. Comp. Zool. 1930, 70, 1–66. [Google Scholar]
- Théobald, N. Les Insectes Fossiles des Terrains Oligocènes de France; G. Thomas: Nancy, France, 1937; p. 473. [Google Scholar]
- Hong, Y.-C. Amber Insects of China; Beijing Science and Technology Press: Beijing, China, 2002; p. 653. (In Chinese) [Google Scholar]
- Bolton, B. Synopsis and classification of Formicidae. Mem. Amer. Entomol. Inst. 2003, 71, 1–370. [Google Scholar]
- Bolton, B. Identification Guide to the Ant Genera of the World; Harvard University Press: Cambridge, MA, USA, 1994; p. 222. [Google Scholar]
- Moreau, C.S.; Bell, C.D.; Vila, R.; Archibald, S.B.; Pierce, N.E. Phylogeny of the ants: Diversification in the age of angiosperms. Science 2006, 312, 101–104. [Google Scholar] [CrossRef]
- Brady, S.G.; Schultz, T.R.; Fisher, B.L.; Ward, P.S. Evaluating alternative hypotheses for the early evolution and diversification of ants. Proc. Natl. Acad. Sci. USA 2006, 103, 18172–18177. [Google Scholar] [CrossRef]
- Borowiec, M.L.; Rabeling, C.; Brady, S.G.; Fisher, B.L.; Schultz, T.R.; Ward, P.S. Compositional heterogeneity and outgroup choice influence the internal phylogeny of the ants. Mol. Phylogenet. Evol. 2019, 134, 111–121. [Google Scholar] [CrossRef]
- Branstetter, M.G.; Longino, J.T.; Ward, P.S.; Faircloth, B.C. Enriching the ant tree of life: Enhanced UCE bait set for genome-scale phylogenetics of ants and other Hymenoptera. Meth. Ecol. Evol. 2017, 8, 768–776. [Google Scholar] [CrossRef]
- Romiguier, J.; Borowiec, M.L.; Weyna, A.; Helleu, Q.; Loire, E.; La Mendola, C.; Rabeling, C.; Fisher, B.L.; Ward, P.S.; Keller, L. Ant phylogenomics reveals a natural selection hotspot preceding the origin of complex eusociality. Curr. Biol. 2022, 32, 2942–2947. [Google Scholar] [CrossRef]
- Marshall, C.R. Five palaeobiological laws needed to understand the evolution of the living biota. Nat. Ecol. Evol. 2017, 1, 0165. [Google Scholar] [CrossRef]
- Louca, S.; Pennell, M.W. Extant timetrees are consistent with a myriad of diversification histories. Nature 2020, 580, 502–505. [Google Scholar] [CrossRef]
- Wipfler, B.; Pohl, H.; Yavorskaya, M.I.; Beutel, R.G. A review of methods for analysing insect structures—The role of morphology in the age of phylogenomics. Curr. Opin. Insect Sci. 2016, 16, 60–68. [Google Scholar] [CrossRef]
- Schwermann, A.H.; dos Santos Rolo, T.; Caterino, M.S.; Bechly, G.; Schmied, H.; Baumbach, T.; van de Kamp, T. Preservation of three-dimensional anatomy in phosphatized fossil arthropods enriches evolutionary inferences. Elife 2016, 5, e12129. [Google Scholar] [CrossRef]
- Van de Kamp, T.; Schwermann, A.H.; dos Santos Rolo, T.; Lösel, P.D.; Engler, T.; Etter, W.; Faragó, T.; Göttlicher, J.; Heuveline, V.; Kopman, A.; et al. Parasitoid biology preserved in mineralized fossils. Nat. Commun. 2018, 9, 3325. [Google Scholar] [CrossRef]
- Barden, P.; Herhold, H.W.; Grimaldi, D.A. A new genus of hell ants from the Cretaceous (Hymenoptera: Formicidae: Haidomyrmecini) with a novel head structure. Syst. Entomol. 2017, 42, 837–846. [Google Scholar] [CrossRef]
- Boudinot, B.E.; Khouri, Z.; Richter, A.; Griebenow, Z.H.; van de Kamp, T.; Perrichot, V.; Barden, P. Evolution and systematics of the Aculeata and kin (Hymenoptera), with emphasis on the ants (Formicoidea: †@@@idae fam. nov., Formicidae). bioRxiv 2022, 1–450. [Google Scholar] [CrossRef]
- Perrichot, V.; Boudinot, B.E.; Engel, M.S.; Xu, C.; Bojarski, B.; Szwedo, J. Ants (Hymenoptera: Formicidae) from Miocene Ethiopian amber: Filling gaps in the geological record of African terrestrial biota. Zool. J. Linn. Soc. 2022. [Google Scholar] [CrossRef]
- Richter, A.; Boudinot, B.E.; Yamamoto, S.; Katzke, J.; Beutel, R.G. The first reconstruction of the head anatomy of a Cretaceous insect, †Gerontoformica gracilis (Hymenoptera, Formicidae), and the early evolution of the ants. Insect Syst. Divers. in press.
- Bouju, V.; Feldberg, K.; Kaasalainen, U.; Schäfer-Verwimp, A.; Hedenäs, L.; Buck, W.R.; Wang, B.; Perrichot, V.; Schmidt, A.R. Miocene Ethiopian amber: A new source of fossil cryptogams. J. Syst. Evol. 2021. [Google Scholar] [CrossRef]
- Bouju, V.; Rosse-Guillevic, S.; Griffon, M.; Bojarski, B.; Szwedo, J.; Perrichot, V. The genus Allodia (Diptera: Mycetophilidae) in Miocene Ethiopian amber. Foss. Rec. 2021, 24, 339–346. [Google Scholar] [CrossRef]
- Belay, T.; Tesfay, I.; Ayalew, A.; Yohannes, G.; Zewdie, T.; Bekele, H.; Tadesse, M.; Demisse, T.; Alemu, T. Geology of the Were-Ilu area, with accompanying geological map (NC 37-7). Geol. Surv. Ethiop. Mem. 2009, 25, 56. [Google Scholar]
- AntWeb Version 8.76.4. California Academy of Science. Available online: https://www.antweb.org (accessed on 8 June 2022).
- Haibel, A.; Ogurreck, M.; Beckmann, F.; Dose, T.; Wilde, F.; Herzen, J.; Müller, M.; Schreyer, A.; Nazmov, V.; Simon, M.; et al. Micro- and nano-tomography at the GKSS Imaging Beamline at PETRA III. Dev. X-Ray Tomogr. VII 2010, 7804, 98–105. [Google Scholar]
- Greving, I.; Wilde, F.; Ogurreck, M.; Herzen, J.; Hammel, J.U.; Hipp, A.; Friedrich, F.; Lottermoser, L.; Dose, T.; Burmester, H.; et al. P05 imaging beamline at PETRA III: First results. In Proceedings of the SPIE—Developments in X-Ray Tomography IX, San Diego, CA, USA, 18–20 August 2014; Volume 9212, p. 92120O-8. [Google Scholar]
- Wilde, F.; Ogurreck, M.; Greving, I.; Hammel, J.U.; Beckmann, F.; Hipp, A.; Lottermoser, L.; Khokhriakov, I.; Lytaev, P.; Dose, T.; et al. Micro-CT at the imaging beamline P05 at PETRA III. AIP Conf. Proc. 2016, 1741, 030035. [Google Scholar] [CrossRef]
- Moosmann, J.; Ershov, A.; Weinhardt, V.; Baumbach, T.; Prasad, M.S.; LaBonne, C.; Xiao, X.; Kashef, J.; Hoffmann, R. Time-lapse X-ray phase-contrast microtomography for In vivo imaging and analysis of morphogenesis. Nat. Protocol. 2014, 9, 294–304. [Google Scholar] [CrossRef]
- Palenstijn, W.J.; Batenburg, K.J.; Sijbers, J. Performance improvements for iterative electron tomography reconstruction using graphics processing units (GPUs). J. Struct. Biol. 2011, 176, 250–253. [Google Scholar] [CrossRef] [Green Version]
- Van Aarle, W.; Palenstijn, W.J.; de Beenhouwer, J.; Altantzis, T.; Bals, S.; Batenburg, K.J.; Sijbers, J. The ASTRA Toolbox: A platform for advanced algorithm development in electron tomography. Ultramicroscopy 2015, 157, 35–47. [Google Scholar] [CrossRef]
- Van Aarle, W.; Palenstijn, W.J.; Cant, J.; Janssens, E.; Bleichrodt, F.; Dabravolski, A.; de Beenhouwer, J.; Batenburg, K.J.; Sijbers, J. Fast and Flexible X-ray Tomography Using the ASTRA Toolbox. Optic. Expr. 2016, 24, 25129–25147. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Meth. 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Lösel, P.D.; van de Kamp, T.; Jayme, A.; Ershov, A.; Faragó, T.; Pichler, O.; Jerome, N.T.; Aadepu, N.; Bremer, S.; Chilingaryan, S.A.; et al. Introducing Biomedisa as an open-source online platform for biomedical image segmentation. Nat. Commun. 2020, 11, 5577. [Google Scholar] [CrossRef] [PubMed]
- Engelkes, K.; Friedrich, F.; Hammel, J.U.; Haas, A. A simple setup for episcopic microtomy and a digital image processing workflow to acquire high-quality volume data and 3D surface models of small vertebrates. Zoomorphology 2018, 137, 213–228. [Google Scholar] [CrossRef]
- Boudinot, B.E.; Moosdorf, O.T.D.; Beutel, R.G.; Richter, A. Anatomy and evolution of the head of Dorylus helvolus (Formicidae: Dorylinae): Patterns of sex- and caste-limited traits in the sausagefly and the driver ant. J. Morphol. 2021, 282, 1616–1658. [Google Scholar] [CrossRef] [PubMed]
- Boudinot, B.E. Contributions to the knowledge of Formicidae (Hymenoptera, Aculeata): A new diagnosis of the family, the first global male-based key to subfamilies, and a treatment of early branching lineages. Eur. J. Taxon. 2015, 120, 1–62. [Google Scholar] [CrossRef]
- Aibekova, L.; Boudinot, B.E.; Beutel, R.G.; Richter, A.; Keller, R.A.; Hita Garcia, F.; Economo, E.P. The skeletomuscular system of the mesosoma of Formica rufa workers (Hymenoptera: Formicidae). Insect Syst. Divers. 2022, 6, 1–26. [Google Scholar] [CrossRef]
- Lieberman, Z.E.; Billen, J.P.J.; van de Kamp, T.; Boudinot, B.E. The ant abdomen: The skeletomuscular and soft tissue anatomy of Amblyopone australis workers (Hymenoptera: Formicidae). J. Morphol. 2022, 283, 693–770. [Google Scholar] [CrossRef]
- Boudinot, B.E. A general theory of genital homologies for the Hexapoda (Pancrustacea) derived from skeletomuscular correspondences, with emphasis on the Endopterygota. Arthropod Struct. Dev. 2018, 47, 563–613. [Google Scholar] [CrossRef]
- Keller, R.A. A phylogenetic analysis of ant morphology (Hymenoptera: Formicidae) with special reference to the poneromorph subfamilies. Bull. Am. Mus. Nat. Hist. 2011, 355, 1–90. [Google Scholar] [CrossRef]
- Yoshimura, M.; Fisher, B.L. A revision of male ants of the Malagasy region (Hymenoptera: Formicidae): Key to subfamilies and treatment of the genera of Ponerinae. Zootaxa 2007, 1654, 21–40. [Google Scholar] [CrossRef]
- Ogata, K. A generic synopsis of the poneroid complex of the family Formicidae in Japan (Hymenoptera). Part 1. Subfamilies Ponerinae and Cerapachyinae. Esakia 1987, 25, 97–132. [Google Scholar] [CrossRef]
- Yoshimura, M.; Onoyama, K. Male-based keys to the subfamilies and genera of Japanese ants (Hymenoptera: Formicidae). Entomol. Sci. 2002, 5, 421–443. [Google Scholar]
- Branstetter, M.G.; Longino, J.T. UCE phylogenomics of New World Cryptopone (Hymenoptera: Formicidae) elucidates genus boundaries, species boundaries, and the vicariant history of a temperate–tropical disjunction. Insect Syst. Divers. 2022, 6, 1–23. [Google Scholar] [CrossRef]
- Wilson, E.O.; Eisner, T.; Wheeler, G.C.; Wheeler, J. Aneuretus simoni Emery, a major link in ant evolution. Bull. Mus. Comp. Zool. 1956, 115, 81–99. [Google Scholar]
- Griebenow, Z. Delimitation of tribes in the subfamily Leptanillinae (Hymenoptera: Formicidae), with a description of the male of Protanilla lini Terayama, 2009. Myrmecol. News 2020, 30, 229–250. [Google Scholar]
- Wilson, E.O. Ants of the Dominican amber (Hymenoptera: Formicidae). 3. The subfamily Dolichoderinae. Psyche 1985, 92, 17–37. [Google Scholar] [CrossRef]
- Wild, A.L.; Cuezzo, F. Rediscovery of a fossil dolichoderine ant lineage (Hymenoptera: Formicidae: Dolichoderinae) and a description of a new genus from South America. Zootaxa 2006, 1142, 57–68. [Google Scholar] [CrossRef]
- Fisher, B.L. Two new dolichoderine ant genera from Madagascar: Aptinoma gen. n. and Ravavy gen. n. (Hymenoptera: Formicidae). Zootaxa 2009, 2118, 37–52. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boudinot, B.E.; Richter, A.K.; Hammel, J.U.; Szwedo, J.; Bojarski, B.; Perrichot, V. Genomic-Phenomic Reciprocal Illumination: Desyopone hereon gen. et sp. nov., an Exceptional Aneuretine-like Fossil Ant from Ethiopian Amber (Hymenoptera: Formicidae: Ponerinae). Insects 2022, 13, 796. https://doi.org/10.3390/insects13090796
Boudinot BE, Richter AK, Hammel JU, Szwedo J, Bojarski B, Perrichot V. Genomic-Phenomic Reciprocal Illumination: Desyopone hereon gen. et sp. nov., an Exceptional Aneuretine-like Fossil Ant from Ethiopian Amber (Hymenoptera: Formicidae: Ponerinae). Insects. 2022; 13(9):796. https://doi.org/10.3390/insects13090796
Chicago/Turabian StyleBoudinot, Brendon E., Adrian K. Richter, Jörg U. Hammel, Jacek Szwedo, Błażej Bojarski, and Vincent Perrichot. 2022. "Genomic-Phenomic Reciprocal Illumination: Desyopone hereon gen. et sp. nov., an Exceptional Aneuretine-like Fossil Ant from Ethiopian Amber (Hymenoptera: Formicidae: Ponerinae)" Insects 13, no. 9: 796. https://doi.org/10.3390/insects13090796