1. Introduction
The bagrada bug,
Bagrada hilaris (Burmeister), is an invasive stink bug, native to Africa, Central Asia, and the Middle East [
1,
2,
3]. It was recorded for the first time in the US in 2008 in California and since has expanded its distribution rapidly to southwestern US states, Hawaii, and Mexico [
4,
5,
6,
7]. Its presence has also been reported in Mediterranean Europe (Pantelleria Island, Italy, and Malta), Australia, Chile, and Southeast Asia [
8,
9]. It primarily feeds on Brassicaceae species, though it is known to have a relatively large host range, feeding and damaging agriculturally important food crops including corn (
Zea mays L.; Poales: Poaceae), kale (
Brassica oleracea L.; Brassicales: Brassicaceae), arugula (
Eruca vescicaria (L.) Cav., Brassicales: Brassicaceae), sunflower (
Helianthus annuus L.; Asterales: Asteraceae), among others [
5,
10], and caper plants (
Capparis spinosa L.; Brassicales: Capparaceae) in southern Europe [
11]. The pest’s impact is particularly severe when large numbers of aggregating nymphal stages induce excessive feeding damage by causing the destruction of apical meristems, young leaves, and terminal growing points [
12]. There are few control strategies available to adequately suppress damaging populations of
B. hilaris [
13]. In the US, chemical control with pyrethroids and neo-nicotinoids demonstrated some impact, but the necessary multiple applications of these broad-spectrum insecticides have negative consequences for the economy and the environment [
5,
14]. Several egg parasitoid species, belonging to the genera
Trissolcus,
Gryon, and
Ooencyrtus, are under consideration as potential agents in classical biological control programs against bagrada bugs [
15,
16,
17]. These parasitoid species have been detected in Pakistan using sentinel eggs, an extremely valid biocontrol tactic based on exposing a consistent number of eggs of the target pest species for an amount of time suitable to obtain the egg parasitoid oviposition [
17].
The sterile insect technique (SIT) is a species-specific pest control method based on the mass rearing, sterilization, and inundative releases of sterile insects (generally males) of the same pest species [
18]. Irradiation, such as with gamma rays and X-rays, is used to sterilize mass-reared insects so that, while they remain sexually competitive, they cannot produce offspring [
19,
20,
21]. Wild females that mate with sterile males do not produce offspring, thus preventing the growth of the pest population [
18,
19]. It is considered among the most environmentally friendly insect pest control method ever developed and there are no documented off-target effects reported [
18,
19].
However, the irradiation biology of Hemiptera remains to be fully studied. Currently, there are no field applications of SIT, mainly because the release of sterile phytophagous hemipteran adults of widespread highly invasive pest species could cause unwanted damage to host crop species [
20]. Nevertheless, the opportune application of an SIT program is capable of achieving suppression and even eradication in particular geographic and/or infrastructural conditions (e.g., greenhouses, siloes, and isolated crop areas) if certain conditions related to the target pest biology (multivoltine, gregarious, suitable mating behavior, in the case of
B. hilaris) and its response to the irradiation are met [
21]. Moreover, for many hemipteran pest species, the use of irradiation to induce egg sterility can be applied in synergy with a classic biological control approach, based upon the use of co-evolved egg parasitoids. Recent data [
22] has confirmed that sterile pentatomid eggs are a suitable substrate for the oviposition and full larval development of their egg parasitoids.
Interest in evaluating the feasibility of applying irradiation techniques in support of classic biological control to manage
B. hilaris arose from the concept that classical biological control strategies for managing pentatomid pests are focused on the use of egg parasitoids [
23,
24,
25,
26]. Egg parasitoids, both in the area of origin of the target pest species or in the new environment, are detected and monitored by using newly oviposited sentinel eggs (lasting just 72 h in the field [
25]) or frozen sentinel eggs (lasting 3–4 days [
26]). The short exposition time is not the only limiting factor: manipulated eggs are less attractive and the distribution of live fertile eggs of pentatomid pest species in the new environment can be considered ethically incorrect and can unintentionally increase the dispersal of the target pest. This work should be considered the first step in the development of a new concept of “sterile sentinel eggs”, aimed at verifying if sterile eggs obtained by mating irradiated males with fertile females can be used in a classical biological control context to improve the management of this pest in a synergic manner [
27,
28].
This can be particularly true for
B. hilaris, a pentatomid species that exhibits the unusual behavior of ovipositing in soil, which differs from all other pentatomid species [
29,
30]. For this reason, special emphasis has been placed on the scelionid egg parasitoid
Gryon aethirium Talamas (formerly
G. gonikopalense (Sharma)) [
31], which is able to detect and oviposit on buried bagrada eggs [
15,
29].
Recent experiments evaluated the effect of irradiation on other invasive pentatomid pests, including the effects of irradiation on the biology of the newly emerged males of the brown marmorated stink bug (BMSB),
Halyomorpha halys Stål [
32], and the effects of high irradiation doses on the ultrastructure of the southern green stink bug (SGSB),
Nezara viridula L. [
33].
The central research questions raised in this study are how effective gamma irradiation is in inducing sterility in both sexes of bagrada and what effects increasing irradiation doses have on its longevity, fecundity (number of eggs oviposited by irradiated females), and fertility (number of hatched eggs in comparison with the number of eggs oviposited). This current research is the initial step of a wider program seeking to assess the feasibility of an SIT approach in synergy with classic biological control for a strategic plan for suppressing B. hilaris on the Italian island of Pantelleria.
4. Discussion
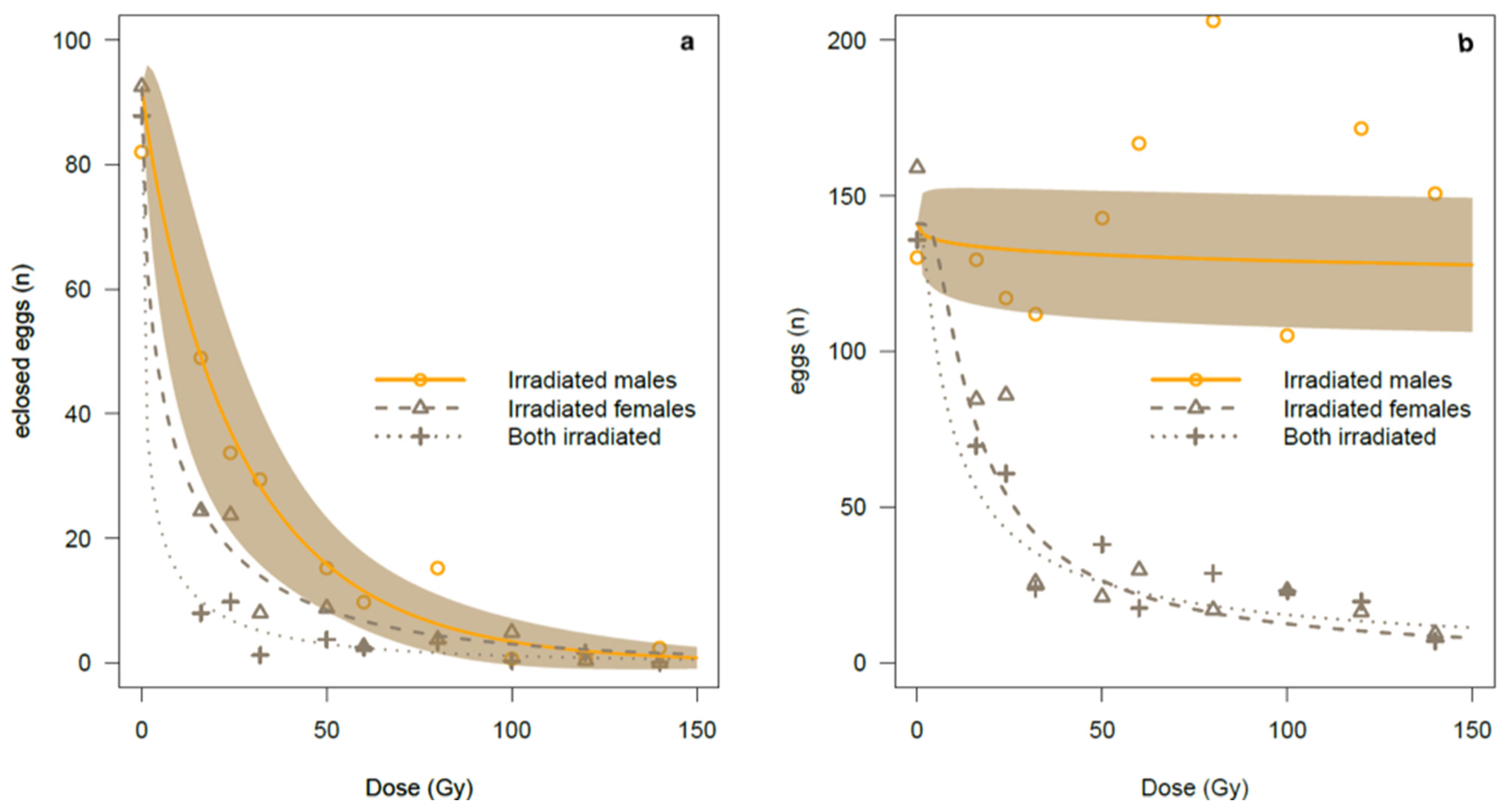
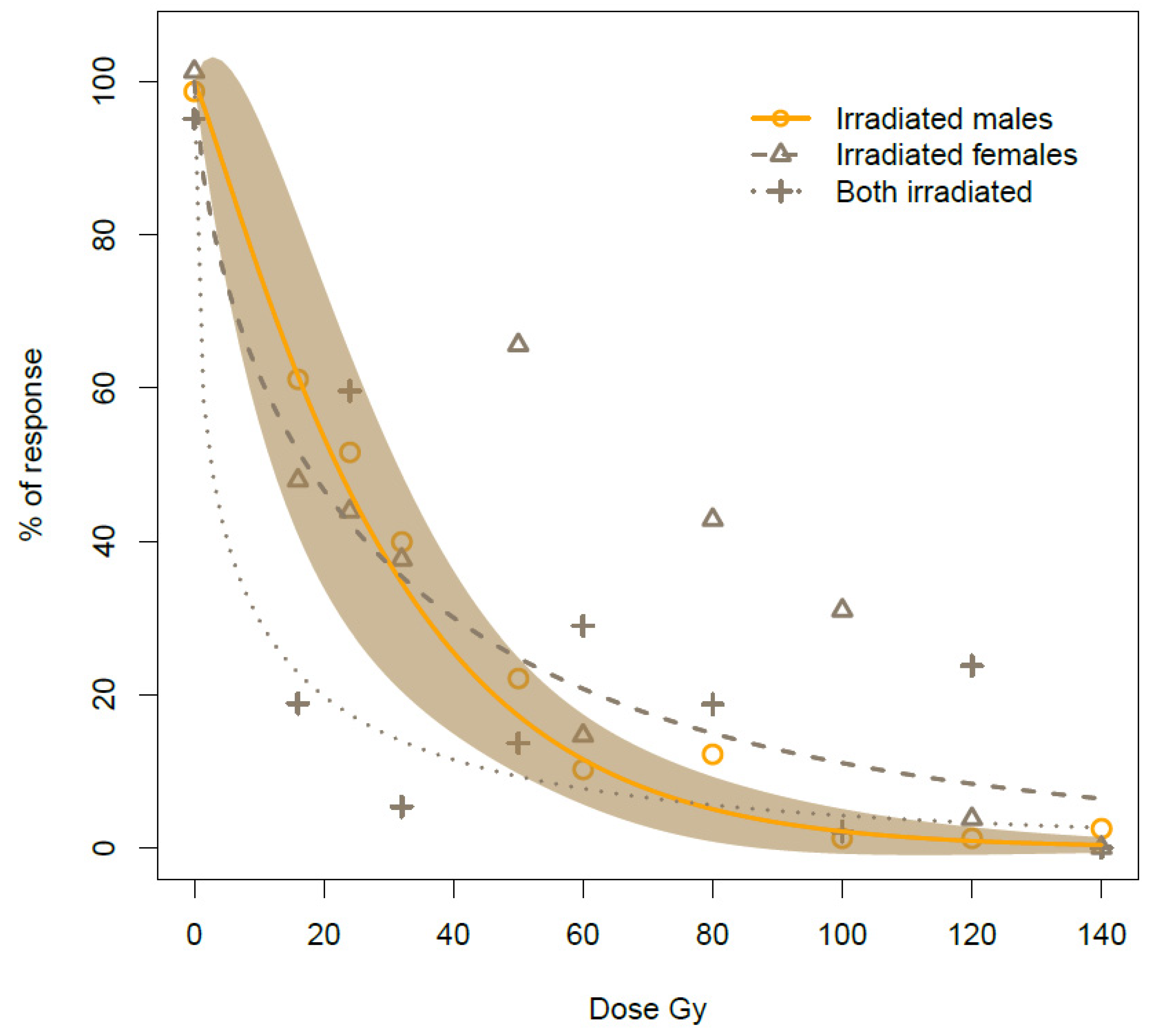
By gradually increasing the irradiation doses on males in the study, we observed a negative impact on reproduction, with almost complete female sterility at the highest dose. As such, male irradiation at 22 Gy induced 50% of estimated sterility on newly emerged bagrada bug adults while 90% sterility was achieved by irradiating newly emerged males with an estimated dose of 64 Gy (
Table 4). In addition, we observed almost complete sterility with doses starting from 100 Gy upwards, resulting in 99% egg sterility at this dose. However, we observed occasional egg eclosion with the first instar nymphs unable to move and feed, resulting in death within three days.
Obtaining sterile eggs from irradiated males and/or females provides a key element that can be used in synergy with a classic biological control approach, based upon the use of co-evolved egg parasitoids, as recently shown with the brown marmorated stinkbug [
22]. This approach supports the new concept of developing a small-scale biofactory for the production of sterile sentinel eggs, to be used to detect new egg parasitoids and to monitor the dispersal of the species released to control
B. hilaris in the new environment. Knowing the optimal irradiation doses for bagrada adults may help in designing future experimental SIT approaches. We acknowledge that this is a very premature stage of an SIT approach, but our study draws the line on how best to guarantee the 100% sterility of an irradiated cohort of bagrada bugs in case we envision mass rearing, irradiating, and releasing in open field conditions.
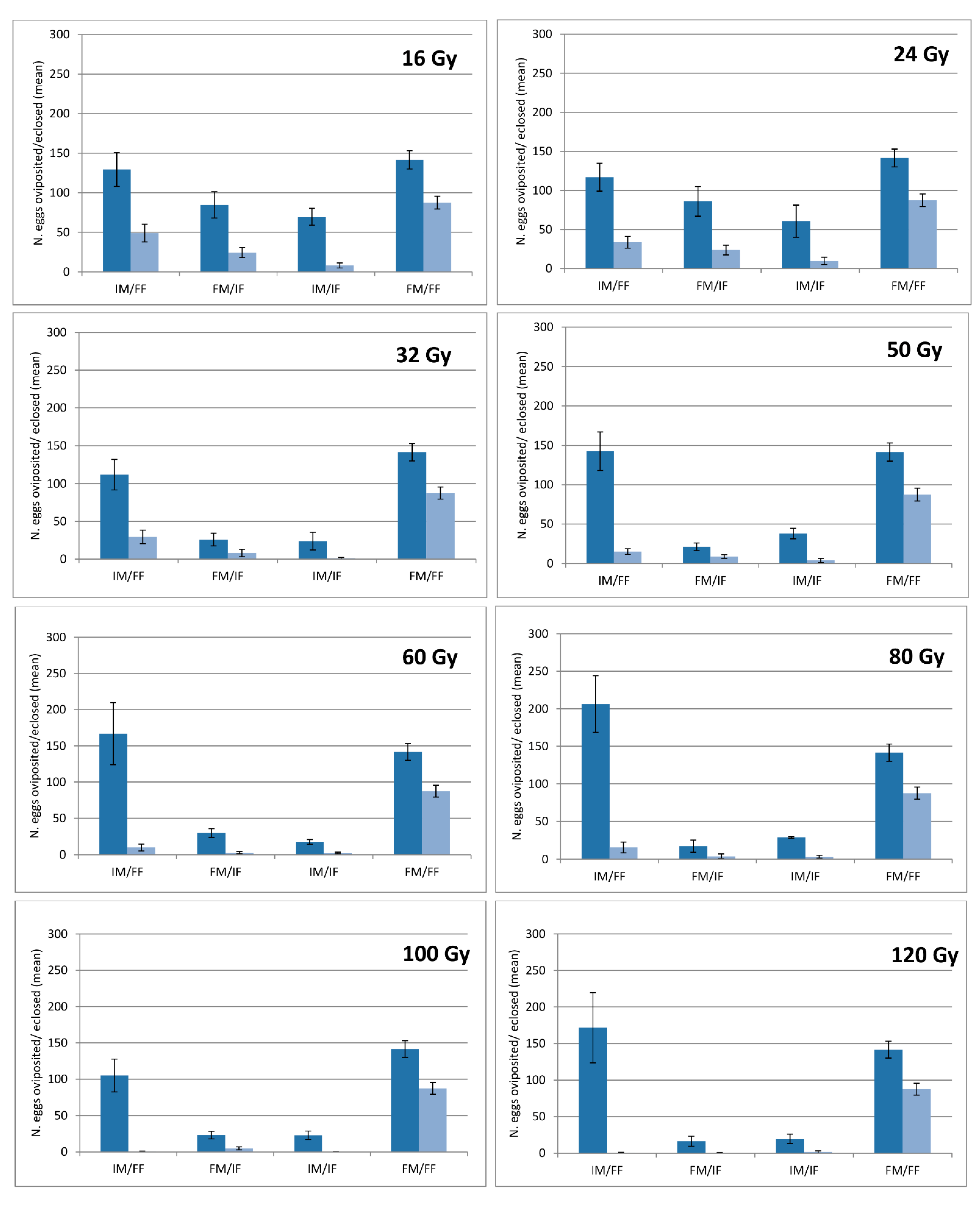
Interestingly, the effects of irradiation on reproduction were slightly different between males and females, not in terms of fertility but in terms of fecundity: the number of eggs oviposited by the irradiated females was negatively affected by the irradiation dose, declining from 60% at 50 Gy up to 80% at 100 Gy, whatever the male status (fertile or irradiated).
Conversely, the sperm of irradiated males did not have any significant effect on the female fecundity. The reproductive sterility is typically induced by exposure to X-rays, electron beams, or gamma rays from a
60Co or
137Cs source [
43], which all cause chromosomal damage. Sterility is usually permanent, although the irradiated males of some species may, over time, regain at least partial fertility, especially following multiple matings [
19,
44]. Irradiation doses suitable for an SIT program that sterilizes males do not induce immobility and/or physiological deformations in the sperm [
45]. However, in irradiated females, they can destroy oogonial cells, but the radiation sensitivity of oocytes varies with such factors as maturity and meiotic stage. As a result, the females of some species may retain a degree of fertility after irradiation, especially when treated late in ovarian development [
20].
In contrast to females, the mating competitiveness of sterile males is a function of their mating propensity and mating compatibility and is a crucial component of the sterile insect quality. Irradiation could be responsible for reducing insect quality in some cases: thus, the ability of released sterile males to compete for mating is critical [
20].
Different from what was observed in
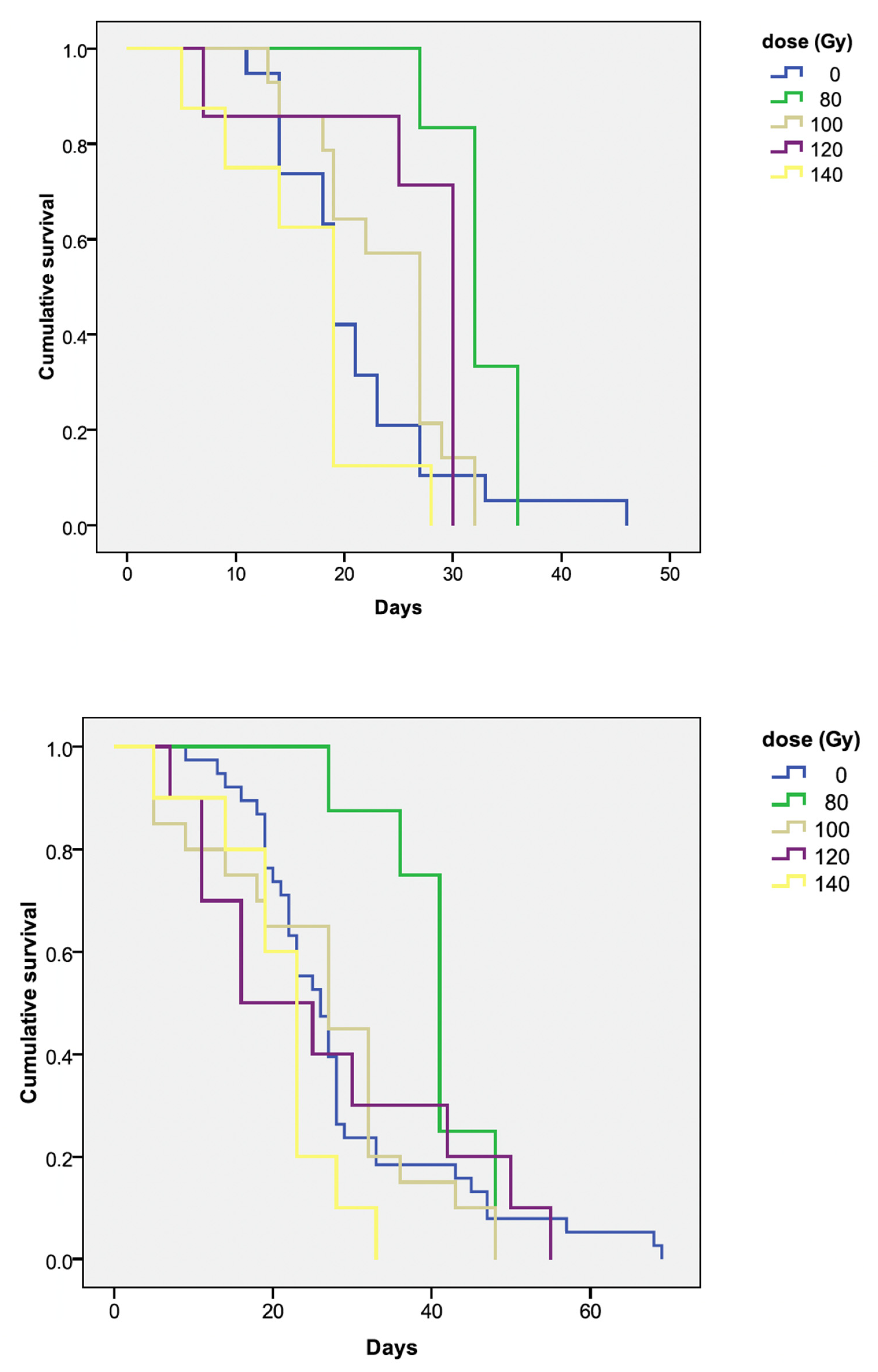
H. halys, where longevity declined significantly at irradiation doses higher than 24 Gy [
21,
29], irradiation even at the four highest doses used did not interfere with the longevity of
B. hilaris. The survival rates of the irradiated adults exposed to 100, 120, or 140 Gy did not differ from the control, whereas male longevity was apparently greater at 80 Gy in comparison with the untreated individuals. If this is applied in a wider approach with releases of irradiated bagrada males, then we can consider that either a stable or increased longevity may favor the success of an SIT approach. In fact, sterile males will remain in the environment for a long period of time, mating with fertile wild-type females to deposit sterile eggs over the course of the vegetative growth of the target crop species to protect, e.g., cole crops, caper plants, or rooibos tea. In addition to stopping or reducing the regeneration of bagrada in the environment, the presence of sterile eggs may attract resident or intentionally released egg parasitoids. This double action of irradiation and biological control needs to be further studied with field trials.
There is not a “perfect” target pest eligible for the SIT [
46]. The history of the SIT has been focused mainly on two orders: Diptera and Lepidoptera [
47]. Other orders and families, such as Hemiptera and Pentatomidae, were disregarded from SIT applications. The fact that the feeding behavior of (sterile) adults induces important economic damage to their host plant crops always remained an issue [
19]. However, this trend changed some years ago, through the preliminary screening of the effects of gamma irradiation on the longevity and fertility of
H. halys, to consider the possibility of using SIT to control BMSB. Previous work on
H. halys and another pentatomid species,
N. viridula, had already demonstrated the potential of SIT in managing alien pentatomid pests under favorable ecological conditions [
29,
30]. The use of the SIT with
H. halys in an area-wide IPM context was mainly considered for potential eradication or suppression programs in the case of early detection in New Zealand [
21,
29].
In the same way, the results presented in this work support the idea that
B. hilaris, a pentatomid with a narrow climatic range, could be suppressed or even eradicated in specific geographic areas and ecological conditions. The invasive Italian population of
B. hilaris confined on the small island of Pantelleria [
11] presents the opportunity to use the SIT in the future as a key component of an area-wide program in synergism with other least-toxic strategies.
In a traditional SIT program, the development of mass rearing facilities able to produce large numbers of sterile adult males that are sexually competitive with the wild-type is a prerequisite [
46,
48]. However, the mass-rearing process can also promote genetic drift, inducing genotypic differences between wild and laboratory populations [
49]. In other words, captivity can cause dramatic shifts in strain genetics, including reductions in diversity (heterozygosity), with a clear increase in the lines better adapted to a given experimental condition [
50,
51]. In the case of bagrada, the hypothesis of using wild populations is envisioned. As such, massive collections of wild bagrada bugs, especially during the fall and winter when the bug is aggregating in huge numbers around the senescent host plants (MC and RFHS, unpublished), are highly realistic. Field observations carried out with the invasive Italian population during the winter of 2020–2021 confirmed the aggregation behavior of
B. hilaris. Thus, the aggregation behavior and diapausing periods must be considered key periods for targeting bagrada control [
47,
52]. In some cases, these massive populations can be used as “small-scale mass-rearing natural biofactories” to provide and release wild-type sterile insects [
47,
49].
The results presented here warrant continued research to completely evaluate the performance of irradiated adults as we document how various irradiation doses impact the life history parameters of the bagrada bug. We began with low irradiation doses (16 Gy), increasing the dose according to physiological, biological, and behavioral responses, as previously described for BMSB [
29,
30]. A total of nine different doses (from 16 to 140 Gy) were evaluated on the newly emerged bagrada adults of both sexes. After BMSB [
29], this is the second example of measuring the impact of irradiation on stinkbugs.
Finally, mating in
B. hilaris is subordinated by female acceptance and mediated by the “correct” male courtship behavior [
53] and the production of a pheromone [
54]. Choice bioassays will be necessary to understand if irradiated males can compete with fertile males in terms of mating. Since the female has polyandric behavior [
55], additional screening must be carried out to evaluate the presence of cryptic sperm competition. In addition, feeding bioassays must be performed to provide a correct evaluation of the impact of the irradiated males, when released into the environment.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}


