
Effects of Adult Feeding Treatments on Longevity, Fecundity, Flight Ability, and Energy Metabolism Enzymes of Grapholita molesta Moths


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Feeding Regimes, Adult Longevity, and Fecundity
2.3. Feeding Regimes and Adult Flight Ability
2.4. Feeding Regimes and Activity of GAPDH and HOAD Enzymes
2.5. Statistical Analysis
3. Results
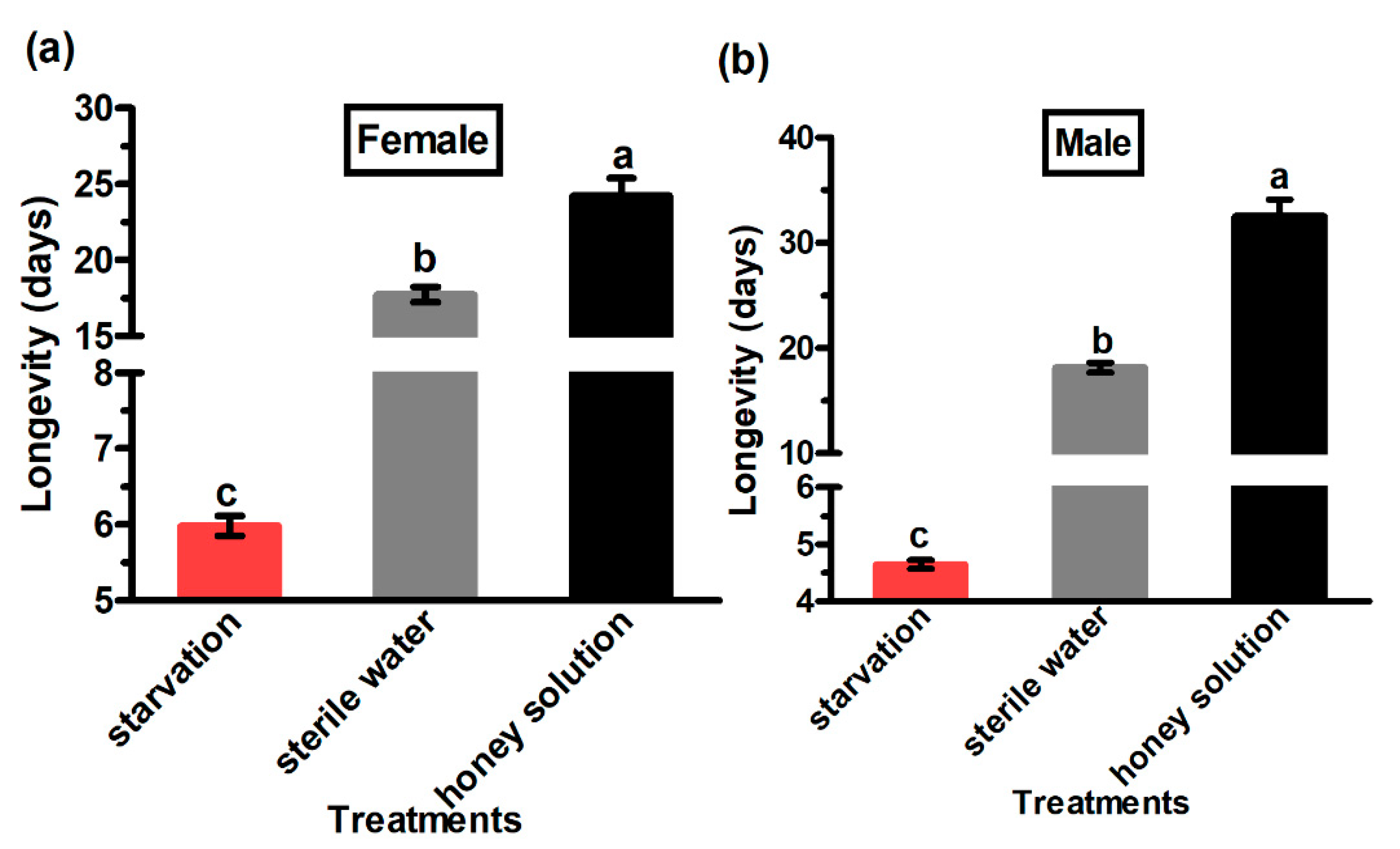
3.1. Adult Longevity
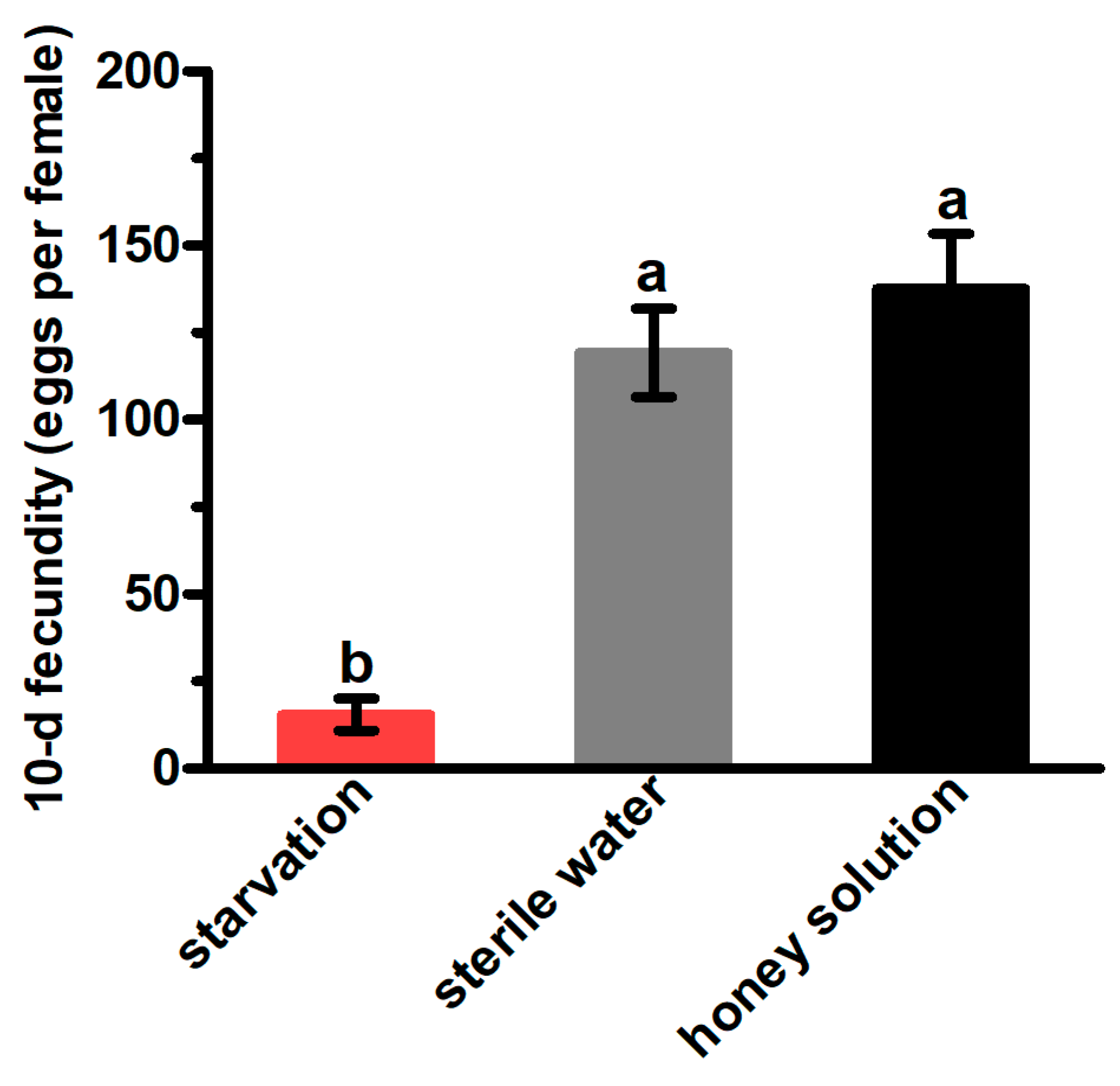
3.2. 10-d Fecundity
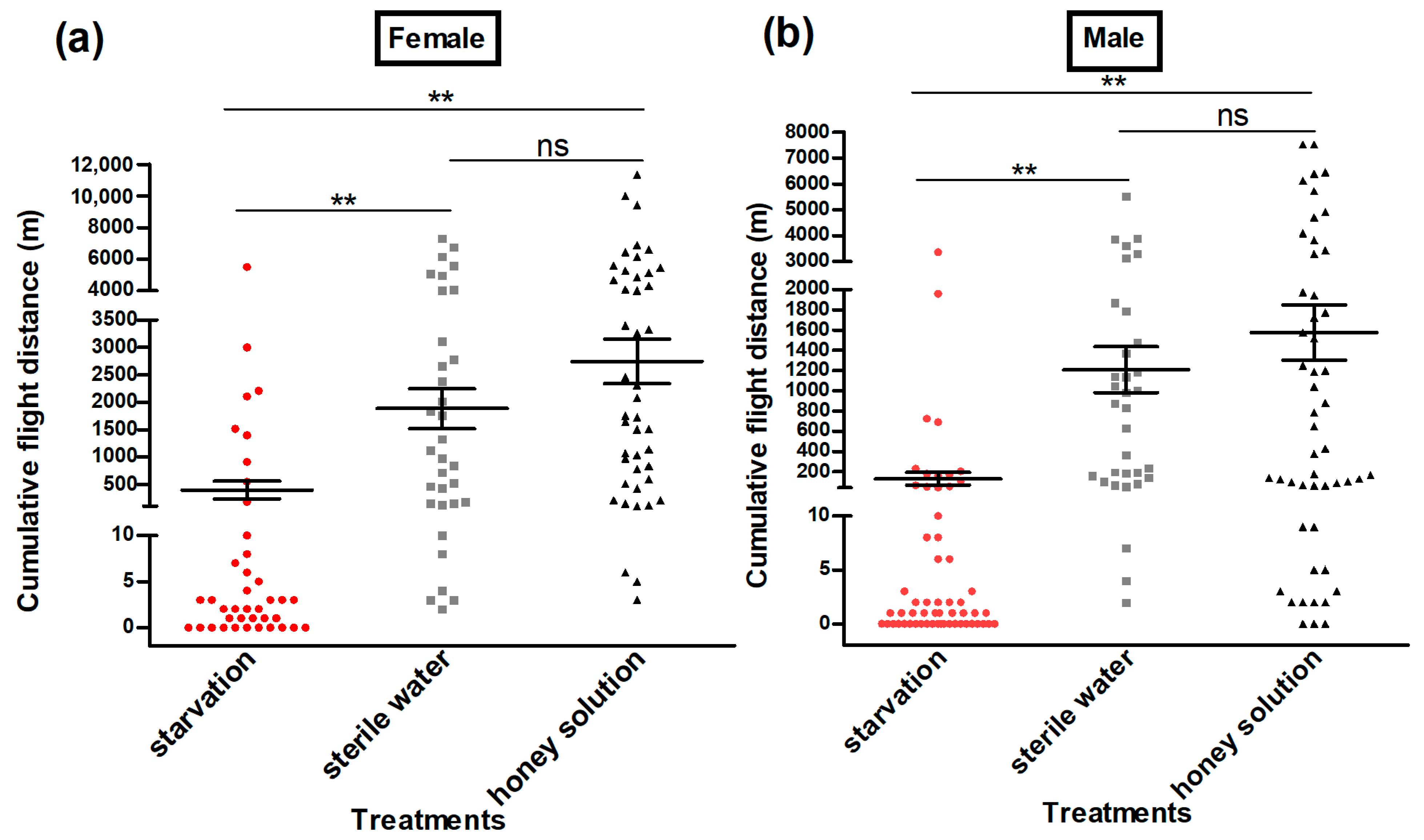
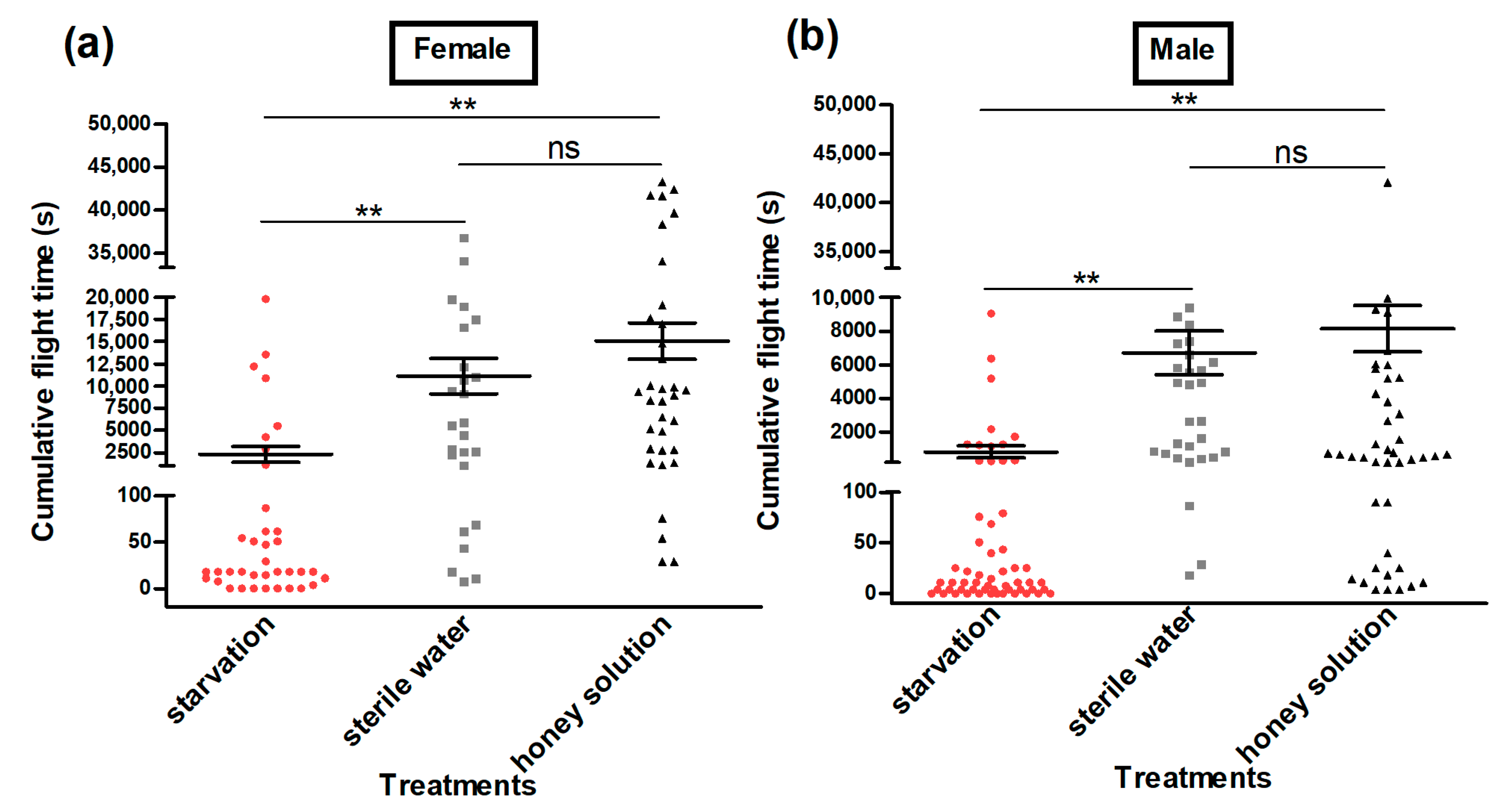
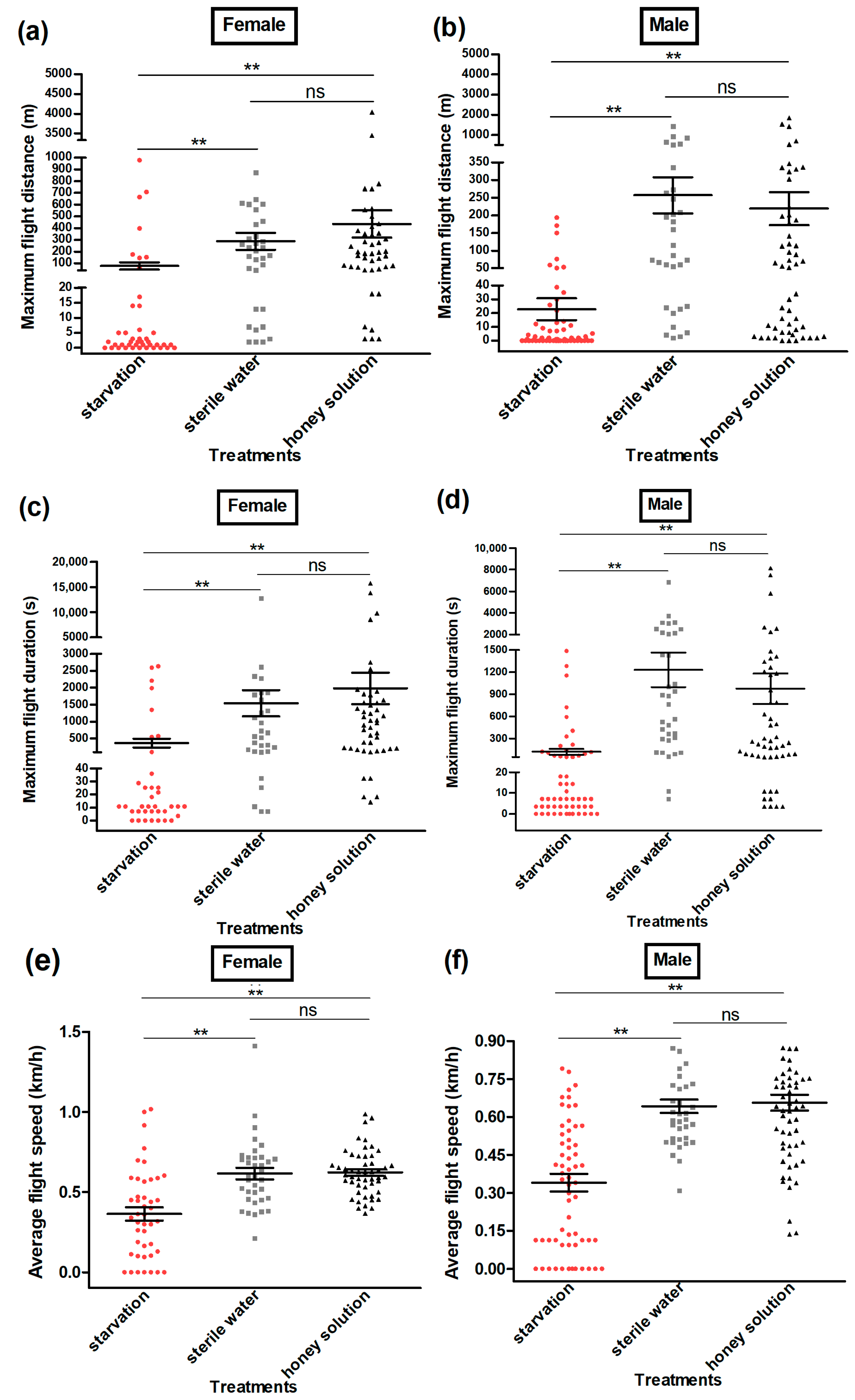
3.3. Flight Ability
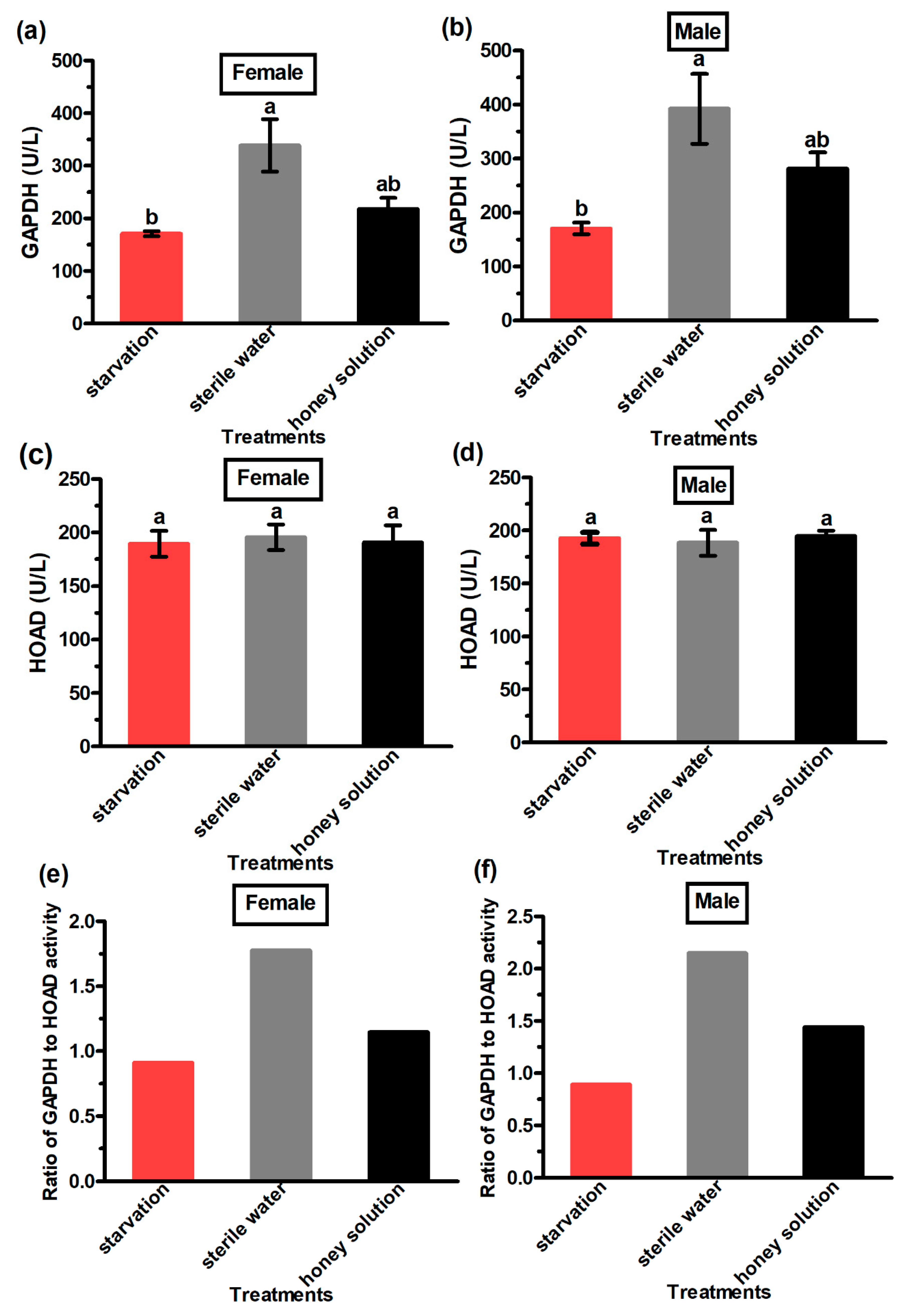
3.4. GAPDH and HOAD Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirk, H.; Dorn, S.; Mazzi, D. Worldwide population genetic structure of the oriental fruit moth (Grapholita molesta), a globally invasive pest. BMC Ecol. 2013, 13, 12. [Google Scholar] [CrossRef]
- Zheng, Y.; Peng, X.; Liu, G.M.; Pan, H.Y.; Dorn, S.; Chen, M.H. High genetic Diversity and structured populations of the oriental fruit moth in its range of origin. PLoS ONE 2013, 8, e78476. [Google Scholar] [CrossRef]
- Monteiro, L.B.; Witt, L.G.; Guiloski, I.C.; dos Santos, R.S.S.; de Assis, H.C.S. Evaluation of resistance management for the oriental fruit moth (Lepidoptera: Tortricidae) to insecticides in Brazilian apple orchards. J. Econ. Entomol. 2020, 3, 1411–1418. [Google Scholar] [CrossRef]
- Simmons, G.S.; Sepulveda, M.C.S.; Barrios, E.A.F.; Villegas, M.I.; Jimenez, R.E.M.; Jerez, A.R.G.; Henderson, R.; Riffo, H.D. Development of sterile insect technique for control of the European grapevine moth, Lobesia botrana, in urban areas of Chile. Insects 2021, 12, 378. [Google Scholar] [CrossRef] [PubMed]
- Marec, F.; Vreysen, M.J.B. Advances and challenges of using the sterile insect technique for the management of pest Lepidoptera. Insects 2019, 10, 371. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.N.; Wang, Y.; Jia, X.T.; Gao, Y.; Fan, R.J.; Li, J.; Ma, R.Y. Emergence and mating behavior of the oriental fruit moth Cydia molesta (lepidoptera: Tortricidae) and its potential for reproduction. Ann. Soc. Entomol. Fra. 2019, 55, 446–453. [Google Scholar] [CrossRef]
- Su, S.; Wang, X.T.; Jian, C.Z.; Ignatus, A.D.; Zhang, X.H.; Peng, X.; Chen, M.H. Life-history traits and flight capacity of Grapholita molesta (Lepidoptera: Tortricidae) using artificial diets with varying sugar content. J. Econ. Entomol. 2021, 114, 112–121. [Google Scholar] [CrossRef]
- Parker, A.G. Principles and practice in area-wide integrated pest management. Mass-rearing for sterile insect release. In Sterile Insect Technique; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 209–232. [Google Scholar]
- Calkins, C.O.; Parker, A.G. Principles and practice in area-wide integrated pest management. Sterile insect quality. In Sterile Insect Technique; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 269–296. [Google Scholar]
- Carpenter, J.E.; Blomefield, T.; Vreysen, M.J.B. A flight cylinder bioassay as a simple, effective quality control test for Cydia pomonella. J. Appl. Entomol. 2012, 136, 711–720. [Google Scholar] [CrossRef]
- Balestrino, F.; Puggioli, A.; Carrieri, M.; Bouyer, J.; Bellini, R. Quality control methods for Aedes albopictus sterile male production. PLoS Neglect. Trop. Diseases 2017, 11, e0005881. [Google Scholar] [CrossRef]
- Guo, J.W.; Li, P.; Zhang, J.; Liu, X.D.; Zhai, B.P.; Hu, G. Cnaphalocrocis medinalis moths decide to migrate when suffering nutrient shortage on the first day after emergence. Insects 2019, 10, 364. [Google Scholar] [CrossRef]
- Nation, J.L. Insect Physiology and Biochemistry, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 223–253. [Google Scholar]
- Wei, K.; Wang, X.Y.; Yang, Z.Q. Effects of supplementary nutrition on parasitism ability and developmental process of a gregarious parasitoid Sclerodermus pupariae (Hymenoptera: Bethylidae). For. Res. 2016, 29, 369–376. [Google Scholar]
- Heimpel, G.E.; Rosenheim, J.A.; Kattari, D. Adult feeding and lifetime reproductive success in the parasitoid Aphytis melinus. Entomol. Exp. Appl. 1997, 83, 305–315. [Google Scholar] [CrossRef]
- Harvey, J.A.; Cloutier, J.; Visser, B.; Ellers, J.; Wackers, F.L.; Gols, R. The effect of different dietary sugars and honey on longevity and fecundity in two hyperparasitoid wasps. J. Insect Physiol. 2012, 58, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Hari, N.S.; Jindal, J.; Malhi, N.S.; Khosa, J.K. Effect of adult nutrition and insect density on the performance of spotted stem borer, Chilo partellus in laboratory cultures. J. Pest Sci. 2008, 81, 23–27. [Google Scholar] [CrossRef]
- Collins, S.R.; Taylor, P.W. Flight ability procedures for mass-reared Queensland fruit flies, Bactrocera tryoni: An assessment of some variations. Entomol. Exp. Appl. 2010, 136, 308–311. [Google Scholar] [CrossRef]
- Bargielowski, I.; Kaufmann, C.; Alphey, L.; Reiter, P.; Koella, J. Flight performance and teneral energy reserves of two genetically-modified and one wild-type strain of the yellow fever mosquito Aedes aegypti. Vector Borne Zoonotic Dis. 2012, 12, 1053–1058. [Google Scholar] [CrossRef]
- Ge, S.S.; He, L.M.; He, W.; Xu, R.B.; Sun, X.T. Determination on moth flight capacity of Spodoptera frugiperda. Plant Prot. 2019, 45, 28–33. [Google Scholar]
- Shirai, Y. Flight activity, reproduction, and adult nutrition of the beet webworm, Spoladea recurvalis (Lepidoptera: Pyralidae). Appl. Entomol. Zool. 2006, 41, 405–414. [Google Scholar] [CrossRef]
- Beenakkers, A.M.T.; Van der Horst, D.J.; Van Marrewijk, W.J.A. Insect flight muscle metabolism. Insect Biochem. 1984, 14, 243–260. [Google Scholar] [CrossRef]
- Li, K.B.; Luo, L.Z. Activities of enzymes in the flight muscle of pupal and adult oriental armyworm, Mythimna separata (Walker). Acta Entomol. Sinica 1999, 42, 37–43. [Google Scholar]
- Beenakkers, A.M.T. Carbohydrate and fat as a fuel for insect flight: A comparative study. J. Insect Physiol. 1969, 15, 353–361. [Google Scholar] [CrossRef]
- He, C.; Meng, Q.K.; Hua, L.; Chen, W. Rearing Technique of oriental fruit moth (Lepodoptera: Tortricidae). J. Shanghai Jiaotong Univ. (Agri. Sci.) 2011, 29, 51–55. [Google Scholar]
- Yu, Q.; Feng, Y.T.; Guo, X.J.; Guo, G.M.; Hao, C.; Fan, R.J. Effects of humidity on the survival and fecundity of Grapholita molesta (Lepidoptera Tortricidae). Acta Entomol. Sinica 2017, 60, 659–665. [Google Scholar]
- Cheng, X.; Wang, J.Y.; Zhang, Q.; Jiang, R.D.; Feng, M.X. How to identify the female and male pupa and adult of the oriental fruit moth. Shanxi Fruits 2015, 6, 60–61. [Google Scholar]
- Hughes, J.; Dorn, S. Sexual differences in the flight performance of the oriental fruit moth. Entomol. Exp. Appl. 2002, 103, 171–182. [Google Scholar] [CrossRef]
- Su, S.; Jian, C.Z.; Zhang, X.H.; Fang, S.S.; Peng, X.; Piñero, J.C.; Chen, M.H. Sublethal effects of abamectin on the development, reproduction, detoxification enzyme activity, and related gene expression of the oriental fruit moth (Lepidoptera: Tortricidae). J. Econ. Entomol. 2021, 114, 2430–2438. [Google Scholar] [CrossRef] [PubMed]
- Forte, S.N.; Ferrrro, A.A.; Alonso, T.S. Content and composition of phosphoglycerols and neutral lipids at different developmental stages of the eggs of the codling moth, Cydia pomonella (Lepidoptera: Tortricidae). Arch. Insect Biochem. Physiol. 2002, 50, 121–130. [Google Scholar] [CrossRef]
- Kim, K.N.; Sin, U.C.; Yun, C.N.; Song, H.S.; Huang, Z.J.; Huang, Q.Y.; Lei, C.L. Influence of green light illumination on several enzymes involved in energy metabolism in the oriental armyworm, Mythimna separata (Lepidoptera: Noctuidae). J. Asia-Pac. Entomol. 2019, 22, 487–492. [Google Scholar] [CrossRef]
- Stevens, P.; Froud, K.; Jamieson, L. Effects of adult feeding on longevity and fecundity of Ctenopseustis obliquana (Lepidoptera: Tortricidae). N. Z. J. Crop Hort. Sci. 2002, 30, 229–234. [Google Scholar] [CrossRef]
- Wang, W.J.; Chen, E.Z.; Li, H.Z. ZIR: Modified Wilcoxon Rank Test and Kruskal Wallis Test for Zero-Inflated Data, ZIR (Zero-Inflated Rank Test) R Package Version 1.0.0. 2021. Available online: https://github.com/chvlyl/ZIR (accessed on 5 August 2022).
- Siekmann, G.; Tenhumberg, B.; Keller, M.A. Feeding and survival in parasitic wasps: Sugar concentration and timing matter. OIKOS 2001, 95, 425–430. [Google Scholar] [CrossRef]
- Eijs, I.E.M.; Ellers, J.; Van Duinen, G.J. Feeding strategies in drosophilid parasitoids: The impact of natural food resources on energy reserves in females. Ecol. Entomol. 1998, 23, 133–138. [Google Scholar] [CrossRef]
- Snart, C.J.P.; Kapranas, A.; Williams, H.; Barrett, D.A.; Hardy, I.C.W. Sustenance and performance: Nutritional reserves, longevity, and contest outcomes of fed and starved adult parasitoid wasps. Front. Ecol. Evol. 2018, 6, 12. [Google Scholar] [CrossRef]
- Savopoulou-Soultani, M.; Milonas, P.G.; Stavridis, D.G. Role of availability of food to the adult Lobesia botrana (Lepidoptera: Tortricidae) in its reproductive performance. J. Econ. Entomol. 1998, 91, 1341–1348. [Google Scholar] [CrossRef]
- Gunn, A.; Gatehouse, A.G. Effects of the availability of food and water on reproduction in the African army worm, Spodoptera exempta. Physiol. Entomol. 2010, 10, 53–63. [Google Scholar] [CrossRef]
- Klassen, W. Area-wide integrated pest management and the sterile insect technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 39–69. [Google Scholar]
- Sciarretta, A.; Trematerra, P. Geostatistical characterization of the spatial distribution of Grapholita molesta and Anarsia lineatella males in agricultural landscape. J. Appl. Entomol. 2006, 130, 73–83. [Google Scholar] [CrossRef]
- Wang, X.G.; Johnson, M.W.; Daane, K.N.; Opp, S. Combined effects of heat stress and food supply on flight performance of olive fruit fly (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2009, 102, 727–734. [Google Scholar] [CrossRef]
- Weber, D.C.; Ferro, D.N. Flight and fecundity of Colorado potato beetles (Coleoptera: Chrysomelidae) fed on different diets. Ann. Entomol. Soc. Am. 1996, 89, 297–306. [Google Scholar] [CrossRef]
- Han, L.Z.; Zhai, B.P.; Zhang, X.X.; Liu, P.L. Activity of enzymes related to energy metabolism in the flight muscle of beet armyworm. Acta Ecol. Sinica 2005, 25, 1101–1106. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, S.; Zhang, X.; Jian, C.; Huang, B.; Peng, X.; Vreysen, M.J.B.; Chen, M. Effects of Adult Feeding Treatments on Longevity, Fecundity, Flight Ability, and Energy Metabolism Enzymes of Grapholita molesta Moths. Insects 2022, 13, 725. https://doi.org/10.3390/insects13080725
Su S, Zhang X, Jian C, Huang B, Peng X, Vreysen MJB, Chen M. Effects of Adult Feeding Treatments on Longevity, Fecundity, Flight Ability, and Energy Metabolism Enzymes of Grapholita molesta Moths. Insects. 2022; 13(8):725. https://doi.org/10.3390/insects13080725
Chicago/Turabian StyleSu, Sha, Xiaohe Zhang, Chengzhi Jian, Baojian Huang, Xiong Peng, Marc J. B. Vreysen, and Maohua Chen. 2022. "Effects of Adult Feeding Treatments on Longevity, Fecundity, Flight Ability, and Energy Metabolism Enzymes of Grapholita molesta Moths" Insects 13, no. 8: 725. https://doi.org/10.3390/insects13080725