
Life Table Parameters and Digestive Enzyme Activity of Spodoptera littoralis (Boisd) (Lepidoptera: Noctuidae) on Selected Legume Cultivars



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Legume Cultivars
2.2. Insect Rearing
2.3. Life History Parameters and Body Mass in Immature Stages of S. littoralis on Legume Cultivars
2.4. Life Table Variables of Adult S. littoralis Developing on Various Legume Cultivars
2.5. Life Table Analysis
2.6. Preparation of Midgut Extracts
2.7. Determination of Proteolytic Activity of S. littoralis after Feeding on Various Legume Cultivars
2.8. Determination of Amylolytic Activity of S. littoralis on Legume Cultivars
2.9. Biochemical Characteristics of Legume Cultivars
2.10. Statistical and Cluster Analysis
3. Results
3.1. Life History Parameters and Body Weight
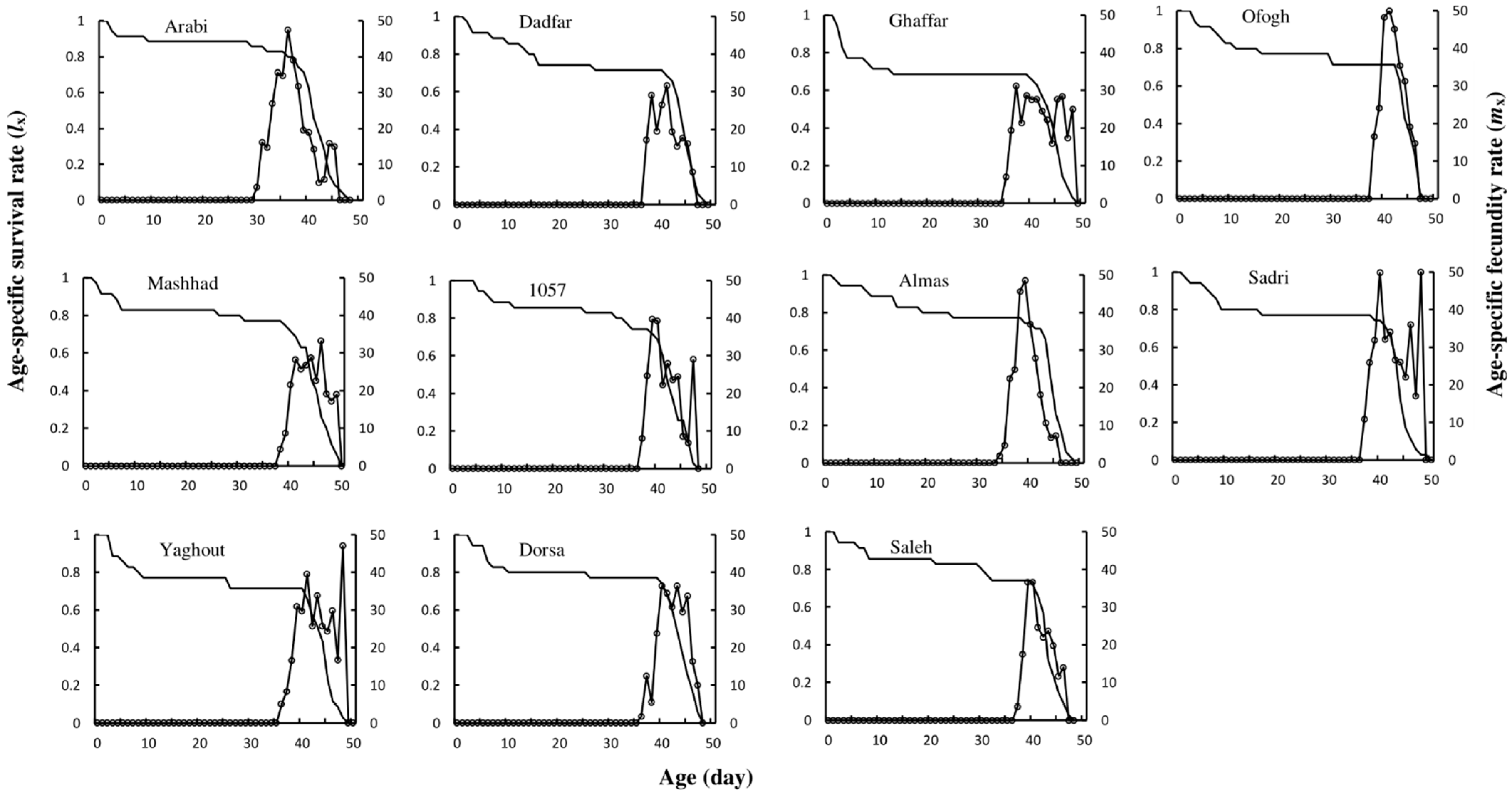
3.2. Adult Life Table Variables
3.3. The Activity of Digestive Enzymes
3.4. Cluster Analysis
3.5. Biochemical Characteristics of Various Legume Cultivars
3.6. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sneh, B.; Schuster, S.; Broza, M. Insecticidal activity of Bacillus thuringiensis strains against the Egyptian cotton leafworm Spodoptera littoralis (Lep.: Noctuidae). Entomophaga 1981, 26, 179–190. [Google Scholar] [CrossRef]
- Azab, S.G.; Sadek, M.M.; Crailsheim, K. Protein metabolism in larvae of the cotton leaf-worm Spodoptera littoralis (Lepidoptera: Noctuidae) and its response to three mycotoxins. Environ. Entomol. 2001, 30, 817–823. [Google Scholar] [CrossRef]
- Zamani Fard, S.; Hemmati, S.A.; Shishehbor, P.; Stelinski, L.L. Growth, consumption and digestive enzyme activities of Spodoptera littoralis (Boisd) on various mung bean cultivars reveal potential tolerance traits. J. Appl. Entomol. 2022, 146, 1–10. [Google Scholar] [CrossRef]
- Shishehbor, P.; Hemmati, S.A. Investigating of secondary metabolites in bean cultivars and their impact on nutritional performance of Spodoptera littoralis (Lep.: Noctuidae). Bull. Entomol. Res. 2022, 112, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Jimenez, A.; Arboleda, J.W.; Lopez Avila, A.; Grossi-de-Sa, M.F. Digestive α-amylases from Tecia solanivora larvae (Lepidoptera: Gelechiidae): Response to pH, temperature and plant amylase inhibitors. Bull. Entomol. Res. 2008, 98, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Smagghe, G.; Carton, B.; Wesemael, W.; Ishaaya, I.; Tirry, L. Ecdysone agonists-mechanism of action and application on Spodoptera species. Pestic. Sci. 1999, 55, 343–389. [Google Scholar] [CrossRef]
- Ongley, E.D. Pesticides as water pollutants. In Control of Water Pollution from Agriculture, Food and Agriculture Organization of the United Nations; United Nations: New York, NY, USA, 1996; pp. 53–66. [Google Scholar]
- Korrat, E.E.E.; Abdelmonem, A.E.; Helalia, A.A.R.; Khalifa, H.M.S. Toxicological study of some conventional and nonconventional insecticides and their mixtures against cotton leaf worm, Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae). Annal. Agric. Sci. 2012, 57, 145–152. [Google Scholar] [CrossRef]
- Panda, N.; Khush, G.S. Host Plant Resistance to Insects; CAB International: Oxon, UK, 1995. [Google Scholar]
- La Rossa, F.R.; Vasicek, A.; Lopez, M.C. Effects of pepper (Capsicum annuum) cultivars on the biology and life table parameters of Myzus persicae (Sulz.) (Hemiptera: Aphididae). Neotrop. Entomol. 2013, 42, 634–641. [Google Scholar] [CrossRef]
- Price, P.W.; Bouton, C.E.; Gross, P.; McPheron, B.A.; Thompson, J.N.; Weis, A.E. Interactions among three trophic levels: Influence of plants on interactions between insect herbivores and natural enemies. Ann. Rev. Ecol. Syst. 1980, 11, 41–65. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.M.; Clement, S.L. Molecular bases of plant resistance to arthropods. Ann. Rev. Entomol. 2012, 57, 309–328. [Google Scholar] [CrossRef]
- Karimi-Malati, A.; Fathipour, Y.; Talebi, A.A.; Bazoubandi, M. Life table parameters and survivorship of Spodoptera exigua (Lepidoptera: Noctuidae) at constant temperatures. Environ. Entomol. 2014, 43, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, J.; Gawlak, M.; Szafranek, J.; Szafranek, B.; Synak, E.; Warchalewski, J.R.; Piasecka-Kwiatkowska, D.; Błaszczak, W.; Jeliński, T.; Fornal, J. The effect of wheat grain composition, cuticular lipids and kernel surface microstructure on feeding, egg-laying, and the development of the granary weevil, Sitophilus granarius (L.). J. Stored. Prod. Res. 2010, 46, 133–141. [Google Scholar] [CrossRef]
- Razmjou, J.; Naseri, B.; Hemati, S.A. Comparative performance of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) on various host plants. J. Pest. Sci. 2014, 87, 29–37. [Google Scholar] [CrossRef]
- Naseri, B.; Hamzavi, F.; Ebadollahi, A.; Sheikh, F. Physicochemical traits of Vicia faba L. seed cultivars affect oviposition preference and demographic parameters of Callosobruchus maculatus (F.) (Coleoptera: Chrysomelidae). J. Stored. Prod. Res. 2022, 95, 101924. [Google Scholar] [CrossRef]
- Nouri-Ganbalani, G.; Borzoui, E.; Shahnavazi, M.; Nouri, A. Induction of resistance against Plutella xylostella (L.) (Lep.: Plutellidae) by jasmonic acid and mealy cabbage aphid feeding in Brassica napus L. Front. Physiol. 2018, 9, 859. [Google Scholar] [CrossRef] [PubMed]
- Abedi, Z.; Golizadeh, A.; Soufbaf, M.; Hassanpour, M.; Jafari-Nodoushan, A.; Akhavan, H.R. Effect of different fruit developmental stages and biochemical metabolites of pomegranate (Ponicaceae) on life history parameters of Ectomyelois ceratoniae (Lepidoptera: Pyralidae). J. Econ. Entomol. 2020, 113, 964–973. [Google Scholar] [CrossRef]
- Shirinbeik Mohajer, S.; Golizadeh, A.; Hassanpour, M.; Fathi, S.A.A.; Sedaratian-Jahromi, A.; Abedi, Z. Interaction between biological parameters of Panonychus citri (Acari: Tetranychidae) and some phytochemical metabolites in different citrus species. Bull. Entomol. Res. 2022, 112, 509–519. [Google Scholar] [CrossRef]
- Heidari, N.; Sedaratian-Jahromi, A.; Ghane-Jahromi, M.; Zalucki, M.P. How bottom-up effects of different tomato cultivars affect population responses of Tuta absoluta (Lep.: Gelechiidae): A case study on host plant resistance. Arthropod Plant Interact. 2020, 14, 181–192. [Google Scholar] [CrossRef]
- Hemati, S.A.; Naseri, B.; Ganbalani, G.N.; Dastjerdi, H.R.; Golizadeh, A. Digestive proteolytic and amylolytic activities and feeding responses of Helicoverpa armigera (Noctuidae: Lepidoptera) on different host plants. J. Econ. Entomol. 2012, 105, 1439–1446. [Google Scholar] [CrossRef]
- Hemmati, S.A.; Takalloo, Z.; Taghdir, M.; Mehrabadi, M.; Balalaei, S.; Moharramipour, S.; Sajedi, R.H. The trypsin inhibitor pro-peptide induces toxic effects in Indianmeal moth, Plodia interpunctella. Pestic. Biochem. Phys. 2012, 171, 104730. [Google Scholar] [CrossRef]
- Babamir-Satehi, A.; Habibpour, B.; Aghdam, H.R.; Hemmati, S.A. Interaction between feeding efficiency and digestive physiology of the pink stem borer, Sesamia cretica Lederer (Lepidoptera: Noctuidae), and biochemical compounds of different sugarcane cultivars. Arthropod Plant Interact. 2022, 16, 1–8. [Google Scholar] [CrossRef]
- Patanka, A.G.; Giri, A.P.; Harsulkar, A.M.; Sainani, M.N.; Deshpande, V.V.; Ranjekar, P.K.; Gupta, V.S. Complexity in specificities and expression of Helicoverpa armigera gut proteases explains polyphagous nature of the insect pest. Insect Biochem. Mol. Biol. 2001, 31, 453–464. [Google Scholar] [CrossRef]
- Pereira, P.J.B.; Lozanov, V.; Patthy, A.; Huber, R.; Bode, W.; Pongor, S.; Strobl, S. Specific inhibition of insect α-amylases: Yellow mealworm α-amylase in complex with the Amaranth α-amylase inhibitor at 2.0 Å resolution. Structure 1999, 9, 1079–1088. [Google Scholar] [CrossRef] [Green Version]
- Hegazi, E.M.; Schopf, R. The influence of temperature on consumption and utilization of artificial diet by Spodoptera littoralis (Boisd.) (Lepidopt., Noctuidae). Z. Angew. Entomol. 1984, 97, 321–326. [Google Scholar] [CrossRef]
- Khedr, M.A.; AL-Shannaf, H.M.; Mead, H.M.; Shaker, S.A. Comparative study to determine food consumption of cotton leafworm, Spodoptera littoralis, on some cotton genotypes. J. Plant. Protec. Res. 2015, 55, 312–321. [Google Scholar] [CrossRef]
- Gacemi, A.; Taibi, A.; Abed, N.E.H.; M’hammedi Bouzina, M.; Bellague, D.; Tarmoul, K. Effect of four host plants on nutritional performance of cotton leafworm, Spodoptera littoralis (Lepidoptera: Noctuidae). J. Crop. Protec. 2019, 8, 361–371. [Google Scholar]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Huang, Y.B.; Chi, H. Age-stage, two-sex life tables of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect. Sci. 2012, 19, 263–273. [Google Scholar] [CrossRef]
- Elpidina, E.N.; Vinokurov, K.S.; Gromenko, V.A.; Rudenshaya, Y.A.; Dunaevsky, Y.E.; Zhuzhikov, D.P. Compartmentalization of proteinases and amylases in Nauphoeta cinerea midgut. Arch. Insect. Biochem. Physiol. 2001, 48, 206–216. [Google Scholar] [CrossRef]
- Bradford, M.A. Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bernfeld, P. Amylase, α and β. Methods Enzymol. 1955, 1, 149–158. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. Available online: http://140.120.197.173/Ecology/prod02.htm (accessed on 1 August 2018).
- SPSS Inc. SPSS Base 16.0 User’s Guide; SPSS Incorporation: Chicago, IL, USA, 2007. [Google Scholar]
- Magaña, A.J.; Dáder, B.; Sancho, G.; Adán, Á.; Morales, I.; Viñuela, E. Comparison of the Parasitization of Chelonus inanitus L. (Hymenoptera: Braconidae) in Two Spodoptera Pests and Evaluation of the Procedure for Its Production. Insects 2022, 13, 99. [Google Scholar] [CrossRef] [PubMed]
- Golizadeh, A.; Abedi, Z. Comparative performance of the Khapra beetle, Trogoderma granarium Everts (Coleoptera: Dermestidae) on various wheat cultivars. J. Stored. Prod. Res. 2016, 69, 159–165. [Google Scholar] [CrossRef]
- Chen, Y.; Ni, X.; Buntin, G.D. Physiological, nutritional, and biochemical bases of corn resistance to foliage-feeding fall armyworm. J. Chem. Ecol. 2009, 35, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Soufbaf, M.; Fathipour, Y.; Karimzadeh, J.; Zalucki, M.P. Bottom-up effect of different host plants on Plutella xylostella (Lepidoptera: Plutellidae): A life-table study on canola. J. Econ. Entomol. 2010, 103, 2019–2027. [Google Scholar] [CrossRef] [PubMed]
- Carey, J.R. Applied Demography for Biologists with Special Emphasis on Insects; Oxford University Press, Inc.: New York, NY, USA, 1993. [Google Scholar]
- Jongsma, M.A.; Bolter, C.J. The adaptation of insects to plant protease inhibitors. J. Insect Physiol. 1997, 43, 885–895. [Google Scholar] [CrossRef]
- Broadway, R.M. Dietary regulation of serine proteinases that are resistant to serine proteinase inhibitors. J. Insect Physiol. 1997, 43, 855–874. [Google Scholar] [CrossRef]
- Girard, C.; Le Metayer, M.; Bonade-Bottino, M.; Pham-Delegue, M.H.; Jouanin, L. High level of resistance to proteinase inhibitors may be conferred by proteolytic cleavage in beetle larvae. Insect Biochem. Mol. Biol. 1998, 28, 229–237. [Google Scholar] [CrossRef]
- Blanco-Labra, A.; Chagolla-Lopez, A.; Martinez-Gallardo, N.; Valdes-Rodriguez, S. Further characterization of the 12 kDa protease/alpha amylase inhibitor present in maize seeds. J. Food Biochem. 1995, 19, 27–41. [Google Scholar] [CrossRef]
- Franco, O.L.; Rigden, D.J.; Melo, F.R.; Grossi-de-Sa, M.F. Plant α-amylase inhibitors and their interaction with insect α-amylases: Structure, function and potential for crop protection. FEBS J. 2002, 268, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, C.; Garcia-Casado, G.; Sanchez-Monge, R.; Gomez, L.; Castenera, P.; Salcedo, G. Three inhibitor types from wheat endosperm are differentially active against α-amylases of lepidoptera pests. Entomol. Exp. Appl. 1993, 66, 47–52. [Google Scholar] [CrossRef]
- Giri, A.P.; Kachole, M.V. Amylase inhibitors of pigeon pea (Cajanus cajan) seeds. Phytochemistry 1998, 47, 197–202. [Google Scholar] [CrossRef]
- Dias, S.C.; Franco, O.L.; Magalhaes, C.P.; de Oliveira-Neto, O.B.; Laumann, R.A.; Figueira, E.L.Z.; Melo, F.R.; Grossi-de-Sa, M.F. Molecular cloning and expression of an α-amylase inhibitor from rye with potential for controlling insect pests. Protein J. 2005, 24, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Franco, O.L.; Rigden, D.J.; Melo, F.R.; Bloch, C., Jr.; Silva, C.P.; Grossi-de-Sa, M.F. Activity of wheat α-amylase inhibitors towards bruchid α-amylases and structural explanation of observed specificities. FEBS J. 2000, 267, 2166–2173. [Google Scholar] [CrossRef]
- Harsulkar, A.M.; Giri, A.P.; Patankar, A.G.; Gupta, V.S.; Sainani, M.N.; Ranjekar, P.K.; Deshpande, V.V. Successive use of non-host plant proteinase inhibitors required for effective inhibition of gut proteinases and larval growth of Helicoverpa armigera. Plant Physiol. 1999, 121, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Sivakumar, S.; Mohan, M.; Franco, O.L.; Thayumanavan, B. Inhibition of insect pest α-amylases by little and finger millet inhibitors. Pestic. Biochem. Physiol. 2006, 85, 155–160. [Google Scholar] [CrossRef]
- Lazarevic, J.; Peric-Mataruga, V.; Vlahovic, M.; Mrdakovic, M. Effects of rearing density on larval growth and activity of digestive enzymes in Lymantria dispar L. (Lepidoptera: Lymantriidae). Folia Biol. 2004, 52, 105–112. [Google Scholar]
- Greenberg, S.M.; Sappington, T.W.; Legaspi, B.C.; Liu, T.X.; Setamou, M. Feeding and life history of Spodoptera exigua (Lepidoptera: Noctuidae) on different host plants. Ann. Entomol. Soc. Am. 2001, 94, 566–575. [Google Scholar] [CrossRef]
- Saeed, S.; Sayyed, A.H.; Ahmad, I. Effect of host plants on life history traits of Spodoptera exigua (Lepidoptera: Noctuidae). J. Pest Sci. 2009, 83, 165–172. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Legume Cultivar | Egg Incubation | Larval Period | Pre-Pupal Period | Pupal Period | Development Time |
|---|---|---|---|---|---|
| Arabi | 3.937 ± 0.043 c | 17.387 ± 0.305 d | 1.515 ± 0.102 c | 9.793 ± 0.157 e | 32.515 ± 0.393 d |
| Dadfar | 4.095 ± 0.051 b | 20.770 ± 0.157 b | 1.960 ± 0.040 a | 10.240 ± 0.103 d | 37.041 ± 0.251 bc |
| Ghaffar | 4.221 ± 0.079 ab | 20.043 ± 0.200 c | 1.542 ± 0.103 bc | 10.293 ± 0.140 cd | 36.086 ± 0.336 c |
| Ofogh | 4.218 ± 0.072 ab | 21.592 ± 0.255 a | 1.999 ± 0.017 a | 10.840 ± 0.167 a | 38.639 ± 0.317 a |
| Mashhad | 4.188 ± 0.69 ab | 21.758 ± 0.404 a | 2.283 ± 0.280 a | 10.41 ± 0.347 abcd | 38.740 ± 0.473 a |
| 1057 | 4.258 ± 0.073 ab | 20.770 ± 0.200 b | 1.787 ± 0.077 ab | 10.384 ± 0.144 bcd | 37.083 ± 0.251 b |
| Almas | 4.121 ± 0.057 b | 19.712 ± 0.142 c | 2.001 ± 0.074 a | 9.888 ± 0.080 e | 35.702 ± 0.180 c |
| Sadri | 4.364 ± 0.082 a | 20.852 ± 0.187 b | 1.814 ± 0.075 a | 10.630 ± 0.157 abc | 37.668 ± 0.250 b |
| Yaghout | 4.129 ± 0.060 b | 21.006 ± 0.223 ab | 1.960 ± 0.040 a | 10.763 ± 0.154 ab | 37.849 ± 0.359 ab |
| Dorsa | 4.212 ± 0.071 ab | 20.786 ± 0.180 b | 1.926 ± 0.050 a | 10.555 ± 0.133 abcd | 37.482 ± 0.238 b |
| Saleh | 4.243 ± 0.075 ab | 20.655 ± 0.114 b | 2.034 ± 0.035 a | 10.230 ± 0.099 d | 37.115 ± 0.168 b |
| Legume Cultivar | APOP (Day) | TPOP (Day) | Oviposition Period (Day) | Fecundity (Offspring) | Female Adult Longevity (Day) | Male Adult Longevity (Day) |
|---|---|---|---|---|---|---|
| Arabi | 1.053 ± 0.052 d | 33.470 ± 0.443 e | 6.849 ± 0.457 ab | 445.988 ± 25.16 a | 9.851 ± 0.527 ab | 9.303 ± 0.445 a |
| Dadfar | 1.073 ± 0.072 cd | 38.643 ± 0.370 ab | 5.853 ± 0.354 bc | 308.520 ± 25.412 b | 8.430 ± 0.364 cd | 7.458 ± 0.433 b |
| Ghaffar | 1.469 ± 0.161 ab | 37.203 ± 0.425 cd | 7.138 ± 0.288 a | 363.198 ± 7.046 ab | 9.001 ± 0.215 bc | 8.216 ± 0.494 ab |
| Ofogh | 1.331 ± 0.125 abc | 39.398 ± 0.267 a | 5.401 ± 0.232 c | 422.150 ± 12.08 a | 7.070 ± 0.366 f | 6.097 ± 0.402 cd |
| Mashhad | 1.643 ± 0.131 a | 39.501 ± 0.460 a | 5.92 ± 0.447 bc | 289.83 ± 11.51 b | 7.351 ± 0.505 def | 5.462 ± 0.285 cd |
| 1057 | 1.214 ± 0.113 bcd | 38.145 ± 0.249 bc | 5.282 ± 0.467 c | 311.258 ± 15.90 b | 7.137 ± 0.494 ef | 6.079 ± 0.492 cd |
| Almas | 1.251 ± 0.143 bcd | 36.871 ± 0.281 d | 6.382 ± 0.451 abc | 406.011 ± 34.40 ab | 10.001 ± 0.396 a | 7.991 ± 0.553 ab |
| Sadri | 1.236 ± 0.104 bcd | 38.703 ± 0.312 ab | 5.825 ± 0.280 bc | 345.396 ± 9.11 ab | 7.532 ± 0.325 def | 5.099 ± 0.375 d |
| Yaghout | 1.307 ± 0.130 abcd | 38.701 ± 0.441 ab | 5.92 ± 0.135 bc | 391.449 ± 10.77 ab | 8.001 ± 0.195 de | 5.582 ± 0.304 cd |
| Dorsa | 1.425 ± 0.172 abc | 38.429 ± 0.355 ab | 5.857 ± 0.367 bc | 357.850 ± 11.19 ab | 7.569 ± 0.443 def | 6.157 ± 0.316 c |
| Saleh | 1.077 ± 0.076 cd | 38.232 ± 0.255 b | 5.925 ± 0.304 bc | 303.915 ± 20.378 b | 7.847 ± 0.457 def | 5.308 ± 0.280 d |
| Legume Cultivar | R0 (Offspring per Adult) | r (Day−1) | (Day−1) | T (Day) |
|---|---|---|---|---|
| Arabi | 242.34 ± 24.20 a | 0.1509 ± 0.006 a | 1.1628 ± 0.007 a | 36.27 ± 0.54 e |
| Dadfar | 123.59 ± 17.21 b | 0.1163 ± 0.007 bc | 1.1234 ± 0.008 bc | 41.09 ± 0.42 bc |
| Ghaffar | 155.58 ± 15.07 ab | 0.1254 ± 0.006 bc | 1.1337 ±0.006 bc | 40.07 ± 0.51 d |
| Ofogh | 180.64 ± 17.81 ab | 0.1237 ± 0.005 bc | 1.1316 ± 0.006 bc | 41.85 ± 0.31 ab |
| Mashhad | 118.78 ± 13.42 b | 0.1115 ± 0.006 c | 1.1180 ± 0.006 c | 42.60 ± 0.43 a |
| 1057 | 124.48 ± 28.93 b | 0.1176 ± 0.006 bc | 1.1248 ± 0.007 bc | 40.78 ± 0.41 d |
| Almas | 184.89 ± 22.84 ab | 0.1304 ± 0.007 b | 1.1393 ± 0.007 b | 39.79 ± 0.35 d |
| Sadri | 168.02 ± 15.42 ab | 0.1241 ± 0.005 bc | 1.1322 ± 0.005 bc | 41.15 ± 0.37 bc |
| Yaghout | 144.76 ± 16.95 ab | 0.1195 ± 0.006 bc | 1.1269 ± 0.007 bc | 41.42 ± 0.48 bc |
| Dorsa | 143.04 ± 15.46 ab | 0.1193 ± 0.005 bc | 1.1267 ± 0.006 bc | 41.40 ± 0.36 bc |
| Saleh | 112.58 ± 12.19 b | 0.1140 ± 0.007 bc | 1.1207 ± 0.008 bc | 41.13 ± 0.29 bc |
| Legume Cultivar | Protein Content | Starch Content |
|---|---|---|
| Arabi | 0.315 ± 0.018 abc | 2.569 ± 0.028 a |
| Dadfar | 0.223 ± 0.006 f | 2.245 ± 0.049 cd |
| Ghaffar | 0.295 ± 0.009 bcd | 2.268 ± 0.010 bcd |
| Ofogh | 0.239 ± 0.005 ef | 2.247 ± 0.055 cd |
| Mashhad | 0.304 ± 0.020 abcd | 2.312 ± 0.077 abcd |
| 1057 | 0.314 ± 0.003 abc | 2.426 ± 0.094 abc |
| Almas | 0.343 ± 0.010 a | 2.441 ± 0.021 abc |
| Sadri | 0.283 ± 0.005 cd | 2.257 ± 0.044 bcd |
| Yaghout | 0.225 ± 0.005 ef | 2.130 ± 0.037 d |
| Dorsa | 0.334 ± 0.005 ab | 2.536 ± 0.049 ab |
| Saleh | 0.267 ± 0.005 de | 2.303 ± 0.087 abcd |
| Parameter | Protein Content | Starch Content |
|---|---|---|
| Pupal weight | −0.059 (0.742) | −0.129 (0.476) |
| Development time | −0.386 (0.026) | −0.325 (0.065) |
| Fecundity | 0.082 (0.650) | 0.318 (0.071) |
| R0 | −0.026 (0.888) | −0.011 (0.954) |
| r | 0.321 (0.068) | 0.609 (0.047) |
| Amylolytic activity | 0.277 (0.119) | 0.619 (0.042) |
| Proteolytic activity | −0.041 (0.819) | −0.007 (0.968) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hemmati, S.A.; Shishehbor, P.; Stelinski, L.L. Life Table Parameters and Digestive Enzyme Activity of Spodoptera littoralis (Boisd) (Lepidoptera: Noctuidae) on Selected Legume Cultivars. Insects 2022, 13, 661. https://doi.org/10.3390/insects13070661
Hemmati SA, Shishehbor P, Stelinski LL. Life Table Parameters and Digestive Enzyme Activity of Spodoptera littoralis (Boisd) (Lepidoptera: Noctuidae) on Selected Legume Cultivars. Insects. 2022; 13(7):661. https://doi.org/10.3390/insects13070661
Chicago/Turabian StyleHemmati, Seyed Ali, Parviz Shishehbor, and Lukasz L. Stelinski. 2022. "Life Table Parameters and Digestive Enzyme Activity of Spodoptera littoralis (Boisd) (Lepidoptera: Noctuidae) on Selected Legume Cultivars" Insects 13, no. 7: 661. https://doi.org/10.3390/insects13070661