
Optimizing the Use of Basil as a Functional Plant for the Biological Control of Aphids by Chrysopa pallens (Neuroptera: Chrysopidae) in Greenhouses


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Plants
2.2. Laboratory Trials
2.3. Greenhouse Assays
2.4. Statistical Analysis
3. Results
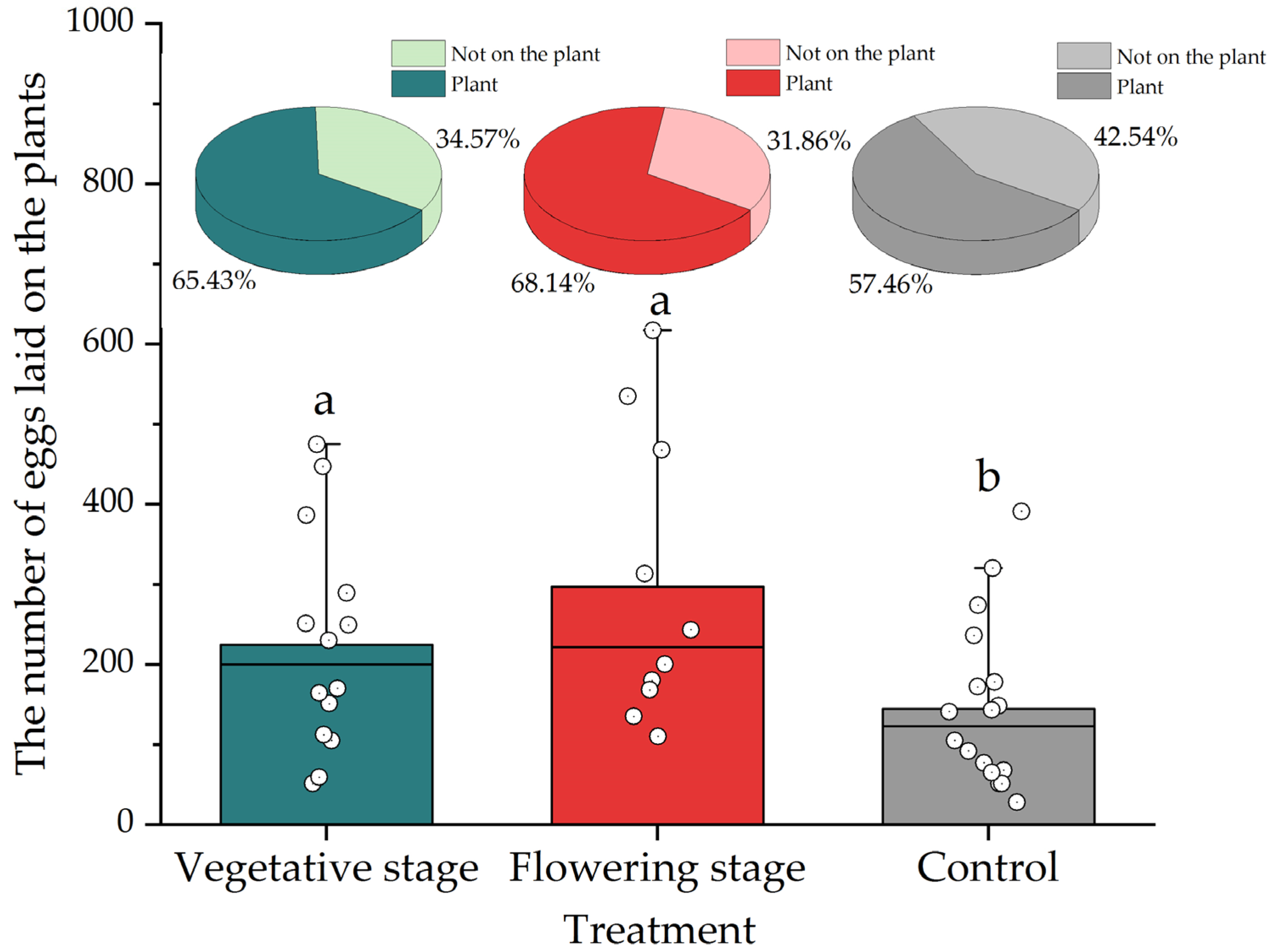
3.1. Laboratory Trials
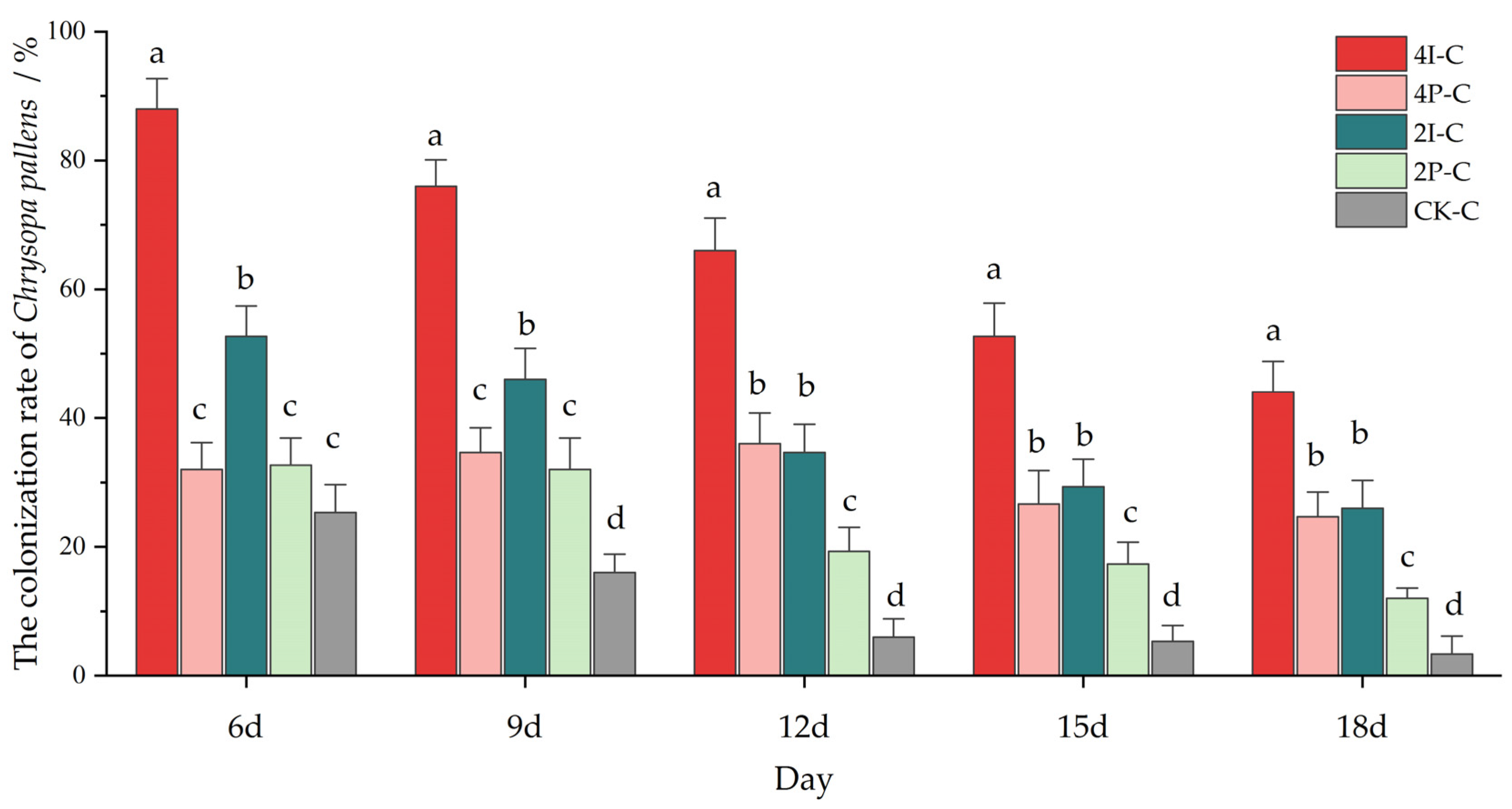
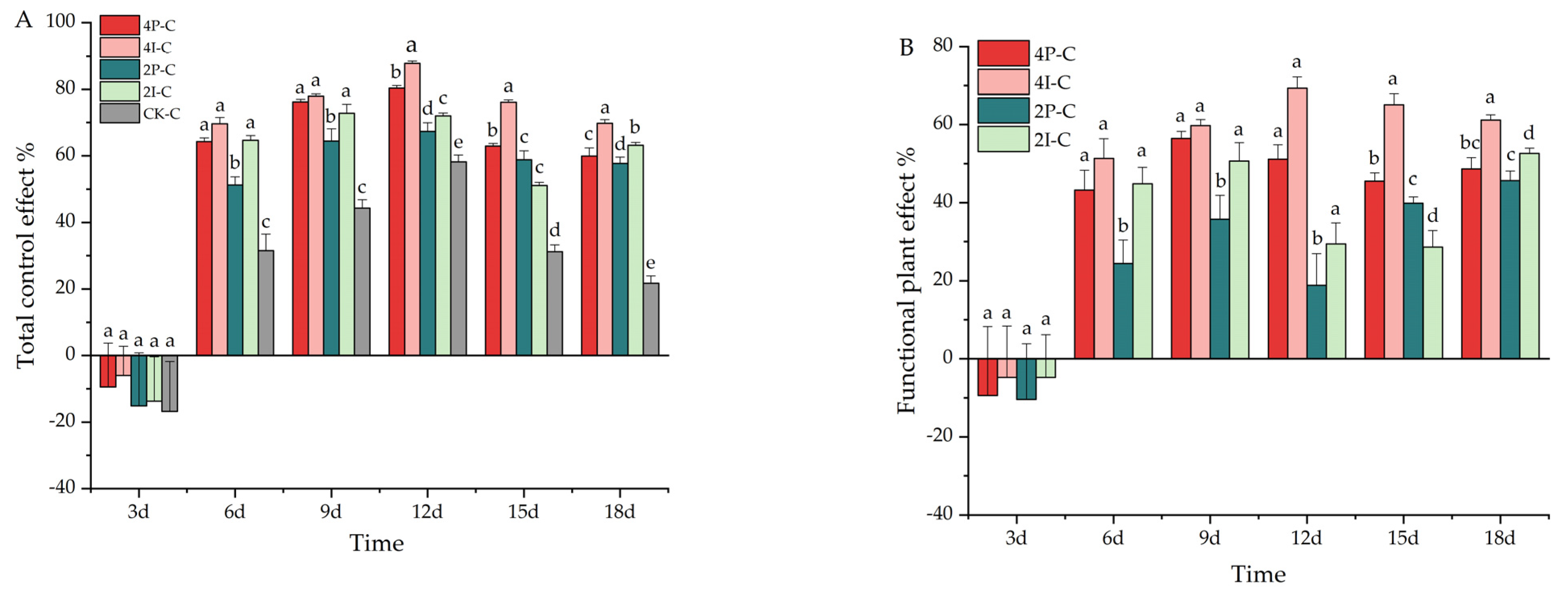
3.2. Greenhouse Assays
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heimpel, G.E.; Mills, N.J. Biological Control: Ecology and Applications; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Kabir, E.; Jahan, S.A. Exposure to pesticides and the associated human health effects. Sci. Total Environ. 2017, 575, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Goldson, S.L.; Wratten, S.D.; Ferguson, C.M.; Gerard, P.J.; Barratt, B.I.P.; Hardwick, S.; McNeill, M.R.; Phillips, C.B.; Popay, A.J.; Tylianakis, J.M.; et al. If and when successful classical biological control fails. Biol. Control 2014, 72, 76–79. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M.S. Habitat management to suppress pest populations: Progress and prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.X.; Jaworski, C.C.; Desneux, N.; Zhang, F.; Wang, S. Long-term and large-scale releases of Trichogramma promote pesticide decrease in maize in northeastern China. Entomol. Gen. 2020, 40, 331–335. [Google Scholar] [CrossRef]
- Zang, L.S.; Wang, S.; Zhang, F.; Desneux, N. Biological control with Trichogramma in China: History, present status and perspectives. Annu. Rev. Entomol. 2021, 66, 463–484. [Google Scholar] [CrossRef]
- Thomine, E.; Mumford, J.; Rusch, A.; Desneux, N. Using crop diversity to lower pesticide use: Socio-ecological approaches. Sci. Total Environ. 2022, 804, 150156. [Google Scholar] [CrossRef]
- González-Chang, M.; Tiwari, S.; Sharma, S.; Wratten, S.D. Habitat management for pest management: Limitations and prospects. Ann. Entomol. Soc. Am. 2019, 112, 302–317. [Google Scholar] [CrossRef]
- Damien, M.; Llopis, S.; Desneux, N.; Baaren, J.V.; Lann, C.L. How does floral nectar quality affect life history strategies in parasitic wasps? Entomol. Gen. 2020, 40, 147–156. [Google Scholar] [CrossRef]
- Wang, S.; Chen, X.; Li, Y.; Pan, B.; Wang, S.; Dai, H.; Wang, S.; Tang, B. Effects of changing temperature on the physiological and biochemical properties of Harmonia axyridis larvae. Entomol. Gen. 2020, 40, 229–241. [Google Scholar] [CrossRef]
- Ardanuy, A.; Figueras, M.; Matas, M.; Arnó, J.; Agustí, N.; Alomar, Ò.; Albajes, R.; Gabarra, R. Banker plants and landscape composition influence colonisation precocity of tomato greenhouses by mirid predators. J. Pest Sci. 2022, 95, 447–459. [Google Scholar] [CrossRef]
- Perovic, D.J.; Gámez-Virués, S.; Landis, D.; Tscharntke, T.; Zalucki, M.P.; Saura, S.; Furlong, M.J.; Desneux, N.; Sciarretta, A.; Balkenhol, N.; et al. Broadening the scope of empirical studies to answer persistent questions in landscape-moderated effects on biodiversity and ecosystem functioning. Adv. Ecol. Res. 2022, 65, 109–131. [Google Scholar]
- Lu, Z.X.; Zhu, P.Y.; Gurr, G.M.; Zheng, X.S.; Read, D.M.; Heong, K.L.; Yang, Y.J.; Xu, H.X. Mechanisms for flowering plants to benefit arthropod natural enemies of insect pests: Prospects for enhanced use in agriculture. Insect Sci. 2014, 21, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, P.; Lavoir, A.V.; Rodriquez-Saona, C.; Desneux, N. Bottom-up forces in agroecosystems and their potential impact on arthropod pest management. Annu. Rev. Entomol. 2022, 67, 239–259. [Google Scholar] [CrossRef]
- Parolin, P.; Bresch, C.; Desneux, N.; Brun, R.; Bout, A.; Boll, R.; Poncet, C. Secondary plants used in biological control: A review. Int. J. Pest Manag. 2012, 58, 91–100. [Google Scholar] [CrossRef]
- Holland, J.M.; Bianchi, F.; Entling, M.H.; Moonen, A.; Smith, B.M.; Jeanneret, P. Structure, function and management of semi-natural habitats for conservation biological control: A review of European studies. Pest Manag. Sci. 2016, 72, 1638–1651. [Google Scholar] [CrossRef]
- Araj, S.E.; Shields, M.W.; Wratten, S.D. Weed floral resources and commonly used insectary plants to increase the efficacy of a whitefly parasitoid. BioControl 2019, 64, 553–561. [Google Scholar] [CrossRef]
- Rodríguez, E.; González, M.; Paredes, D.; Campos, M.; Benítez, E. Selecting native perennial plants for ecological intensification in Mediterranean greenhouse horticulture. Bull. Entomol. Res. 2017, 108, 694–704. [Google Scholar] [CrossRef] [Green Version]
- Belén, C.; Mónica, G.; Emilio, B.; Eva, D.M.; Gemma, C.O.; Mercedes, C.; Estefanía, R. Spider communities and biological control in native habitats surrounding greenhouses. Insects 2018, 9, 33. [Google Scholar]
- Frank, S.D. Biological control of arthropod pests using banker plant systems: Past progress and future directions. Biol. Control 2010, 52, 8–16. [Google Scholar] [CrossRef]
- Chen, X.; Jaworski, C.C.; Dai, H.J.; Liang, Y.Y.; Guo, X.J.; Wang, S.; Zang, L.S.; Desneux, N. Combining banker plants to achieve long-term pest control in multi-pest and multi-natural enemy cropping systems. J. Pest Sci. 2022, 95, 685–697. [Google Scholar] [CrossRef]
- Togni, P.H.B.; Venzon, M.; Muniz, C.A.; Martins, E.F.; Pallini, A.; Sujii, E.R. Mechanisms underlying the innate attraction of an aphidophagous coccinellid to coriander plants: Implications for conservation biological control. Biol. Control 2016, 92, 77–84. [Google Scholar] [CrossRef]
- Ju, Q.; Ouyang, F.; Gu, S.; Qiao, F.; Yang, Q.; Qu, M.; Ge, F. Strip intercropping peanut with maize for peanut aphid biological control and yield enhancement. Agric. Ecosyst. Environ. 2019, 286, 106682. [Google Scholar] [CrossRef]
- Ouyang, F.; Su, W.W.; Zhang, Y.S.; Liu, X.H.; Su, J.; Zhang, Q.Q.; Men, X.Y.; Ju, Q.; Ge, F. Ecological control service of the predatory natural enemy and its maintaining mechanism in rotation-intercropping ecosystem via wheat-maize-cotton. Agric. Ecosyst. Environ. 2020, 301, 107024. [Google Scholar] [CrossRef]
- Wang, J.; Li, S.; Yang, J.; Guo, M.C.; Dai, H.J.; Ramirez-Romero, R.; Jin, Z.Y.; Wang, S. The fitness of mass rearing food on the establishment of Chrysopa pallens in a banker plant system under fluctuating temperature conditions. Insects 2021, 12, 1014. [Google Scholar] [CrossRef] [PubMed]
- Tauber, M.J.; Tauber, C.A.; Daane, K.M.; Hagen, K.T.S. Commercialization of predators: Recent lessons from green lacewings (Neuroptera: Chrysopidae: Chrosoperla). Am. Entomol. 2000, 46, 26–38. [Google Scholar] [CrossRef]
- Wu, H.H.; Zhang, L.S.; Chen, H.Y. Effect of temperature and release height on dispersal behavior of Chrysopa pallens (Rambur) and Chrysopa formosa Brauer adults. Chin. J. Biol. Control 2014, 30, 587–592. [Google Scholar]
- Yang, Y.J.; Li, S.; Fang, Y.; Wang, S.; Yue, Y.L.; Gu, J.J. The olfactory behavioral response of Chrysopa pallens to aromatic plant Ocimum basilicum and its major volatiles in different periods. J. Environ. Entomol. Accepted, to be published. 2022. [Google Scholar]
- Kéita, S.; Vincent, C.; Schmit, J.P.; Arnason, J.T.; Bélanger, A. Efficacy of essential oil of Ocimum basilicum L. and O. gratissimum L. applied as an insecticidal fumigant and powder to control Callosobruchus maculatus (Fab.). J. Stored Prod. Res. 2001, 37, 339–349. [Google Scholar] [CrossRef]
- Maukanovi-Joci, M.P.; Rani, D.V.; Stevanovi, Z.P.D. Floral nectaries of basil (Ocimum basilicum): Morphology, anatomy and possible mode of secretion. South Afr. J. Bot. 2007, 73, 636–641. [Google Scholar] [CrossRef] [Green Version]
- Hullé, M.; Chaubet, B.; Turpeau, E.; Simon, J.C. Encyclop’ aphid: A website on aphids and their natural enemies. Entomol. Gen. 2020, 40, 97–101. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils--a review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, A.K.; Landis, D.A. Attractiveness of michigan native plants to arthropod natural enemies and herbivores. Environ. Entomol. 2007, 36, 751–765. [Google Scholar] [CrossRef] [PubMed]
- Damien, M.; Lann, C.L.; Desneux, N.; Alford, L.; Hassan, D.A.; Georges, R.; Baaren, J.V. Flowering cover crops in winter increase pest control but not trophic link diversity. Agric. Ecosyst. Environ. 2017, 247, 418–425. [Google Scholar] [CrossRef] [Green Version]
- Wäckers, F.L.; van Rijn, P.C.J. Pick and Mix: Selecting flowering plants to meet the requirements of target biological control insects. In Biodiversity and Insect Pests; Gurr, G.M., Wratten, S.D., Snyder, W.E., Read, D., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2012; pp. 139–165. [Google Scholar]
- Mathews, C.R.; Brown, M.W.; Wackers, F.L. Comparison of peach cultivars for provision of extrafloral nectar resources to Harmonia axyridis (Coleoptera: Coccinellidae). Environ. Entomol. 2016, 45, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Snyder, W.E. Give predators a complement: Conserving natural enemy biodiversity to improve biocontrol. Biol. Control 2019, 135, 73–82. [Google Scholar] [CrossRef]
- Berkvens, N.; Bonte, J.; Berkvens, D.; Tirry, L.; De Clercq, P. Influence of diet and photoperiod on development and reproduction of European populations of Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae). BioControl 2007, 53, 211–221. [Google Scholar] [CrossRef]
- Hatt, S.; Osawa, N. The role of Perilla frutescens flowers on fitness traits of the ladybird beetle Harmonia axyridis. BioControl 2019, 64, 381–390. [Google Scholar] [CrossRef]
- Evans, E.W.; Stevenson, A.T.; Richards, D.R. Essential versus alternative foods of insect predators benefits of a mixed diet. Oecologia 1999, 121, 107–112. [Google Scholar]
- Robinson, K.A.; Jonsson, M.; Wratten, S.D.; Wade, M.R.; Buckley, H.L. Implications of floral resources for predation by an omnivorous lacewing. Basic Appl. Ecol. 2008, 9, 172–181. [Google Scholar] [CrossRef]
- Rahat, S.; Gurr, G.M.; Wratten, S.D.; Mo, J.; Neeson, R. Effect of plant nectars on adult longevity of the stinkbug parasitoid, Trissolcus basalis. Int. J. Pest Manag. 2005, 51, 321–324. [Google Scholar] [CrossRef]
- Naranjo, S.E.; Gibson, R.L. Phytophagy in predaceous Heteroptera: Effects on life history and population dynamics. In Zoophytophagous Heteroptera: Implications for Life History and Integrated Pest Management, Thomas Say Symposium Proceedings; Wiedenmann, R., Alomar, O., Eds.; Entomological Society of America: Lanham, MD, USA, 1996; pp. 57–93. [Google Scholar]
- Wang, J.; Li, S.; Fang, Y.; Zhang, F.; Jin, Z.Y.; Desneux, N.; Wang, S. Enhanced and sustainable control of Myzus persicae by repellent plants in organic pepper and eggplant greenhouses. Pest Manag. Sci. 2022, 78, 428–437. [Google Scholar] [CrossRef] [PubMed]
- van Rijn, P.C.J.; Wäckers, F.L.; Cadotte, M. Nectar accessibility determines fitness, flower choice and abundance of hoverflies that provide natural pest control. J. Appl. Ecol. 2016, 53, 925–933. [Google Scholar] [CrossRef] [Green Version]
- Riddick, E.W. Volatile and non-volatile organic compounds stimulate oviposition by aphidophagous predators. Insects 2020, 11, 683. [Google Scholar] [CrossRef] [PubMed]
- Herrera, R.A.; Ruano, F.; Ramírez, C.; Frischie, S.; Campos, M. Attraction of green lacewings (Neuroptera: Chrysopidae) to native plants used as ground cover in woody Mediterranean agroecosystems. Biol. Control 2019, 139, 104066. [Google Scholar] [CrossRef] [Green Version]
- Reddy, G.V.P. Plant volatiles mediate orientation and plant preference by the predator Chrysoperla carnea Stephens (Neuroptera: Chrysopidae). Biol. Control 2002, 25, 49–55. [Google Scholar] [CrossRef]
- Hatt, S.; Uytenbroeck, R.; Lopes, T.C.M.; Mouchon, P.; Francis, F. Identification of flower functional traits affecting abundance of generalist predators in perennial multiple species wildflower strips. Arthropod-Plant Interact. 2019, 13, 127–137. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Pallini, A.; Bernardo, A.; Veiga, V.R.; Janssen, A. Associative learning in immature lacewings (Ceraeochrysa cubana). Entomol. Exp. Appl. 2019, 157, 775–783. [Google Scholar] [CrossRef] [Green Version]
- Batista, M.C.; Fonseca, M.C.M.; Teodoro, A.V.; Martins, E.F.; Pallini, A.; Venzon, M. Basil (Ocimum basilicum L.) attracts and benefits the green lacewing Ceraeochrysa cubana Hagen. Biol. Control 2017, 110, 98–106. [Google Scholar] [CrossRef]
- Lundgren, J.G.; Wyckhuys, K.; Desneux, N. Population responses by Orius insidiosus to vegetational diversity. BioControl 2009, 54, 135–142. [Google Scholar] [CrossRef]
- Vankosky, M.A.; VanLaerhoven, S.L. Does host plant quality affect the oviposition decisions of an omnivore? Insect Sci. 2017, 24, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, B.A.; Cottrell, T.E. Oviposition response of green lacewings (Neuroptera: Chrysopidae) to aphids (Hemiptera: Aphididae) and potential attractants on pecan. Environ. Entomol. 2007, 36, 577–583. [Google Scholar] [CrossRef] [Green Version]
- Keulder, R.; Van, D. Patterns of lacewing (Neuroptera: Chrysopidae) flight activity, flight height and spatial distribution of eggs on maize plants. Afr. Entomol. 2013, 21, 95–102. [Google Scholar] [CrossRef]
- Gontijo, L.M. Engineering natural enemy shelters to enhance conservation biological control in field crops. Biol. Control 2019, 130, 155–163. [Google Scholar] [CrossRef]
- Balzan, M.V.; Wäckers, F.L. Flowers to selectively enhance the fitness of a host-feeding parasitoid: Adult feeding by Tuta absoluta and its parasitoid Necremnus artynes. Biol. Control 2013, 67, 21–31. [Google Scholar] [CrossRef]
- Begg, G.S.; Cook, S.M.; Dye, R.; Ferrante, M.; Franck, P.; Lavigne, C.; Mansion-Vaquie, A.; Pell, J.K.; Petit, S. A functional overview of conservation biological control. Crop Prot. 2016, 97, 145–158. [Google Scholar] [CrossRef]
- Zhang, X.M.; Lövei, G.L.; Ferrante, M.; Yang, N.W.; Wan, F.H. The potential of trap and barrier cropping to decrease densities of the whitefly Bemisia tabaci MED on cotton in China. Pest Manag. Sci. 2020, 76, 366–374. [Google Scholar] [CrossRef]
- Song, B.Z.; Zhang, J.; Hu, J.H.; Wu, H.Y.; Kong, Y.; Yao, Y.C. Temporal dynamics of the arthropod community in pear orchards intercropped with aromatic plants. Pest Manag. Sci. 2011, 67, 1107–1114. [Google Scholar]
- Collard, B.; Tixier, P.; Carval, D.; Lavigne, C.; Delattre, T.J.E.M. Spatial organisation of habitats in agricultural plots affects per-capita predator effect on conservation biological control: An individual based modelling study. Ecol. Model. 2018, 388, 124–135. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day | Source | Lacewings | Aphids | ||||
|---|---|---|---|---|---|---|---|
| F | df | p-Value | F | df | p-Value | ||
| 3 d | Spatiality | 162.45 | 1 | <0.001 | 33.9 | 1 | <0.001 |
| Density | 33.8 | 1 | <0.001 | 30.1 | 1 | <0.001 | |
| Spatiality × Density | 36.45 | 1 | <0.001 | 6.37 | 1 | 0.014 | |
| 9 d | Spatiality | 87.678 | 1 | <0.001 | 5.33 | 1 | 0.025 |
| Density | 30.558 | 1 | <0.001 | 14.4 | 1 | <0.001 | |
| Spatiality × Density | 21.395 | 1 | <0.001 | 2.39 | 1 | 0.128 | |
| 12 d | Spatiality | 56.886 | 1 | <0.001 | 14.1 | 1 | <0.001 |
| Density | 63.775 | 1 | <0.001 | 80.8 | 1 | <0.001 | |
| Spatiality × Density | 5.954 | 1 | 0.018 | 0.56 | 1 | 0.457 | |
| 15 d | Spatiality | 38.811 | 1 | <0.001 | 2.9 | 1 | 0.094 |
| Density | 28.681 | 1 | <0.001 | 74.9 | 1 | <0.001 | |
| Spatiality × Density | 5.268 | 1 | 0.025 | 38.2 | 1 | <0.001 | |
| 18 d | Spatiality | 42.169 | 1 | <0.001 | 27 | 1 | <0.001 |
| Density | 35.692 | 1 | <0.001 | 9.32 | 1 | 0.003 | |
| Spatiality × Density | 1.08 | 1 | 0.303 | 2.32 | 1 | 0.133 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Y.; Li, S.; Xu, Q.; Wang, J.; Yang, Y.; Mi, Y.; Jin, Z.; Desneux, N.; Wang, S. Optimizing the Use of Basil as a Functional Plant for the Biological Control of Aphids by Chrysopa pallens (Neuroptera: Chrysopidae) in Greenhouses. Insects 2022, 13, 552. https://doi.org/10.3390/insects13060552
Fang Y, Li S, Xu Q, Wang J, Yang Y, Mi Y, Jin Z, Desneux N, Wang S. Optimizing the Use of Basil as a Functional Plant for the Biological Control of Aphids by Chrysopa pallens (Neuroptera: Chrysopidae) in Greenhouses. Insects. 2022; 13(6):552. https://doi.org/10.3390/insects13060552
Chicago/Turabian StyleFang, Yan, Shu Li, Qingxuan Xu, Jie Wang, Yajie Yang, Yingying Mi, Zhenyu Jin, Nicolas Desneux, and Su Wang. 2022. "Optimizing the Use of Basil as a Functional Plant for the Biological Control of Aphids by Chrysopa pallens (Neuroptera: Chrysopidae) in Greenhouses" Insects 13, no. 6: 552. https://doi.org/10.3390/insects13060552