
Toxicity and Residual Activity of Insecticides against Diadegma insulare, a Parasitoid of the Diamondback Moth

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Parasitoids
2.2. Laboratory Bioassay (Acute Toxicity)
2.3. Residual Activity (Persistence) of Insecticides to D. insulare
2.4. Analysis of Data
3. Results
3.1. Laboratory Bioassay (Acute Toxicity)
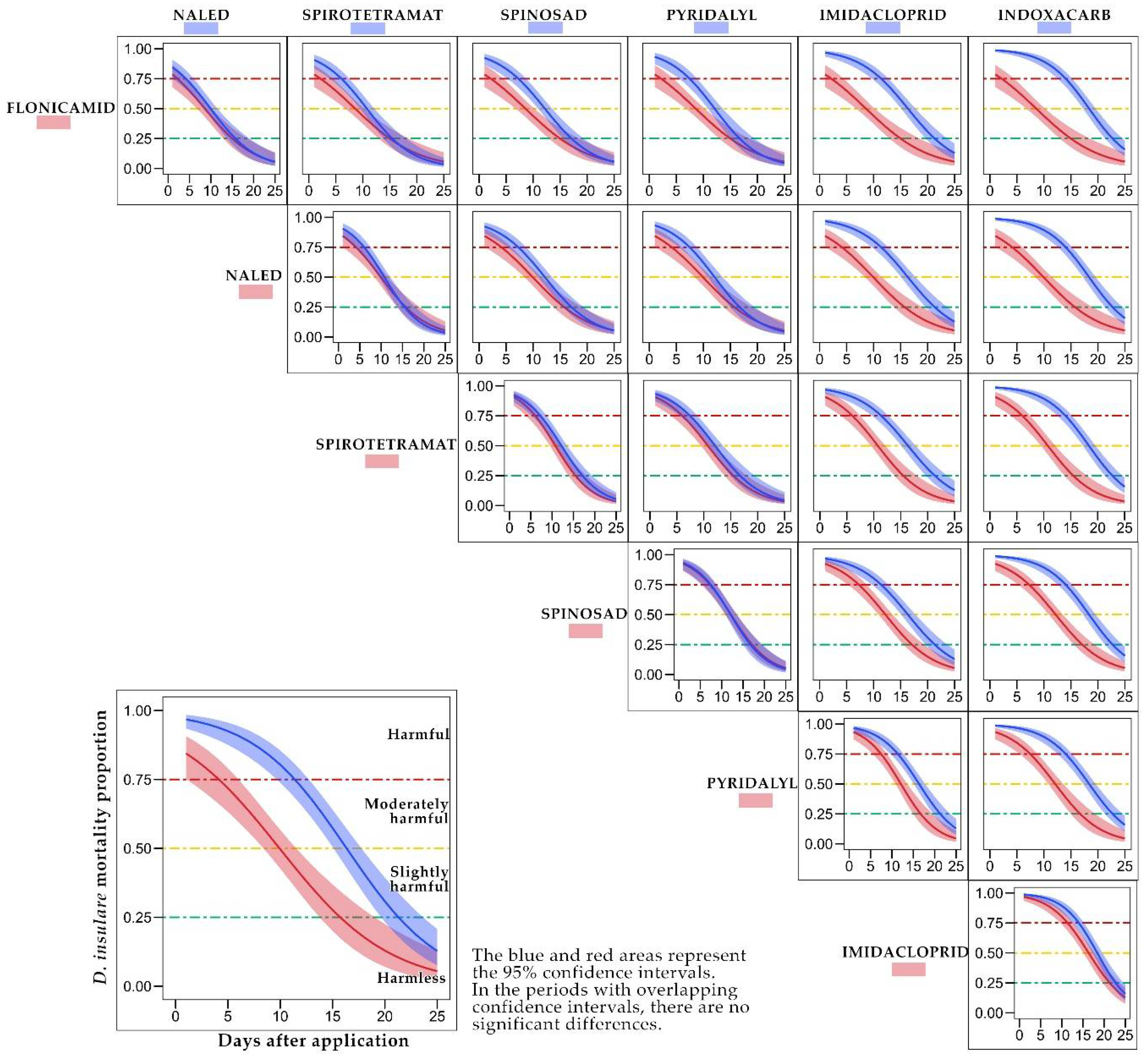
3.2. Residual Activity (Persistence) of Insecticides to D. insulare
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, S.-S.; Chen, F.-Z.; Zalucki, M. Development and Survival of the Diamondback Moth (Lepidoptera: Plutellidae) at Constant and Alternating Temperatures. Environ. Èntomol. 2002, 31, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Furlong, M.J.; Ju, K.H.; Su, P.W.; Chol, J.K.; Il, R.C.; Zalucki, M. Integration of endemic natural enemies and Bacillus thuringiensis to manage insect pests of Brassica crops in North Korea. Agric. Ecosyst. Environ. 2008, 125, 223–238. [Google Scholar] [CrossRef]
- Sayani, Z.; Mikani, A.; Mosallanejad, H. Biochemical Resistance Mechanisms to Fenvalerate in Plutella xylostella (Lepidoptera: Plutellidae). J. Econ. Èntomol. 2019, 112, 1372–1377. [Google Scholar] [CrossRef]
- Xu, J.; Shelton, A.M.; Cheng, X. Comparison of Diadegma insulare (Hymenoptera: Ichneumonidae) and Microplitis plutellae (Hymenoptera: Braconidae) as Biological Control Agents of Plutella xylostella (Lepidoptera: Plutellidae): Field Parasitism, Insecticide Susceptibility, and Host-Searching. J. Econ. Èntomol. 2001, 94, 14–20. [Google Scholar] [CrossRef]
- Cordero, R.J.; Bloomquist, J.R.; Kuhar, T.P. Susceptibility of two diamondback moth parasitoids, Diadegma insulare (Cresson) (Hymenoptera; Ichneumonidae) and Oomyzus sokolowskii (Kurdjumov) (Hymenoptera; Eulophidae), to selected commercial insecticides. Biol. Control 2007, 42, 48–54. [Google Scholar] [CrossRef]
- Cerna, E.; Ail, C.; Landeros, J.; Sánchez, S.; Badii, M.; Aguirre, L.; Ochoa, Y. Comparison of toxicity and selectivity of the pest Bactericera cockerelli and Its Predator Chrysoperla carnea. Agrociencia 2012, 46, 783–793. [Google Scholar]
- Jafary-Jahed, M.; Razmjou, J.; Nouri-Ganbalani, G.; Naseri, B.; Hassanpour, M.; Leppla, N.C. Life Table Parameters and Oviposition Preference ofPlutella xylostella(Lepidoptera: Plutellidae) on Six Brassicaceous Crop Plants. J. Econ. Èntomol. 2018, 112, 932–938. [Google Scholar] [CrossRef]
- Díaz-Gomez, O.; Rodríguez, J.C.; Shelton, A.M.; Lagunes-T, A.; Bujanos-M, R. Susceptibility of Plutella xylostella (L.) (Lepidoptera: Plutellidae) Populations in Mexico to Commercial Formulations of Bacillus thuringiensis. J. Econ. Èntomol. 2000, 93, 963–970. [Google Scholar] [CrossRef] [Green Version]
- Santoyo, J.A.; Martínez, C.O. Tecnología de Producción de Brócoli; Folleto Técnico; Fundación Produce Sinaloa A. C.: Sinaloa, México, 2012; p. 29. [Google Scholar]
- Xu, Y.-Y.; Liu, T.-X.; Leibee, G.L.; Jones, W.A. Effects of selected insecticides on Diadegma insulare (Hymenoptera: Ichneumonidae), a parasitoid of Plutella xylostella(Lepidoptera: Plutellidae). Biocontrol Sci. Technol. 2004, 14, 713–723. [Google Scholar] [CrossRef]
- Pérez, M.A.; Navarro, H.; Miranda, E. Residuos de plaguicidas en hortalizas: Problemática y riesgo en México. Rev. Int. Contam. Ambient. 2013, 29, 45–64. [Google Scholar]
- Liu, X.; Chen, M.; Collins, H.L.; Onstad, D.; Roush, R.; Zhang, Q.; Shelton, A.M. Effect of insecticides and Plutella xylostella (Lepidoptera: Plutellidae) genotype on a predator and parasitoid and implications for the evolution of insecticide resistance. J. Econ. Èntomol. 2012, 105, 354–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bujanos Muñiz, R.; Marín Jarillo, A.; Díaz Espino, L.F.; Gámez Vázquez, A.J.; Ávila Perches, M.Á.; Herrera Vega, R.; Dorantes González, J.R.A.; Gámez Vázquez, F.P. Manejo Integrado de la Palomilla Dorso de Diamante Plutella xylostella (L.) en la Región del Bajío, México. Celaya, Gto.; Informe Técnico 27; Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias: Celya Guanajuato, México, 2013; p. 44. [Google Scholar]
- Gong, W.; Yan, H.-H.; Gao, L.; Guo, Y.-Y.; Xue, C.-B. Chlorantraniliprole Resistance in the Diamondback Moth (Lepidoptera: Plutellidae). J. Econ. Èntomol. 2014, 107, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Wang, C.-H.; Wang, Y.-Y.; Sun, S.-Q.; Wang, H.-H.; Xue, C.-B. Resistance to Diamide Insecticides in Plutella xylostella (Lepidoptera: Plutellidae): Comparison Between Lab-Selected Strains and Field-Collected Populations. J. Econ. Èntomol. 2018, 111, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, J.; Cao, X.; Wang, F.; Yang, Y.; Wu, S.; Wu, Y. Long-term monitoring and characterization of resistance to chlorfenapyr in Plutella xylostella(Lepidoptera: Plutellidae) from China. Pest Manag. Sci. 2018, 75, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-Z.; Li, Y.-X.; Collins, H.L.; Gusukuma-Minuto, L.; Mau, R.F.L.; Thompson, G.D.; Shelton, A.M. Monitoring and Characterization of Diamondback Moth (Lepidoptera: Plutellidae) Resistance to Spinosad. J. Econ. Èntomol. 2002, 95, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Sarfraz, M.; Keddie, A.B.; Dosdall, L.M. Biological control of the diamondback moth, Plutella xylostella: A review. Biocontrol Sci. Technol. 2005, 15, 763–789. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Wu, S.; Yang, T.; Zhu, C.; Gao, C. Monitoring Field Populations of Plutella xylostella(Lepidoptera: Plutellidae) for Resistance to Eight Insecticides in China. Fla. Èntomol. 2015, 98, 65–73. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, X.; Shen, J.; Mao, K.; You, H.; Li, J. Susceptibility of field populations of the diamondback moth, Plutella xylostella, to a selection of insecticides in Central China. Pestic. Biochem. Physiol. 2016, 132, 38–46. [Google Scholar] [CrossRef]
- Bommarco, R.; Miranda, F.; Bylund, H.; Björkman, C. Insecticides Suppress Natural Enemies and Increase Pest Damage in Cabbage. J. Econ. Èntomol. 2011, 104, 782–791. [Google Scholar] [CrossRef]
- Karimzadeh, J.; Besharatnejad, M.H. Ecological control of Plutella xylostella (Lepidoptera, Plutellidae) using trap cropping and Bt applications. Arch. Phytopathol. Plant Prot. 2019, 52, 1326–1347. [Google Scholar] [CrossRef]
- Löhr, B.; Kfir, R. Diamondback Moth Plutella xylostella (L.) in Africa. A Review with Emphasis on Biological Control. Available online: https://agritrop.cirad.fr/519637/ (accessed on 8 March 2022).
- Martínez-Castillo, M.; Leyva, J.; Cibrian-Tovar, J.; Bujanos-Muñiz, R. Parasitoid diversity and impact on populations of the diamondback moth Plutella xylostella (L.) on Brassica crops in central México. BioControl 2002, 47, 23–31. [Google Scholar] [CrossRef]
- Álvarez-Cabrera, G.M. Propuesta de Ichneumónidos (Hymenoptera) para el control biológico de insectos plaga en México. Agro Product. 2017, 10, 78–83. [Google Scholar]
- Tovar Hernández, H.; Bautista Martínez, N.; Vera Graziano, J.; Suárez Vargas, A.D.; Ramírez Alarcón, S. Fluctuación poblacional y parasitismo de larvas de Copitarsia decolora Guenée, Plutella xylostella L. y Trichoplusia ni Hübner (Lepidoptera) en Brassica oleracea L. Acta Zoológica Mex. 2007, 23, 183–196. [Google Scholar]
- Hermansson, J. Biology of the Diamondback Moth (Plutella xylostella) and Its Future Impact in Swedish Oilseed Rape Production. Available online: https://stud.epsilon.slu.se/9645/ (accessed on 7 March 2022).
- Barrios-Díaz, B.; Alatorre-Rosas, R.; Calyecac-Cortero, H.G.; Bautista-Martínez, N. Identificación y fluctuación poblacional de plagas de col (Brassica oleracea var. capitata) y sus enemigos naturales en Acatzingo, Puebla. Agrociencia 2004, 38, 239–248. [Google Scholar]
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The Sublethal Effects of Pesticides on Beneficial Arthropods. Annu. Rev. Èntomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Cloyd, R. Indirect Effects of Pesticides on Natural Enemies. In Pesticides-Advances in Chemical and Botanical Pesticides; Soundararajan, R.P., Ed.; IntechOpen: London, UK, 2012; pp. 127–150. [Google Scholar] [CrossRef]
- Idris, A.B.; Grafius, E. Differential Toxicity of Pesticides to Diadegma insulate (Hymenoptera: Ichneumonidae) and Its Host, the Diamondback Moth (Lepidoptera: Plutellidae). J. Econ. Èntomol. 1993, 86, 529–536. [Google Scholar] [CrossRef]
- Sourakov, A.; Mitchell, E. A wasp parasitoid, Diadegma insulare (Cresson) (Insecta: Hymenotera: Ichneumonidae). Available online: https://edis.ifas.ufl.edu/publication/IN281 (accessed on 8 March 2022).
- Hill, T.A.; Foster, R.E. Effect of Insecticides on the Diamondback Moth (Lepidoptera: Plutellidae) and Its Parasitoid Diadegma insulare (Hymenoptera: Ichneumonidae). J. Econ. Èntomol. 2000, 93, 763–768. [Google Scholar] [CrossRef]
- Hill, T.A.; Foster, R.E. Influence of Selected Insecticides on the Population Dynamics of Diamondback Moth (Lepidoptera: Plutellidae) and Its Parasitoid, Diadegma insulare (Hymenoptera: Ichneumonidae), in Cabbage. J. Èntomol. Sci. 2003, 38, 59–71. [Google Scholar] [CrossRef]
- Morales, F.; Lagunes, A.; Rodriguez, C. Comparación de cuatro métodos de bioensayo en la determinación de la toxicidad a insecticidas en Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). Entomotropica 2015, 30, 227–235. [Google Scholar]
- Luna-Cruz, A.; Rodríguez-Leyva, E.; Lomeli-Flores, J.R.; Ortega-Arenas, L.D.; Bautista-Martínez, N.; Pineda, S. Toxicity and Residual Activity of Insecticides Against Tamarixia triozae(Hymenoptera: Eulophidae), a Parasitoid of Bactericera cockerelli (Hemiptera: Triozidae). J. Econ. Èntomol. 2015, 108, 2289–2295. [Google Scholar] [CrossRef]
- Hassan, S.A. Guidelines for testing the effects of pesticides on beneficial organisms. IOBC/WPRS Bull. 1992, 15, 1–186. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2020; Available online: https://www.r-project.org/ (accessed on 14 January 2022).
- Hassan, S.A.; Bigler, F.; Bogenschütz, H.; Boller, E.; Brun, J.; Calis, J.N.M.; Coremans-Pelseneer, J.; Duso, C.; Grove, A.; Heimbach, U.; et al. Results of the sixth joint pesticide testing programme of the IOBC/WPRS-working group pesticides and beneficial organisms. EBioContromophagal 1994, 39, 107–119. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Èntomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Williams, T.; Valle, J.; Viñuela, E. Is the Naturally Derived Insecticide Spinosad Compatible with Insect Natural Enemies? Biocontrol. Sci. Technol. 2003, 13, 459–475. [Google Scholar] [CrossRef]
- Bostanian, N.J.; Akalach, M. The contact toxicity of indoxacarb and five other insecticides toOrius insidiosus (Hemiptera: Anthocoridae) andAphidius colemani (Hymenoptera: Braconidae), beneficials used in the greenhouse industry. Pest Manag. Sci. 2004, 60, 1231–1236. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, N.; Saito, S.; Hirose, T.; Suzuki, M.; Matsuo, S.; Izumi, K.; Nagatomi, T.; Ikegami, H.; Umeda, K.; Tsushima, K.; et al. The discovery of pyridalyl: A novel insecticidal agent for controlling lepidopterous pests. Pest Manag. Sci. 2003, 60, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Isayama, S.; Saito, S.; Kuroda, K.; Umeda, K.; Kasamatsu, K. Pyridalyl, a novel insecticide: Potency and insecticidal selectivity. Arch. Insect Biochem. Physiol. 2005, 58, 226–233. [Google Scholar] [CrossRef]
- Francesena, N.; Haramboure, M.; Smagghe, G.; Stadler, T.; Schneider, M.I. Preliminary studies of effectiveness and selectivity of Movento on Bemisia tabaci and its parasitoid Eretmocerus mundus. Commun. Agric. Appl. Biol. Sci. 2012, 77, 727–733. [Google Scholar]
- Mani, M. Hundred and sixty years of Australian lady bird beetle Crypotolaemus montrouzieri Mulsant—A global view. Biocontrol Sci. Technol. 2018, 28, 938–952. [Google Scholar] [CrossRef]
- Martinou, A.; Seraphides, N.; Stavrinides, M. Lethal and behavioral effects of pesticides on the insect predator Macrolophus pygmaeus. Chemosphere 2013, 96, 167–173. [Google Scholar] [CrossRef]
- Lira, A.C.S.; Zanardi, O.Z.; Beloti, V.H.; Bordini, G.P.; Yamamoto, P.T.; Parra, J.R.P.; Carvalho, G.A. Lethal and Sublethal Impacts of Acaricides on Tamarixia radiata (Hemiptera: Eulophidae), an Important Ectoparasitoid of Diaphorina citri (Hemiptera: Liviidae). J. Econ. Èntomol. 2015, 108, 2278–2288. [Google Scholar] [CrossRef] [PubMed]
- IRAC. Interactive MoA Classification | Insecticide Resistance Action Committee (IRAC). Available online: https://irac-online.org/modes-of-action/ (accessed on 7 March 2022).
- Tudi, M.; Ruan, H.D.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D. Agriculture Development, Pesticide Application and Its Impact on the Environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef] [PubMed]
- Wigglesworth, V.B. Insect Physiology; Methuen & Co., Ltd.: London, UK, 1966; pp. 1–14. [Google Scholar]
- Biondi, A.; Mommaerts, V.; Smagghe, G.; Viñuela, E.; Zappalà, L.; Desneux, N. The non-target impact of spinosyns on beneficial arthropods. Pest Manag. Sci. 2012, 68, 1523–1536. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.G.; Nguyen, R. Toxicity of pesticides to Tamarixia radiata, a parasitoid of the Asian citrus psyllid. BioControl 2010, 55, 601–611. [Google Scholar] [CrossRef]
- Saunders, D.G.; Bret, B.L. Fate of spinosad in the environment. Down Earth 1997, 52, 14–20. [Google Scholar]
- Luna-Cruz, A.; Lomeli-Flores, J.R.; Rodríguez-Leyva, E.; Ortega-Arenas, L.D.; La Peña, A.H.-D. Toxicidad de cuatro insecticidas sobre Tamarixia triozae (Burks) (Hymenoptera: Eulophidae) y su hospedero Bactericera cockerelli (Sulc) (Hemiptera: Triozidae). Acta Zoológica Mex. 2011, 27, 509–526. [Google Scholar] [CrossRef] [Green Version]
- Crouse, G.D.; Sparks, T.C.; Schoonover, J.; Gifford, J.; Dripps, J.; Bruce, T.; Larson, L.L.; Garlich, J.; Hatton, C.; Hill, R.L.; et al. Recent advances in the chemistry of spinosyns. Pest Manag. Sci. 2001, 57, 177–185. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Active Ingredient (a.i.) | Commercial Name | Concentration (mg a.i. L−1) | Recommended Dose | Toxicological Group | Mode of Action |
|---|---|---|---|---|---|
| Emamectin Benzoate | Proclaim® 05 SG | 5.00 | 0.30–0.40 kg/ha | Avermectin | Glutamate-gated chloride channel allosteric modulators. Acts on the nervous system and muscle system. |
| Spinosad | Spintor® 12 SC | 44.20 | 0.10 L/ha | Spinosins | Nicotinic acetylcholine receptor allosteric modulators; acts on the central nervous system. |
| Spirotetramat | Movento® 150 OD | 15.30 | 0.3–0.4 L/ha | Tetronic acid | Inhibitors of acetyl CoA carboxylase; inhibit lipid biosynthesis and growth regulator. |
| Imidacloprid | Confidor® 350 SC | 35.00 | 0.15–0.30 L/ha | Neonicotinoid | Nicotinic acetylcholine receptor competitive modulators; acts on the nervous system. |
| Spinetoram | Palgus® | 5.87 | 0.25–0.30 L/ha | Spinosins | Nicotinic acetylcholine receptor allosteric modulators; acts on the central nervous system. |
| Indoxacarb | Avaunt® 150 EC | 15.84 | 0.30–0.50 L/ha | Oxadiacines | Voltage-dependent sodium channel blockers. Acts on the nervous system. |
| Flonicamid | Beleaf® | 50.00 | 0.10–0.25 kg/ha | Pirydinecarboxiamides | Chordotonal organ modulators undefined target site; acts on the nervous system. |
| Naled | Dibrom® 8 | 66.5 | 0.750–1.25 L/ha | Organophosphates | Acetylcholinesterase inhibitors; acts on the nervous system. |
| Pyridalyl | Pleo® 50 EC | 44.9 | 0.20–0.35 L/ha | Derivatives of dihaloprene | Compounds of an unknown or uncertain mode of action. |
| Treatment | Cumulative Mortality (%±EE) 1 | IOBC Toxicity Categories 2 at 72 h | ||
|---|---|---|---|---|
| 24 h | 48 h | 72 h | ||
| Indoxacarb | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | Highly harmful (Category 4, >99% mortality) |
| Emamectine benzoate | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | |
| Imidacloprid | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | |
| Naled | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | |
| Spinoteram | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | |
| Spinosad | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | |
| Flonicamid | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | |
| Spirotetramat | 26.0 ± 0.6 b | 64.0 ± 0.2 b | 64.0 ± 0.2 b | Slightly harmful (Category 2, 30–79% mortality) |
| Pyridalyl | 18.0 ± 0.3 b | 38.0 ± 0.2 c | 48.0 ± 0.5 c | |
| Water + InexA | 10.0 ± 0.0 c | 27.0 ± 0.2 d | 27.0 ± 0.2 d | Harmless (Category 1, <30% mortality |
| Control | 1.11 ± 0.0 c | 1.11 ± 0.0 e | 2.2 ± 0.0 e | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Cerón, D.; Rodríguez-Leyva, E.; Lomeli-Flores, J.R.; Soto-Rojas, L.; Ramírez-Alarcón, S.; Segura-Miranda, A. Toxicity and Residual Activity of Insecticides against Diadegma insulare, a Parasitoid of the Diamondback Moth. Insects 2022, 13, 514. https://doi.org/10.3390/insects13060514
Ramírez-Cerón D, Rodríguez-Leyva E, Lomeli-Flores JR, Soto-Rojas L, Ramírez-Alarcón S, Segura-Miranda A. Toxicity and Residual Activity of Insecticides against Diadegma insulare, a Parasitoid of the Diamondback Moth. Insects. 2022; 13(6):514. https://doi.org/10.3390/insects13060514
Chicago/Turabian StyleRamírez-Cerón, Daniel, Esteban Rodríguez-Leyva, J. Refugio Lomeli-Flores, Lauro Soto-Rojas, Samuel Ramírez-Alarcón, and Antonio Segura-Miranda. 2022. "Toxicity and Residual Activity of Insecticides against Diadegma insulare, a Parasitoid of the Diamondback Moth" Insects 13, no. 6: 514. https://doi.org/10.3390/insects13060514