
Towards Predictions of Interaction Dynamics between Cereal Aphids and Their Natural Enemies: A Review


Abstract
:Simple Summary
Abstract
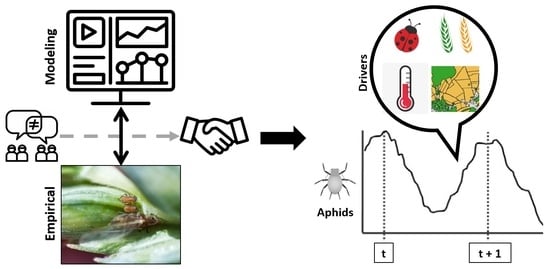
1. Introduction
2. Materials and Methods
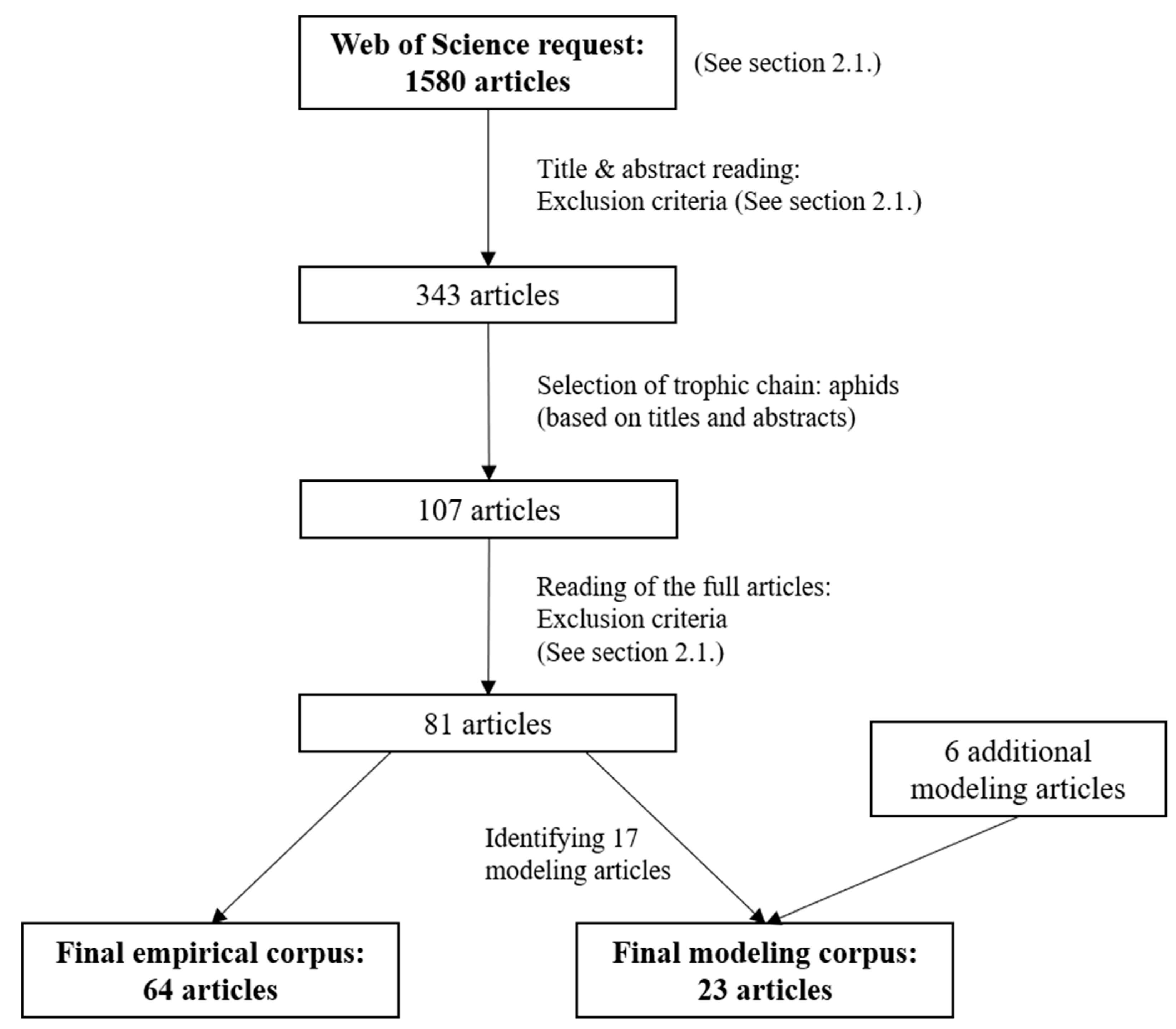
2.1. Literature Search
- the studied organisms (pests and natural enemies) must be invertebrates;
- studies must consider annual field crops in temperate climates, and;
- studies must report temporal and/or spatiotemporal population dynamics (of pests and/or their natural enemies) or pest–natural enemy interaction dynamics (predation, parasitism).
2.2. Analysis and Data Assimilation
3. Results
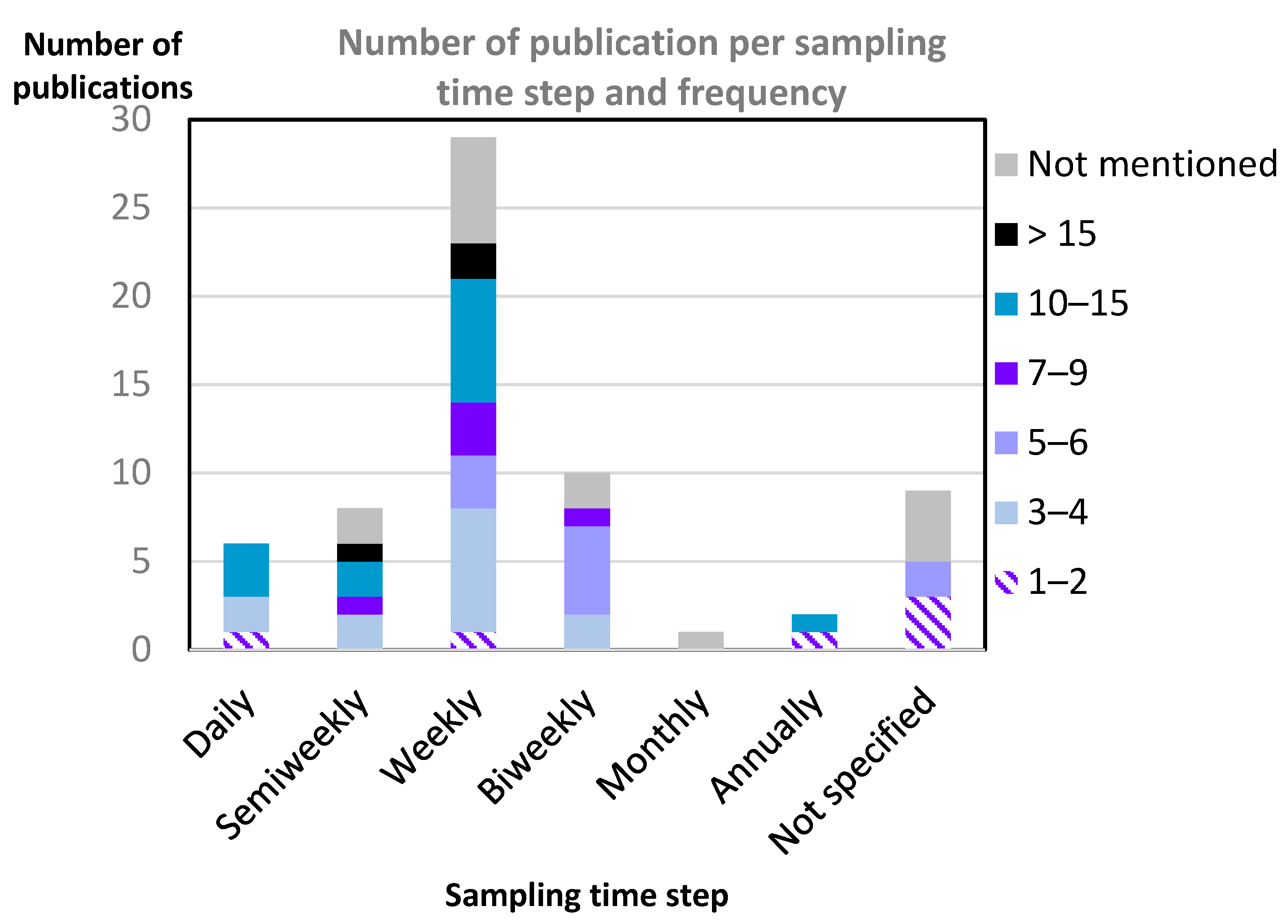
3.1. Overview of Selected Publications
3.2. Relationships between Predictor and Response Indicators
3.2.1. Quantification in Empirical and Modeling Studies
3.2.2. Drivers of Aphid Abundance and Population Growth Rates and Their Natural Enemies Abundance Dynamics
3.2.3. Relative Rankings of Predictor Indicators in Conjoint Analyses
3.3. Ecological Hypotheses
4. Discussion
4.1. The Need for Crossover between Empirical and Modeling Studies
4.2. A Diversity of Models but a Lack of Integration of Key Drivers
4.3. Key Drivers of Aphid Dynamics and Research Issues
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fauvergue, X. Biocontrôle Elements Pour Une Protection Agroecologique des Cultures; Éditions Quae: Versailles, France, 2020. [Google Scholar]
- Begg, G.S.; Cook, S.M.; Dye, R.; Ferrante, M.; Franck, P.; Lavigne, C.; Birch, A.N.E.A. functional overview of conservation biological control. Crop Prot. 2017, 97, 145–158. [Google Scholar] [CrossRef]
- Duru, M.; Therond, O.; Fares, M. Designing agroecological transitions; A review. Agron. Sustain. Dev. 2015, 35, 1237–1257. [Google Scholar] [CrossRef] [Green Version]
- Welch, K.D.; Harwood, J.D. Temporal dynamics of natural enemy-pest interactions in a changing environment. Biol. Control. 2014, 75, 18–27. [Google Scholar] [CrossRef]
- Rusch, A.; Bommarco, R.; Jonsson, M.; Smith, H.G.; Ekbom, B. Flow and stability of natural pest control services depend on complexity and crop rotation at the landscape scale. J. Appl. Ecol. 2013, 50, 345–354. [Google Scholar] [CrossRef]
- Rusch, A.; Chaplin-Kramer, R.; Gardiner, M.M.; Hawro, V.; Holland, J.; Landis, D.; Bommarco, R. Agricultural landscape simplification reduces natural pest control: A quantitative synthesis. Agric. Ecosyst. Environ. 2016, 221, 198–204. [Google Scholar] [CrossRef] [Green Version]
- Schellhorn, N.A.; Gagic, V.; Bommarco, R. Time will tell: Resource continuity bolsters ecosystem services. Trends Ecol. Evol. 2015, 30, 524–530. [Google Scholar] [CrossRef]
- Jeanneret, P.; Aviron, S.; Alignier, A.; Lavigne, C.; Helfenstein, J.; Herzog, F.; Petit, S. Agroecology landscapes. Landsc. Ecol. 2021, 36, 2235–2257. [Google Scholar] [CrossRef]
- Vialatte, A. Ecologie du puceron S. Avenae Dans un Paysage Agricole: Application au Raisonnement de la Lutte Intégrée et Chimique sur Blé au Printemps. Ph.D. Thesis, Agrocampus Ouest, Rennes, France, 2006. [Google Scholar]
- Lundgren, J.G.; Hesler, L.S.; Clay, S.A.; Fausti, S.F. Insect communities in soybeans of eastern South Dakota: The effects of vegetation management and pesticides on soybean aphids, bean leaf beetles, and their natural enemies. Crop Prot. 2013, 43, 104–118. [Google Scholar] [CrossRef] [Green Version]
- Meisner, M.H.; Harmon, J.P.; Ives, A.R. Temperature effects on long-term population dynamics in a parasitoid-host system. Ecol. Monogr. 2014, 84, 457–476. [Google Scholar] [CrossRef]
- Raymond, L.; Ortiz-Martínez, S.A.; Lavandero, B. Temporal variability of aphid biological control in contrasting landscape contexts. Biol. Control. 2015, 90, 148–156. [Google Scholar] [CrossRef]
- Ekbom, B.S.; Wiktelius, S.; Chiverton, P.A. Can polyphagous predators control the bird cherry-oat aphid (Rhopalosiphum padi) in spring cereals? A simulation study. Entomol. Exp. Appl. 1992, 65, 215–223. [Google Scholar] [CrossRef]
- Van Der Werf, W. How Do Immigration Rates Affect Predator/Prey Interactions in Field Crops? In Predictions from Simple Models and an Example Involving the Spread of Aphid-Borne Viruses Sugar Beet; Aarhus University: Aarhus, Denmark, 1995; Volume 70, pp. 295–312. [Google Scholar]
- Kindlmann, P.; Dixon, A.F. Modelling Population Dynamics of Aphids and Their Natural Enemies. In Aphid Biodiversity under Environmental Change; Kindlmann, P., Dixon, A., Michaud, J., Eds.; Springer: Dordrecht, The Netherlands, 2010. [Google Scholar] [CrossRef]
- Chaplin-Kramer, R.; O’Rourke, M.E.; Blitzer, E.J.; Kremen, C.A. meta-analysis of crop pest and natural enemy response to landscape complexity. Ecol. Lett. 2011, 14, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Karp, D.S.; Chaplin-Kramer, R.; Meehan, T.D.; Martin, E.A.; DeClerck, F.; Grab, H.; Zou, Y. Crop pests and predators exhibit inconsistent responses to surrounding landscape composition. Proc. Natl. Acad. Sci. USA 2018, 115, E7863–E7870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratsimba, N.; Therond, O.; Parry, H.; Monteil, C.; Vialatte, A. Inconsistent responses of conservation biocontrol to landscape structure: New insights from a network-based review. Ecol. Appl. 2021, 32, e02456. [Google Scholar] [CrossRef] [PubMed]
- Tixier, P.; Peyrard, N.; Aubertot, J.N.; Gaba, S.; Radoszycki, J.; Caron-Lormier, G.; Sabbadin, R. Modelling Interaction Networks for Enhanced Ecosystem Services in Agroecosystems. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherland, 2013; Volume 49, pp. 437–480. [Google Scholar] [CrossRef]
- Petit, S.; Muneret, L.; Carbonne, B. Landscape-scale. Expansion of Agroecology to Enhance Natural Pest Control: A Systematic Review. In The Future of Agricultural Landscapes, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 63, pp. 1–48. [Google Scholar] [CrossRef]
- Poggi, S.; Le Cointe, R.; Riou, J.B.; Larroudé, P.; Thibord, J.B.; Plantegenest, M. Relative influence of climate and agroenvironmental factors on wireworm damage risk in maize crops. J. Pest Sci. 2018, 91, 585–599. [Google Scholar] [CrossRef]
- Ro, T.H.; Long, G.E. GPA-phenodynamics—A simulation model for the population dynamics and phenology of green peach aphid in potato: Formulation, validation, and analysis. Ecol. Model. 1999, 119, 197–209. [Google Scholar] [CrossRef]
- Hallett, R.H.; Bahlai, C.A.; Xue, Y.; Schaafsma, A.W. Incorporating. natural enemy units into a dynamic action threshold for the soybean aphid, Aphis glycines (Homoptera: Aphididae). Pest Manag. Sci. 2014, 70, 879–888. [Google Scholar] [CrossRef]
- Nault, L.R. Arthropod Transmission of plant viruses: A new synthesis. Ann. Entomol. Soc. Am. 1997, 90, 521–541. [Google Scholar] [CrossRef]
- Katis, N.I.; Tsitsipis, J.A.; Stevens, M.; Powell, G. Transmission of Plant Viruses. In Aphids as Crop Pests; van Emden, H.F., Harrington, R., Eds.; CABI: London, UK, 2007; pp. 353–377. [Google Scholar]
- Dedryver, C.; Ralec, A.L. The conflicting relationships between aphids and men: A review of aphid damage and control strategies. Comptes. Rendus Biol. 2010, 333, 539–553. [Google Scholar] [CrossRef]
- Blackman, R.L.; Eastop, V.F. Taxonomic Issue. In Aphids as Crop Pests, 2nd ed.; van Emden, H.F., Harrington, R., Eds.; CABI International: Wallingford, UK; pp. 1–29.
- Hill, D.S. Agricultural Insect Pests of Temperate Regions and Their Control; Cambridge University Press: Cambridge, UK, 1987; p. 659. [Google Scholar] [CrossRef]
- Brodeur, J.; Hajek, A.E.; Heimpel, G.E.; Sloggett, J.J.; Mackauer, M.; Pell, J.J.; Volkl, W. Predators, parasitoids and pathogens. In Aphids as Crop Pests, 2nd ed.; van Emden, H.F., Harrington, R., Eds.; CABI International: Wallingford, UK; pp. 225–261.
- Alexandridis, N.; Marion, G.; Chaplin-Kramer, R.; Dainese, M.; Ekroos, J.; Grab, H.; Clough, Y. Models of natural pest control: Towards predictions across agricultural landscapes. Biol. Control. 2021, 163, 104761. [Google Scholar] [CrossRef]
- Fidelis, E.G.; Farias, E.S.; Lopes, M.C.; Sousa, F.F.; Zanuncio, J.C.; Picanço, M.C. Contributions of climate, plant phenology and natural enemies to the seasonal variation of aphids on cabbage. J. Appl. Entomol. 2019, 143, 365–370. [Google Scholar] [CrossRef]
- Kataria, R.; Kumar, D. Population dynamics of Aphis craccivora (Koch) and its natural enemies on bean crop in relation to weather parameters in Vadodara, Gujarat, India. Legume Res. 2017, 40, 571–579. [Google Scholar] [CrossRef] [Green Version]
- Costamagna, A.C.; Landis, D.A. Lack of strong refuges allows top-down control of soybean aphid by generalist natural enemies. Biol. Control. 2011, 57, 184–192. [Google Scholar] [CrossRef]
- Dahlin, I.; Rubene, D.; Glinwood, R.; Ninkovic, V. Pest suppression in cultivar mixtures is influenced by neighbor-specific plant–plant communication. Ecol. Appl. 2018, 28, 2187–2196. [Google Scholar] [CrossRef] [Green Version]
- Whitney, K.S.; Meehan, T.D.; Kucharik, C.J.; Zhu, J.; Whitney, K.S.; Meehan, T.D.; Gratton, C. Explicit modeling of abiotic and landscape factors reveals precipitation and forests associated with aphid abundance. Ecol. Appl. 2016, 26, 2600–2610. [Google Scholar] [CrossRef]
- Costamagna, A.C.; Venables, W.N.; Schellhorn, N.A. Landscape-scale pest suppression is mediated by timing of predator arrival. Ecol. Appl. 2015, 25, 1114–1130. [Google Scholar] [CrossRef]
- Alhmedi, A.; Haubruge, E.; Francis, F. Effect of stinging nettle habitats on aphidophagous predators and parasitoids in wheat and green pea fields with special attention to the invader Harmonia axyridis Pallas (Coleoptera: Coccinellidae). Entomol. Sci. 2009, 12, 349–358. [Google Scholar] [CrossRef]
- Zhao, Z.; Reddy, G.V.P. Semi-natural habitats mediate influence of inter-annual landscape variation on cereal aphid-parasitic wasp system in an agricultural landscape. Biol. Control. 2019, 128, 17–23. [Google Scholar] [CrossRef]
- Elliott, N.; Kieckhefer, R.W. Response by coccinellids to spatial variation in cereal aphid density. Popul. Ecol. 2000, 42, 81–90. [Google Scholar] [CrossRef]
- Costamagna, A.C.; Landis, D.A. Predators exert top-down control of soybean aphid across a gradient of agricultural management systems. Ecol. Appl. 2006, 16, 1619–1628. [Google Scholar] [CrossRef] [Green Version]
- Bommarco, R.; Wetterlind, S.; Sigvald, R. Cereal aphid populations in non-crop habitats show strong density dependence. J. Appl. Ecol. 2007, 44, 1013–1022. [Google Scholar] [CrossRef]
- Chen, K.; Hopper, K.R. Diuraphis noxia (Homoptera: Aphididae) Population Dynamics and Impact of Natural Enemies in the Montpellier Region of Southern France. Environ. Entomol. 1997, 26, 866–875. [Google Scholar] [CrossRef]
- Chaplin-Kramer, R.; Kremen, C. Pest control experiments show benefits of complexity at landscape and local scales. Ecol. Appl. 2012, 22, 1936–1948. [Google Scholar] [CrossRef] [PubMed]
- Hesler, L.S. Inventory and assessment of foliar natural enemies of the soybean aphid (Hemiptera: Aphididae) in South Dakota. Environ. Entomol. 2014, 43, 577–588. [Google Scholar] [CrossRef] [Green Version]
- Rhainds, M.; Yoo, H.J.; Kindlmann, P.; Voegtlin, D.; Castillo, D.; Rutledge, C.; O’Neil, R.J. Two-year oscillation cycle in abundance of soybean aphid in Indiana. Agric. For. Entomol. 2010, 12, 251–257. [Google Scholar] [CrossRef]
- Evans, E.W. Indirect interactions in biological control of insects: Pests and natural enemies in alfalfa. Ecol. Appl. 1996, 6, 920–930. [Google Scholar] [CrossRef]
- Yoo, H.J.S.; O’Neil, R.J. Temporal relationships between the generalist predator, Orius insidiosus, and its two major prey in soybean. Biol. Control. 2009, 48, 168–180. [Google Scholar] [CrossRef]
- Winder, L.; Alexander, C.J.; Holland, J.M.; Symondson, W.O.C.; Perry, J.N.; Woolley, C. Predatory activity and spatial pattern: The response of generalist carabids to their aphid prey. J. Anim. Ecol. 2005, 74, 443–454. [Google Scholar] [CrossRef]
- Karley, A.J.; Pitchford, J.W.; Douglas, A.E.; Parker, W.E.; Howardh, J.J. The causes and processes of the mid-summer population crash of the potato aphids Macrosiphum euphorbiae and Myzus persicae (Hemiptera: Aphididae). Bull. Entomol. Res. 2003, 93, 425–438. [Google Scholar] [CrossRef]
- Bortolotto, O.C.; De Oliveira Menezes Júnior, A.; Hoshino, A.T. Aphidophagous parasitoids can forage wheat crops before aphid infestation, Parana State, Brazil. J. Insect Sci. 2015, 15, 40. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, A.; Brodeur, J. Estimating parasitoid impact on aphid populations in the field. Biol. Control. 2018, 119, 33–42. [Google Scholar] [CrossRef]
- Rutledge, C.E.; O’Neil, R.J. Orius insidiosus (Say) as a predator of the soybean aphid, Aphis glycines Matsumura. Biol. Control. 2005, 33, 56–64. [Google Scholar] [CrossRef]
- Donatelli, M.; Magarey, R.D.; Bregaglio, S.; Willocquet, L.; Whish, J.P.M.; Savary, S. Modelling the impacts of pests and diseases on agricultural systems. Agric. Syst. 2017, 155, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Lasso, E.; Motisi, N.; Avelino, J.; Corrales, J.C. FramePests: A comprehensive framework for crop pests modeling and forecasting. IEEE Access 2021, 9, 115579–115598. [Google Scholar] [CrossRef]
- Malard, J.J.; Adamowski, J.F.; Rojas Díaz, M.; Nassar, J.B.; Anandaraja, N.; Tuy, H.; Melgar-Quiñonez, H.R. Development of a new modular software tool for agroecological food web model development: Tiko’n. Eur. J. Agron. 2020, 116, 126033. [Google Scholar] [CrossRef]
- Bahlai, C.A.; Weiss, R.M.; Hallett, R.H. A mechanistic model for a tritrophic interaction involving soybean aphid, Its host plants, And multiple natural enemies. Ecol. Model. 2013, 254, 54–70. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Van Der Werf, W. The Effect of the Area and Configuration of Hibernation Sites on the Control of Aphids by Coccinella septempunctata (Coleoptera: Coccinellidae) in Agricultural Landscapes: A Simulation Study. Environ. Entomol. 2003, 32, 1290–1304. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, F.J.J.A.; Honěk, A.; Van Der Werf, W. Changes in agricultural land use can explain population decline in a ladybeetle species in the Czech Republic: Evidence from a process-based spatially explicit model. Landsc. Ecol. 2007, 22, 1541–1554. [Google Scholar] [CrossRef]
- Plantegenest, M.; Pierre, J.S.; Dedryver, C.A.; Kindlmann, P. Assessment of the relative impact of different natural enemies on population dynamics of the grain aphid Sitobion avenae in the field. Ecol. Entomol. 2001, 26, 404–410. [Google Scholar] [CrossRef] [Green Version]
- Curtsdotter, A.; Banks, H.T.; Banks, J.E.; Jonsson, M.; Jonsson, T.; Laubmeier, A.N.; Bommarco, R. Ecosystem function in predator–prey food webs—Confronting dynamic models with empirical data. J. Anim. Ecol. 2019, 88, 196–210. [Google Scholar] [CrossRef] [Green Version]
- Plantegenest, M.; Pierre, J.S.; Caillaud, C.; Simon, J.C.; Dedryver, C.A.; Cluzeau, S. A model to describe the reproductvity rate in the aphid Sitobion avenae—A case study. Eur. J. Entomol. 1996, 93, 545–553. [Google Scholar]
- Miksanek, J.R.; Heimpel, G.E. A matrix model describing host-parasitoid population dynamics: The case of Aphelinus certus and soybean aphid. PLoS ONE 2019, 14, e0218217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preedy, K.F.; Chaplain, M.A.J.; Leybourne, D.J.; Marion, G.; Karley, A.J. Learning-induced switching costs in a parasitoid can maintain diversity of host aphid phenotypes although biocontrol is destabilized under abiotic stress. J. Anim. Ecol. 2020, 89, 1216–1229. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; O’Neil, R.J. Potential of an alternative prey to disrupt predation of the generalist predator, Orius insidiosus, on the pest aphid, Aphis glycines, via short-term indirect interactions. Bull. Entomol. Res. 2008, 98, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Jacometti, M.; Jørgensen, N.; Wratten, S. Enhancing biological control by an omnivorous lacewing: Floral resources reduce aphid numbers at low Aphid densities. Biol. Control. 2010, 55, 159–165. [Google Scholar] [CrossRef]
- Jonsson, M.; Bommarco, R.; Ekbom, B.; Smith, H.G.; Bengtsson, J.; Caballero-Lopez, B.; Olsson, O. Ecological production functions for biological control services in agricultural landscapes. Methods Ecol. Evol. 2014, 5, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Martínez, S.; Staudacher, K.; Baumgartner, V.; Traugott, M.; Lavandero, B. Intraguild predation is independent of landscape context and does not affect the temporal dynamics of aphids in cereal fields. J. Pest Sci. 2020, 93, 235–249. [Google Scholar] [CrossRef]
- Roubinet, E.; Jonsson, T.; Malsher, G.; Staudacher, K.; Traugott, M.; Ekbom, B.; Jonsson, M. High redundancy as well as complementary prey choice characterize generalist predator food webs in agroecosystems. Sci. Rep. 2018, 8, 8054. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, J.R.; Myers, S.W.; Gratton, C. Density-dependent responses of soybean aphid (Aphis glycines Matsumura) populations to generalist predators in mid to late season soybean fields. Biol. Control. 2007, 43, 111–118. [Google Scholar] [CrossRef]
- Jonsson, M.; Sigvald, R. Suction-trap catches partially predict infestations of the grain aphid Sitobion avenae in winter wheat fields. J. Appl. Entomol. 2016, 140, 553–557. [Google Scholar] [CrossRef] [Green Version]
- Letourneau, D.K.; Armbrecht, I.; Rivera, B.S.; Lerma, J.; Carmona, E.J.; Daza, M.C.; Trujillo, A.R. Does plant diversity benefit agroecosystems? A synthetic review. Ecol. Appl. 2011, 21, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.B.; Chen, J.; Lian, L.Y.; Francis, F.; Haubruge, E.; Bragard, C.; Cheng, D. Influence of Garlic Intercropping or Active Emitted Volatiles in Releasers on Aphid and Related Beneficial in Wheat Fields in China. J. Integr. Agric. 2013, 12, 467–473. [Google Scholar] [CrossRef]
- Amini, B.; Madadi, H.; Desneux, N.; Lotfalizadeh, H.A. Impact of irrigation systems on seasonal occurrence of Brevicoryne brassicae and its parasitism by Diaeretiella rapae on canola. J. Entomol. Res. Soc. 2012, 14, 15–26. [Google Scholar]
- Thies, C.; Roschewitz, I.; Tscharntke, T. The landscape context of cereal aphid-parasitoid interactions. Proc. R. Soc. B Biol. Sci. 2005, 272, 203–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, J.M.; Douma, J.C.; Crowley, L.; James, L.; Kor, L.; Stevenson, D.R.W.; Smith, B.M. Semi-natural habitats support biological control, pollination and soil conservation in Europe. A review. Agron. Sustain. Dev. 2017, 37, 31. [Google Scholar] [CrossRef] [Green Version]
- Alignier, A.; Raymond, L.; Deconchat, M.; Menozzi, P.; Monteil, C.; Sarthou, J.P.; Ouin, A. The effect of semi-natural habitats on aphids and their natural enemies across spatial and temporal scales. Biol. Control 2014, 77, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Tscharntke, T.; Karp, D.S.; Chaplin-Kramer, R.; Batáry, P.; DeClerck, F.; Gratton, C.; Zhang, W. When natural habitat fails to enhance biological pest control—Five hypotheses. Biol. Conserv. 2016, 204, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Gilabert, A.; Gauffre, B.; Parisey, N.; Le Gallic, J.F.; Lhomme, P.; Bretagnolle, V.; Plantegenest, M. Influence of the surrounding landscape on the colonization rate of cereal aphids and phytovirus transmission in autumn. J. Pest Sci. 2017, 90, 447–457. [Google Scholar] [CrossRef]
- Holland, J.M.; Jeanneret, P.; Moonen, A.C.; Van Der Werf, W.; Rossing, W.A.H.; Antichi, D.; Veromann, E. Correction: Approaches to identify the value of seminatural habitats for conservation biological control. Insects 2020, 11, 195. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Desneux, N.; Lu, Y.; Wu, K. Key aphid natural enemies showing positive effects on wheat yield through biocontrol services in northern China. Agric. Ecosyst. Environ. 2018, 266, 1–9. [Google Scholar] [CrossRef]
- Arshad, M.; Ahmad, S.; Sufyan, M.; Abdin, Z.; Maqsood, S. Population dynamics of aphids and their natural enemies associated with strip-intercropping in wheat crop. Pak. J. Zool. 2018, 50, 1225–1230. [Google Scholar] [CrossRef]
- Bahlai, C.A.; Werf WVander O’Neal, M.; Hemerik, L.; Landis, D.A. Shifts in dynamic regime of an invasive lady beetle are linked to the invasion and insecticidal management of its prey. Ecol. Appl. 2015, 25, 1807–1818. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Weis, J.J.; Forbes, A.E.; Tilmon, K.J.; Ives, A.R. Biodiversity as both a cause and consequence of resource availability: A study of reciprocal causality in a predator-prey system. J. Anim. Ecol. 2006, 75, 497–505. [Google Scholar] [CrossRef] [Green Version]
- Costamagna, A.C.; Van Der Werf, W.; Bianchi, F.J.J.A.; Landis, D.A. An exponential growth model with decreasing r captures bottom-up effects on the population growth of Aphis glycines Matsumura (Hemiptera: Aphididae). Agric. For. Entomol. 2007, 9, 297–305. [Google Scholar] [CrossRef]
- Dai, H.; Wang, Y.; Du, Y.; Ding, J. Effects of plant trichomes on herbivores and predators on soybeans. Insect Sci. 2010, 17, 406–413. [Google Scholar] [CrossRef]
- Díaz, B.M.; Legarrea, S.; Marcos-García M, Á.; Fereres, A. The spatio-temporal relationships among aphids, the entomophthoran fungus, Pandora neoaphidis, and aphidophagous hoverflies in outdoor lettuce. Biol. Control. 2010, 53, 304–311. [Google Scholar] [CrossRef]
- Duffy, C.; Fealy, R.; Fealy, R.M. An improved simulation model to describe the temperature-dependent population dynamics of the grain aphid, Sitobion avenae. Ecol. Modell. 2017, 354, 140–171. [Google Scholar] [CrossRef] [Green Version]
- Evans, E.W.; Toler, T.R. Aggregation of polyphagous predators in response to multiple prey: Ladybirds (Coleoptera: Coccinellidae) foraging in alfalfa. Popul. Ecol. 2007, 49, 29–36. [Google Scholar] [CrossRef]
- Fox, T.B.; Landis, D.A.; Cardoso, F.F.; Difonzo, C.D. Impact of predation on establishment of the soybean aphid, Aphis glycines in soybean, Glycine max. BioControl 2005, 50, 545–563. [Google Scholar] [CrossRef]
- Gagic, V.; Hänke, S.; Thies, C.; Scherber, C.; Tomanović, Ž; Tscharntke, T. Agricultural intensification and cereal aphid-parasitoid-hyperparasitoid food webs: Network complexity, temporal variability and parasitism rates. In Oecologia; 2012; Volume 170, pp. 1099–1109. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, M.M.; Landis, D.A. Impact of intraguild predation by adult Harmonia axyridis (Coleoptera: Coccinellidae) on Aphis glycines (Hemiptera: Aphididae) biological control in cage studies. Biol. Control. 2007, 40, 386–395. [Google Scholar] [CrossRef]
- Gebauer, K.; Hemerik, L.; Meyhöfer, R. Effects of climate change on pest-parasitoid dynamics: Development of a simulation model and first results. J. Plant Dis. Prot. 2015, 122, 28–35. [Google Scholar] [CrossRef]
- Ghahramani, M.; Karimzadeh, R.; Iranipour, S.; Sciarretta, A. Does harvesting affect the spatio-temporal signature of pests and natural enemies in alfalfa fields? Agronomy 2019, 9, 532. [Google Scholar] [CrossRef] [Green Version]
- Gross, K.; Ives, A.R.; Nordheim, E.V. Estimating fluctuating vital rates from time-series data: A case study of aphid biocontrol. Ecology 2005, 86, 740–752. [Google Scholar] [CrossRef]
- Holland, J.M.; Winder, L.; Woolley, C.; Alexander, C.J.; Perry, J.N. The spatial dynamics of crop and ground active predatory arthropods and their aphid prey in winter wheat. Bull. Entomol. Res. 2004, 94, 419–431. [Google Scholar] [CrossRef]
- Imbert, C.; Papaïx, J.; Husson, L.; Warlop, F.; Lavigne, C. Estimating population dynamics parameters of cabbage pests in temperate mixed apple tree-cabbage plots compared to control vegetable plots. Crop Prot. 2020, 129, 105037. [Google Scholar] [CrossRef]
- Khodeir, I.A.; Khattab, M.A.; Rakha, O.M.; Sharabash, A.S.; Ueno, T.; Mousa, K.M. Population densities of pest aphids and their associated natural enemies on faba bean in Kafr el–Sheikh, Egypt. J. Fac. Agric. 2020, 65, 97–102. [Google Scholar] [CrossRef]
- Kindlmann, P.; Houdková, K. Intraguild predation: Fiction or reality? Popul. Ecol. 2006, 48, 317–322. [Google Scholar] [CrossRef]
- Liu, J.; Yan, Y.; Ali, A.; Wang, N.; Zhao, Z.; Yu, M. Effects of wheat-maize intercropping on population dynamics of wheat aphids and their natural enemies. Sustainability 2017, 9, 1390. [Google Scholar] [CrossRef] [Green Version]
- Merrill, S.C.; Peairs, F.B. Quantifying Russian Wheat Aphid Pest Intensity Across the Great Plains. Environ. Entomol. 2013, 41, 1505–1515. [Google Scholar] [CrossRef]
- Miao, J.; Wu, K.; Hopper, K.R.; Li, G. Population dynamics of Aphis glycines (Homoptera: Aphididae) and impact of natural enemies in Northern China. Environ. Entomol. 2007, 36, 840–848. [Google Scholar] [CrossRef]
- Newman, J.A. Climate change and the fate of cereal aphids in Southern Britain. Glob. Chang. Biol. 2005, 11, 940–944. [Google Scholar] [CrossRef]
- Newman, J.A.; Gibson, D.J.; Parsons, A.J.; Thornley JH, M. How predictable are aphid population responses to elevated CO2? J. Anim. Ecol. 2003, 72, 556–566. [Google Scholar] [CrossRef]
- Newman, J.A. Climate change and cereal aphids: The relative effects of increasing CO2 and temperature on aphid population dynamics. Glob. Chang. Biol. 2004, 10, 5–15. [Google Scholar] [CrossRef]
- Newman, J.A. Using the output from global circulation models to predict changes in the distribution and abundance of cereal aphids in Canada: A mechanistic modeling approach. Glob. Chang. Biol. 2006, 12, 1634–1642. [Google Scholar] [CrossRef]
- Ong, T.W.Y.; Allen, D.; Vandermeer, J. Huffaker revisited: Spatial heterogeneity and the coupling of ineffective agents in biological control. Ecosphere 2018, 9, e02299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz-Martínez, S.A.; Lavandero, B. The effect of landscape context on the biological control of Sitobion avenae: Temporal partitioning response of natural enemy guilds. J. Pest Sci. 2018, 91, 41–53. [Google Scholar] [CrossRef]
- Paulson, M.D.; Houston, A.I.; McNamara, J.M.; Payne RJ, H. Seasonal dispersal of pests: One surge or two? J. Evol. Biol. 2009, 22, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rodríguez, J.; Shortall, C.R.; Bell, J.R. Large-scale migration synchrony between parasitoids and their host. Ecol. Entomol. 2015, 40, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Piyaratne MK, D.K.; Zhao, H.; Meng, Q. APHIDSim: A population dynamics model for wheat aphids based on swallowtail catastrophe theory. Ecol. Modell. 2013, 253, 9–16. [Google Scholar] [CrossRef]
- Seiter, N.J.; Miskelley, A.D.; Lorenz, G.M.; Joshi, N.K.; Studebaker, G.E.; Kelley, J.P. Impact of Planting Date on Melanaphis sacchari (Hemiptera: Aphididae) Population Dynamics and Grain Sorghum Yield. J. Econ. Entomol. 2019, 112, 2731–2736. [Google Scholar] [CrossRef]
- Smits, N.; Dupraz, C.; Dufour, L. Unexpected lack of influence of tree rows on the dynamics of wheat aphids and their natural enemies in a temperate agroforestry system. Agrofor. Syst. 2012, 85, 153–164. [Google Scholar] [CrossRef]
- Tenhumberg, B.; Poehling, H. Syrphids as natural enemies of cereal aphids in Germany: Aspects of their biology and efficacy in different years and regions. Agric. Ecosyst. Environ. 1995, 52, 39–43. [Google Scholar] [CrossRef] [Green Version]
- Tran, D.H.; Khac PLe Thi, T.; Dong, H.; Ueno, T. Control efficacy of pongam (Pongamia pinnata L.) leaf extract against the turnip aphid Lipaphis pseudobrassicae (Davis) (Hemiptera: Aphididae). J. Fac. Agr. Kyushu Univ. 2016, 61, 141–145. [Google Scholar] [CrossRef]
- Vollhardt IM, G.; Bianchi FJ, J.A.; Wäckers, F.L.; Thies, C.; Tscharntke, T. Spatial distribution of flower vs. honeydew resources in cereal fields may affect aphid parasitism. Biol. Control 2010, 53, 204–213. [Google Scholar] [CrossRef]
- Wang, H.H.; Grant, W.E.; Elliott, N.C.; Brewer, M.J.; Koralewski, T.E.; Westbrook, J.K.; Sword, G.A. Integrated modelling of the life cycle and aeroecology of wind-borne pests in temporally-variable spatially-heterogeneous environment. Ecol. Modell. 2019, 399, 23–38. [Google Scholar] [CrossRef]
- Wang, L.; Hui, C.; Sandhu, H.S.; Li, Z.; Zhao, Z. Population dynamics and associated factors of cereal aphids and armyworms under global change. Sci. Rep. 2015, 5, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Piyaratne MK, D.K.; Zhao, H.; Li, C.; Hu, Z.; Hu, X. Butterfly catastrophe model for wheat aphid population dynamics: Construction, analysis and application. Ecol. Modell. 2014, 288, 55–61. [Google Scholar] [CrossRef]
- Zhao, Z.H.; Hui, C.; Li, Z.H.; Li, B.L. Habitat heterogeneity stabilizes the spatial and temporal interactions between cereal aphids and parasitic wasps. Basic Appl. Ecol. 2015, 16, 510–518. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, L.; Chen, J.; Francis, F.; Haubruge, E.; Liu, Y.; Cheng, D. Adaptation of wheat-pea intercropping pattern in china to reduce Sitobion avenae (Hemiptera: Aphididae) occurrence by promoting natural enemies. Agroecol. Sustain. Food Syst. 2013, 37, 1001–1016. [Google Scholar] [CrossRef]
- Zumoffen, L.; Salto, C.; Salvo, A. Preliminary study on parasitism of aphids (Hemiptera: Aphididae) in relation to characteristics of alfalfa fields (Medicago sativa L.) in the Argentine Pampas. Agric. Ecosyst. Environ. 2012, 159, 49–54. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Natural Enemies | Aphids | Crop Outcomes | Total | |||
|---|---|---|---|---|---|---|
| Response Indicators | Empirical | Modeling | Empirical | Modeling | Empirical | |
| Abundance | 54 | 5 | 52 | 9 | - | 120 |
| Population growth rate | 7 | 6 | 44 | 9 | - | 66 |
| Migration/flux | 1 | - | 8 | 3 | - | 12 |
| Parasitism | - | - | 10 | 2 | - | 12 |
| Predation | - | - | 8 | 3 | - | 11 |
| Community diversity | 7 | - | 2 | - | - | 9 |
| Agricultural results * | - | - | 2 | - | 6 | 8 |
| Pest suppression | - | 2 | 4 | - | - | 6 |
| Spatiotemporal stability | 3 | - | 2 | - | - | 5 |
| Intraguild predation | - | - | - | 2 | - | 2 |
| Biocontrol | - | - | - | - | 1 | 1 |
| Total | 74 | 13 | 132 | 26 | 7 | 252 |
| Response Variable | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aphids Abundance Dynamics | Aphids Growth Rate Dynamics | Enemy Abundance Dynamics | TOT | ||||||||||||||||
| Predictor Variable | Pos | NS | Neg | Pos | NS | Neg | Pos | NS | Neg | ||||||||||
| E | M | E | M | E | M | E | M | E | M | E | M | E | M | E | M | E | M | ||
| Insecticide use | - | - | - | - | 2 | - | - | - | - | - | 4 | - | - | - | 1 | - | 2 | - | 9 |
| Fertilizer use | - | - | - | - | - | 1 | - | - | - | - | - | 1 | - | - | - | - | - | 1 | 3 |
| Tillage | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | 1 | - | 1 | - | 3 |
| Sowing date | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Insecticide delay | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | 1 |
| Temperature | 1 | - | - | - | 1 | 1 | 6 | 5 | 3 | - | - | - | - | - | 1 | - | - | - | 18 |
| Precipitation | 1 | - | 1 | - | 1 | 1 | - | - | 2 | - | - | - | 1 | - | - | - | - | - | 7 |
| Humidity | - | - | - | - | 2 | - | - | - | 1 | - | - | - | 3 | - | - | - | - | - | 6 |
| Atmospheric CO2 | - | 1 | - | - | - | 2 | - | 1 | - | - | - | 1 | - | - | - | - | - | - | 5 |
| Wind speed | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | 1 |
| % intercropping | 1 | - | - | - | 4 | - | - | - | - | - | 1 | - | 4 | - | - | - | 2 | - | 12 |
| Intensification | 1 | - | 1 | - | - | - | - | - | 1 | - | - | - | 2 | - | - | - | 1 | - | 6 |
| Crop type | 1 | 1 | - | - | 1 | - | - | - | - | - | 1 | - | - | - | 1 | - | - | - | 5 |
| Agroforestry | - | - | 2 | - | - | - | 1 | - | - | - | - | - | - | - | 1 | - | - | - | 4 |
| % natural borders | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Irrigation | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Aphid abundance | 2 | - | - | - | - | - | - | - | 2 | - | 3 | - | 9 | 1 | 6 | - | - | - | 23 |
| Enemy abundance | - | - | 2 | - | 6 | 1 | - | - | 1 | - | 10 | - | - | - | - | - | - | - | 20 |
| Alternative resources | - | - | - | - | 2 | - | 1 | - | - | - | - | - | 2 | - | 2 | - | 1 | - | 8 |
| Enemy diversity | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Predation | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Migration / flux | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | 1 |
| Parasitism | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | 1 |
| Aphid growth rate | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 1 |
| Landscape complexity | 1 | - | 3 | - | - | - | - | - | - | - | - | - | 1 | - | 3 | - | 2 | - | 10 |
| % SNH | 2 | - | - | - | 2 | - | - | - | - | - | - | - | 2 | 1 | - | - | - | - | 7 |
| % grassland | 1 | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| % crop | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 1 |
| SNH proximity | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | 1 |
| Timing in season | 1 | - | - | - | - | - | 1 | - | - | - | - | - | 1 | - | - | - | - | - | 3 |
| Plant stage | 2 | - | - | - | 1 | - | 1 | - | 1 | - | 2 | - | 1 | - | - | - | - | - | 8 |
| Plant morphology | - | - | 1 | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | 2 |
| TOTAL = | 61 | 53 | 59 | 173 | |||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stell, E.; Meiss, H.; Lasserre-Joulin, F.; Therond, O. Towards Predictions of Interaction Dynamics between Cereal Aphids and Their Natural Enemies: A Review. Insects 2022, 13, 479. https://doi.org/10.3390/insects13050479
Stell E, Meiss H, Lasserre-Joulin F, Therond O. Towards Predictions of Interaction Dynamics between Cereal Aphids and Their Natural Enemies: A Review. Insects. 2022; 13(5):479. https://doi.org/10.3390/insects13050479
Chicago/Turabian StyleStell, Eric, Helmut Meiss, Françoise Lasserre-Joulin, and Olivier Therond. 2022. "Towards Predictions of Interaction Dynamics between Cereal Aphids and Their Natural Enemies: A Review" Insects 13, no. 5: 479. https://doi.org/10.3390/insects13050479