
Genotype-by-Diet Interactions for Larval Performance and Body Composition Traits in the Black Soldier Fly, Hermetia illucens

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Genetic Characterisation of Experimental BSF Strains
2.2. Rearing of Experimental BSF Strains
2.3. Experimental Diets
2.4. Experimental Procedures
2.5. Assessment of Larval Development and Sample Processing
2.6. Compositional Analyses and Calculations
2.7. Statistical Evaluations
3. Results
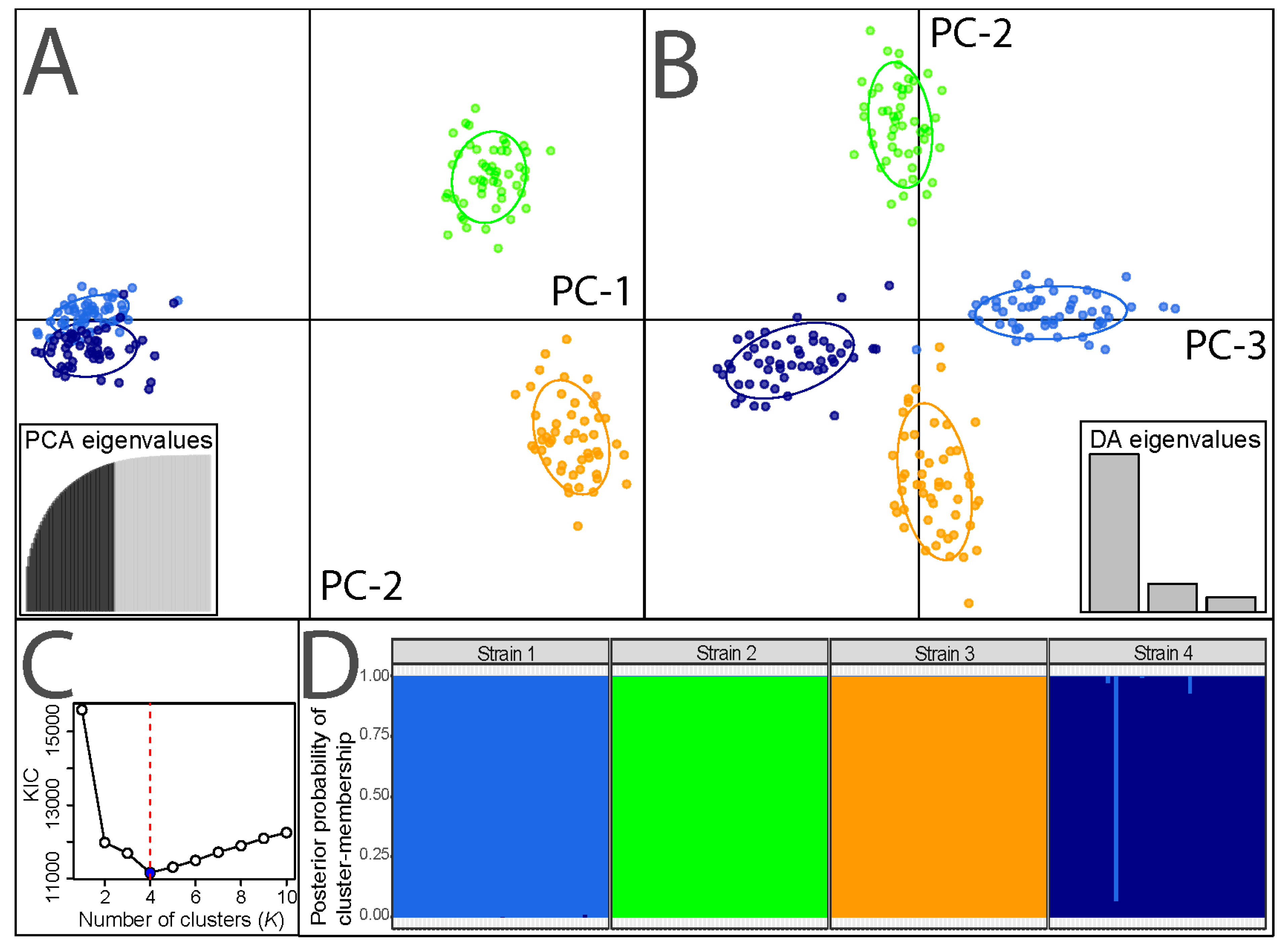
3.1. Population Genetic Characteristics of Experimental BSF Strains
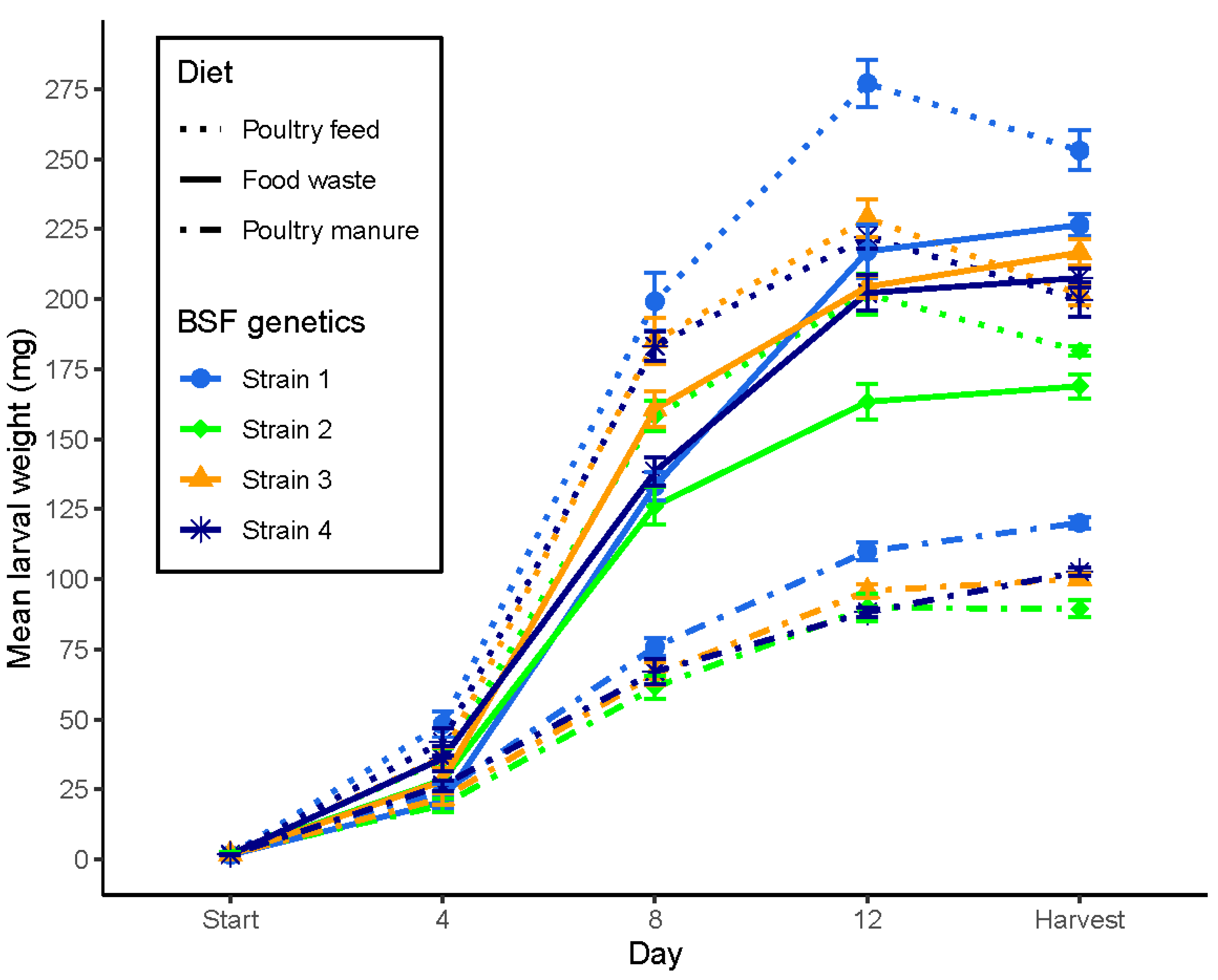
3.2. Larval Development According to Genetic Background and Diet
3.3. Body Composition of Harvested Larvae: Concentrations and Total Amounts of Nutrients
3.4. Larval Bioconversion Efficiency, Dietary Substrate Reduction Rates, and Systemic Losses
3.5. Amino Acid Profiles, Specific Protein Contents, and Nitrogen-to-Protein Conversion
4. Discussion
4.1. G × E Effects on Larval Fattening Performance and Body Composition Traits
4.2. G × E Effects on Bioconversion Efficiency and Sustainability Potential
4.3. G × E Effects in the Context of BSF Developmental Biology and Fitness
4.4. Implications for BSF Genetic Management and Breeding
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Van Huis, A. Insects as food and feed, a new emerging agricultural sector: A review. J. Ins. Food Feed 2020, 6, 27–44. [Google Scholar] [CrossRef] [Green Version]
- Gasco, L.; Biasato, I.; Dabbou, S.; Schiavone, A.; Gai, F. Animals fed insect-based diets: State-of-the-art on digestibility, performance and product quality. Animals 2019, 9, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makkar, H.P.S.; Tran, G.; Henze, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Tech. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Bosch, G.; van Zanten, H.H.E.; Zamprogna, A.; Veenenbos, M.; Meijer, N.P.; van der Fels-Klerx, H.J.; van Loon, J.J.A. Conversion of organic resources by black soldier fly larvae: Legislation, efficiency and environmental impact. J. Clean. Prod. 2019, 222, 355–363. [Google Scholar] [CrossRef]
- Surendra, K.C.; Tomberlin, J.K.; van Huis, A.; Cammack, J.A.; Heckmann, L.-H.L.; Khanal, S.K. Rethinking organic wastes bioconversion: Evaluating the potential of the black soldier fly (Hermetia illucens (L.)) (Diptera: Stratiomyidae) (BSF). Waste Manag. 2020, 117, 58–80. [Google Scholar] [CrossRef]
- Smetana, S.; Spykman, R.; Heinz, V. Environmental aspects of insect mass production. J. Ins. Food Feed 2021, 7, 553–571. [Google Scholar] [CrossRef]
- Čičková, H.; Newton, G.L.; Lacy, R.C.; Kozánek, M. The use of fly larvae for organic waste treatment. Waste Manag. 2015, 35, 68–80. [Google Scholar] [CrossRef]
- Jagtap, S.; Garcia-Garcia, G.; Duong, L.; Swainson, M.; Martindale, W. Codesign of food system and circular economy approaches for the development of livestock feeds from insect larvae. Foods 2021, 10, 1701. [Google Scholar] [CrossRef]
- Elhag, O.; Zhang, Y.; Xiao, X.; Cai, M.; Zheng, L.; Jordan, H.R.; Tomberlin, J.K.; Huang, F.; Yu, Z.; Zhang, J. Inhibition of zoonotic pathogens naturally found in pig manure by black soldier fly larvae and their intestine bacteria. Insects 2022, 13, 66. [Google Scholar] [CrossRef]
- Sogari, G.; Amato, M.; Biasato, I.; Chiesa, S.; Gasco, L. The potential role of insects as feed: A multi-perspective review. Animals 2019, 9, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewusie, E.A.; Kwapong, P.K.; Ofosu-Budu, G.; Sandrock, C.; Akumah, A.; Nartey, E.; Teye-Gaga, C.; Agyarkwah, S.K.; Adamtey, N. Development of black soldier fly, Hermetia illucens (Diptera: Stratiomyidae) in selected organic market waste fractions in Accra, Ghana. Asian J. Biotechnol. Bioresour. Technol. 2018, 4, 1–16. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; van Huis, A. Black soldier fly from pest to ‘crown jewel’ of the insects as feed industry: An historical perspective. J. Ins. Food Feed 2020, 6, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Stadtlander, T.; Stamer, A.; Buser, A.; Wohlfahrt, J.; Leiber, F.; Sandrock, C. Hermetia illucens meal as fish meal replacement for rainbow trout on farm. J. Ins. Food Feed 2017, 3, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Henry, M.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Review on the use of insects in the diet of farmed fish: Past and future. Anim. Feed Sci. Tech. 2015, 203, 1–22. [Google Scholar] [CrossRef]
- Mohan, K.; Rajan, D.K.; Muralisankar, T.; Ganesan, A.R.; Sathishkumar, P.; Revathi, N. Use of black soldier fly (Hermetia illucens L.) larvae meal in aquafeeds for a sustainable aquaculture industry: A review of past and future needs. Aquaculture 2022, 553, 738095. [Google Scholar] [CrossRef]
- Pérez-Pacheco, R.; Hinojosa-Garro, D.; Ruíz-Ortíz, F.; Camacho-Chab, J.C.; Ortega-Morales, B.O.; Alonso-Hernández, N.; Fonseca-Muñoz, A.; Landero-Valenzuela, N.; Loeza-Concha, H.J.; Diego-Nava, F.; et al. Growth of the black soldier fly Hermetia illucens (Diptera: Stratiomyidae) on organic-waste residues and its application as supplementary diet for Nile Tilapia Oreochromis niloticus (Perciformes: Cichlidae). Insects 2022, 13, 326. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; Shafi, M.E.; Alghamdi, W.Y.; Abdelnour, S.A.; Shehata, A.M.; Noreldin, A.E.; Ashour, E.A.; Swelum, A.A.; Al-Sagan, A.A.; Alkhateeb, M.; et al. Black soldier fly (Hermetia illucens) meal as a promising feed ingredient for poultry: A comprehensive review. Agriculture 2020, 10, 339. [Google Scholar] [CrossRef]
- Dörper, A.; Veldkamp, T.; Dicke, M. Use of black soldier fly and house fly in feed to promote sustainable poultry production. J. Ins. Food Feed 2021, 7, 761–780. [Google Scholar] [CrossRef]
- DiGiacomo, K.; Leury, B.J. Review: Insect meal: A future source of protein feed for pigs? Animal 2019, 13, 3022–3030. [Google Scholar] [CrossRef] [Green Version]
- Veldkamp, T.; Vernooij, A.G. Use of insect products in pig diets. J. Ins. Food Feed 2021, 7, 781–793. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Finke, M.D. Nutritional value of insects and ways to manipulate their composition. J. Ins. Food Feed 2021, 7, 639–659. [Google Scholar] [CrossRef]
- Heuel, M.; Sandrock, C.; Leiber, F.; Mathys, A.; Gold, M.; Zurbrügg, C.; Gangnat, I.D.M.; Kreuzer, M.; Terranova, M. Black soldier fly larvae meal and fat can completely replace soybean cake and oil in diets for laying hens. Poult. Sci. 2021, 100, 101034. [Google Scholar] [CrossRef] [PubMed]
- Heuel, M.; Kreuzer, M.; Sandrock, C.; Leiber, F.; Mathys, A.; Gold, M.; Zurbrügg, C.; Gangnat, I.D.M.; Terranova, M. Transfer of lauric and myristic acid from black soldier fly larval lipids to egg yolk lipids of hens is low. Lipids 2021, 56, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Heuel, M.; Sandrock, C.; Leiber, F.; Mathys, A.; Gold, M.; Zurbrügg, C.; Gangnat, I.D.M.; Kreuzer, M.; Terranova, M. Black soldier fly larvae meal and fat as a replacement for soybeans in organic broilers diets: Effects on performance, body N retention, carcass, and meat quality. Brit. Poult. Sci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.-X.; Tomberlin, J.; Vanlaerhoven, S. Ability of black soldier fly (Diptera: Stratiomyidae) larvae to recycle food waste. Environ. Entomol. 2015, 44, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Jucker, C.; Erba, D.; Leonardi, M.G.; Lupi, D.; Savoldelli, S. Assessment of vegetable and fruit substrates as potential rearing media for Hermetia illucens (Diptera: Stratiomyidae) larvae. Environ. Entomol. 2017, 46, 1415–1423. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; van Broekhoven, S.; van Huis, A.; van Loon, J.J.A. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [Green Version]
- Oonincx, D.G.A.B.; van Huis, A.; van Loon, J.J.A. Nutrient utilisation by black soldier flies fed with chicken, pig, or cow manure. J. Ins. Food Feed 2015, 1, 131–139. [Google Scholar] [CrossRef]
- Gold, M.; Cassar, C.M.; Zurbrügg, C.; Kreuzer, M.; Boulos, S.; Diener, S.; Mathys, A. Biowaste treatment with black soldier fly larvae: Increasing performance through the formulation of biowastes based on protein and carbohydrates. Waste Manag. 2020, 102, 319–329. [Google Scholar] [CrossRef]
- Scala, A.; Cammack, J.A.; Salvia, R.; Scieuzo, C.; Franco, A.; Bufo, S.A.; Tomberlin, J.K.; Falabella, P. Rearing substrate impacts growth and macronutrient composition of Hermetia illucens (L.) (Diptera: Stratiomyidae) larvae produced at an industrial scale. Sci. Rep. 2020, 10, 19448. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, Z.; Zhang, J.; Zhou, S.; Xiong, Q. Conversion of mixtures of soybean curd residue and kitchen waste by black soldier fly larvae (Hermetia illucens L.). Insects 2022, 13, 23. [Google Scholar] [CrossRef] [PubMed]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed—A review. J. Ins. Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Tschirner, M.; Simon, A. Influence of different growing substrates and processing on the nutrient composition of black soldier fly larvae destined for animal feed. J. Ins. Food Feed 2015, 1, 249–259. [Google Scholar] [CrossRef]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef]
- Lalander, C.; Diener, S.; Zurbrügg, C.; Vinnerås, B. Effects of feedstock on larval development and process efficiency in waste treatment with black soldier fly (Hermetia illucens). J. Clean. Prod. 2019, 208, 211–219. [Google Scholar] [CrossRef]
- Fuso, A.; Barbi, S.; Macavei, L.I.; Luparelli, A.V.; Maistrello, L.; Montorsi, M.; Sforza, S.; Caligiani, A. Effect of the rearing substrate on total protein and amino acid composition in black soldier fly. Foods 2021, 10, 1773. [Google Scholar] [CrossRef]
- Liland, N.S.; Biancarosa, I.; Araujo, P.; Biemans, D.; Bruckner, C.G.; Waagbo, R.; Torstensen, B.E.; Lock, E.J. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 2017, 12, e0183188. [Google Scholar] [CrossRef]
- Ewald, N.; Vidakovic, A.; Langeland, M.; Kiessling, A.; Sampels, S.; Lalander, C. Fatty acid composition of black soldier fly larvae (Hermetia illucens)—Possibilities and limitations for modification through diet. Waste Manag. 2020, 102, 40–47. [Google Scholar] [CrossRef]
- Liu, X.; Chen, X.; Wang, H.; Yang, Q.Q.; Rehman, K.U.; Li, W.; Cai, M.M.; Li, Q.; Mazza, L.; Zhang, J.B.; et al. Dynamic changes of nutrient composition throughout the entire life cycle of black soldier fly. PLoS ONE 2017, 12, e0182601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, M.; Tomberlin, J.K.; Diener, S.; Zurbrügg, C.; Mathys, A. Decomposition of biowaste macronutrients, microbes, and chemicals in black soldier fly larval treatment: A review. Waste Manag. 2018, 82, 302–318. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, I.; Newman, L.P.; Gill, H.; Danaher, J. The influence of food waste rearing substrates on black soldier fly larvae protein composition: A systematic review. Insects 2021, 12, 608. [Google Scholar] [CrossRef] [PubMed]
- Jensen, K.; Kristensen, T.; Heckmann, L.-H.; Sørensen, J. Breeding and maintaining high-quality insects. In Insects as Food and Feed: From Production to Consumtion; van Huis, A., Tomberlin, J.K., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2017; pp. 175–198. [Google Scholar]
- Rumbos, C.; Adamaki-Sotiraki, C.; Gourgouta, M.; Karapanagiotidis, I.; Asimaki, A.; Mente, E.; Athanassiou, C. Strain matters: Strain effect on the larval growth and performance of the yellow mealworm, Tenebrio molitor L. J. Ins. Food Feed 2021, 7, 1195–1205. [Google Scholar] [CrossRef]
- Zhou, F.; Tomberlin, J.K.; Zheng, L.Y.; Yu, Z.N.; Zhang, J.B. Developmental and waste reduction plasticity of three black soldier fly strains (Diptera: Stratiomyidae) raised on different livestock manures. J. Med. Entomol. 2013, 50, 1224–1230. [Google Scholar] [CrossRef]
- Park, S.; Choi, H.; Choi, J.-y.; Jeong, G. Population structure of the exotic black soldier fly, Hermetia illucens (Diptera: Stratiomyidae) in Korea. Korean J. Environ. Ecol. 2017, 31, 520–528. [Google Scholar] [CrossRef]
- Ståhls, G.; Meier, R.; Sandrock, C.; Hauser, M.; Šašić Zorić, L.; Laiho, E.; Aracil, A.; Doderović, J.; Badenhorst, R.; Unadirekkul, P.; et al. The puzzling mitochondrial phylogeography of the black soldier fly (Hermetia illucens), the commercially most important insect protein species. BMC Evol. Biol. 2020, 20, 60. [Google Scholar] [CrossRef]
- Bosch, G.; Oonincx, D.G.A.B.; Jordan, H.R.; Zhang, J.; van Loon, J.J.A.; van Huis, A.; Tomberlin, J.K. Standardisation of quantitative resource conversion studies with black soldier fly larvae. J. Ins. Food Feed 2020, 6, 95–109. [Google Scholar] [CrossRef]
- Kaya, C.; Generalovic, T.N.; Ståhls, G.; Hauser, M.; Samayoa, A.C.; Nunes-Silva, C.G.; Roxburgh, H.; Wohlfahrt, J.; Ewusie, E.A.; Kenis, M.; et al. Global population genetic structure and demographic trajectories of the black soldier fly, Hermetia illucens. BMC Biology 2021, 19, 94. [Google Scholar] [CrossRef]
- Eriksson, T.; Picard, C.J. Genetic and genomic selection in insects as food and feed. J. Ins. Food Feed 2021, 7, 661–682. [Google Scholar] [CrossRef]
- Zhan, S.; Fang, G.; Cai, M.; Kou, Z.; Xu, J.; Cao, Y.; Bai, L.; Zhang, Y.; Jiang, Y.; Luo, X.; et al. Genomic landscape and genetic manipulation of the black soldier fly Hermetia illucens, a natural waste recycler. Cell Res. 2020, 30, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Generalovic, T.N.; McCarthy, S.A.; Warren, I.A.; Wood, J.M.D.; Torrance, J.; Sims, Y.; Quail, M.; Howe, K.; Pipan, M.; Durbin, R.; et al. A high-quality, chromosome-level genome assembly of the black soldier fly (Hermetia illucens L.). G3 2021, 11, jkab085. [Google Scholar] [CrossRef] [PubMed]
- Oldenbroek, K.; van der Waaij, L. Textbook Animal Breeding and Genetics for BSc Students; Groen Kennisnet; Centre for Genetic Resources; The Netherlands and Animal Breeding and Genomics Centre: Wageningen, The Netherlands, 2015; Available online: http://wiki.groenkennisnet.nl/display/TAB/ (accessed on 30 November 2021).
- Sandrock, C.; Razmjou, J.; Vorburger, C. Climate effects on life cycle variation and population genetic architecture of the black bean aphid, Aphis fabae. Mol. Ecol. 2011, 20, 4165–4181. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.K.; Williams, S.; Springston, M.; Brown, J.; Freeman, K.; DesRoches, C.E.; Sokolowski, M.B.; Gibson, G. Genotype-by-diet interactions drive metabolic phenotype variation in Drosophila melanogaster. Genetics 2010, 185, 1009–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Günter, F.; Beaulieu, M.; Freiberg, K.F.; Welzel, I.; Toshkova, N.; Žagar, A.; Simčič, T.; Fischer, K. Genotype-environment interactions rule the response of a widespread butterfly to temperature variation. J. Evol. Biol. 2020, 33, 920–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meixner, M.D.; Kryger, P.; Costa, C. Effects of genotype, environment, and their interactions on honey bee health in Europe. Curr. Opin. Insect Sci. 2015, 10, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [Green Version]
- Goudet, J.; Jombart, T. hierfstat: Estimation and Tests of Hierarchical F-Statistics; R-package Version 0.04-22. 2015. Available online: httttps://CRAN.R-project/package=hierfstat (accessed on 6 May 2020).
- Paradis, E. pegas: An R package for population genetics with an integrated-modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef] [Green Version]
- Adamack, A.T.; Gruber, B. PopGenReport: Simplifying basic population genetic analyses in R. Methods Ecol. Evol. 2014, 5, 384–387. [Google Scholar] [CrossRef]
- Gruber, B.; Adamack, A.T. Landgenreport: A new R function to simplify landscape genetic analysis using resistance surface layers. Mol. Ecol. Resour. 2015, 15, 1172–1178. [Google Scholar] [CrossRef] [PubMed]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. Peerj 2014, 2, e281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamvar, Z.N.; Brooks, J.C.; Grünwald, N.J. Novel R tools for analysis of genome-wide population genetic data with emphasis on clonality. Front. Genet. 2015, 6, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, F.I.; Adams, P.E.; Schneiders, B.B. strataG: An R package for manipulating, summarizing and analysing population genetic data. Mol. Ecol. Resour. 2017, 17, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- Do, C.; Waples, R.S.; Peel, D.; Macbeth, G.M.; Tillett, B.J.; Ovenden, J.R. NeEstimator v2: Re-implementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Mol. Ecol. Resour. 2014, 14, 209–214. [Google Scholar] [CrossRef]
- Beugin, M.P.; Gayet, T.; Pontier, D.; Devillard, S.; Jombart, T.A.-O. A fast likelihood solution to the genetic clustering problem. Methods Ecol. Evol. 2018, 9, 1006–1016. [Google Scholar] [CrossRef] [Green Version]
- Akogul, S.; Erisoglu, M. A Comparison of information criteria in clustering based on mixture of multivariate normal distributions. Math. Comput. Appl. 2016, 21, 34. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis; Association of Official Analytical Chemists International,: Arlington, VA, USA, 1997. [Google Scholar]
- Janssen, R.H.; Vincken, J.P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-protein conversion factors for three edible insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef] [PubMed]
- VDLUFA. Das VDLUFA-Methodenbuch, Band III, Die Chemische Untersuchung von Futtermitteln. 2012. Available online: https://www.vdlufa.de/Methodenbuch/index.php?lang=de (accessed on 21 January 2019).
- Llames, C.R.; Fontaine, J. Determination of Amino Acids in Feeds: Collaborative Study. J. AOAC Int. 1994, 77, 1362–1402. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Poultry, 9 ed.; National Academies Press: Washington, DC, USA, 1994. [Google Scholar]
- Fox, J.; Weisberg, S.; Price, B.; Adler, D.; Bates, D.; Baud-Bovy, G.; Bolker, B. car: Companion to Applied Regression; R package version 3.0-3. 2019. Available online: https://CRAN. R-project.org/package=car (accessed on 14 May 2021).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.V. Least-squares means: The R package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Ripley, B.; Venables, B.; Bates, D.M.; Hornik, K.; Gebhardt, A.; Firth, D.; Ripley, M.B. Package ‘mass’. Cran r 2013, 538, 113–120. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneaous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Rosario-Martinez, H. Phia: Post-Hoc Interaction Analysis; R Package Version 0.2-1. 2015. Available online: https://CRAN. R-project.org/package=phia (accessed on 14 May 2021).
- Heuel, M.; Kreuzer, M.; Sandrock, C.; Leiber, F.; Mathys, A.; Guggenbühl, B.; Gangnat, I.D.M.; Terranova, M. Feeding value of black soldier fly larvae compared to soybean in methionine- and lysine-deficient laying hen diets. J. Ins. Food Feed 2022. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Swine, 9 ed.; National Academies Press: Washington, DC, USA, 1998. [Google Scholar]
- Koethe, M.; Taubert, J.; Vervuert, I. Impact of lysine supplementation on growth and development of Hermetia illucens larvae. J. Ins. Food Feed 2021, 8, 35–44. [Google Scholar] [CrossRef]
- Woods, M.; Hoffman, L.; Pieterse, E. Artificial diets for neonatal black soldier fly (Hermetia illucens) larvae. J. Ins. Food Feed 2019, 5, 99–105. [Google Scholar] [CrossRef]
- Bretscher, H.; O’Connor, M.B. The role of muscle in insect energy homeostasis. Front. Physiol. 2020, 11, 1254. [Google Scholar] [CrossRef] [PubMed]
- Gasco, L.; Józefiak, A.; Henry, M. Beyond the protein concept: Health aspects of using edible insects on animals. J. Ins. Food Feed 2021, 7, 715–741. [Google Scholar] [CrossRef]
- Chen, J.; Hou, D.; Pang, W.; Nowar, E.E.; Tomberlin, J.K.; Hu, R.; Chen, H.; Xie, J.; Zhang, J.; Yu, Z.; et al. Effect of moisture content on greenhouse gas and NH3 emissions from pig manure converted by black soldier fly. Sci. Total Environ. 2019, 697, 133840. [Google Scholar] [CrossRef] [PubMed]
- Ermolaev, E.; Lalander, C.; Vinnerås, B. Greenhouse gas emissions from small-scale fly larvae composting with Hermetia illucens. Waste Manag. 2019, 96, 65–74. [Google Scholar] [CrossRef]
- Pang, W.; Hou, D.; Chen, J.; Nowar, E.E.; Li, Z.; Hu, R.; Tomberlin, J.K.; Yu, Z.; Li, Q.; Wang, S. Reducing greenhouse gas emissions and enhancing carbon and nitrogen conversion in food wastes by the black soldier fly. J. Environ. Manag. 2020, 260, 110066. [Google Scholar] [CrossRef]
- Parodi, A.; De Boer, I.J.M.; Gerrits, W.J.J.; Van Loon, J.J.A.; Heetkamp, M.J.W.; Van Schelt, J.; Bolhuis, J.E.; Van Zanten, H.H.E. Bioconversion efficiencies, greenhouse gas and ammonia emissions during black soldier fly rearing—A mass balance approach. J. Clean. Prod. 2020, 271, 122488. [Google Scholar] [CrossRef]
- Parodi, A.; Gerrits, W.J.J.; Van Loon, J.J.A.; De Boer, I.J.M.; Aarnink, A.J.A.; Van Zanten, H.H.E. Black soldier fly reared on pig manure: Bioconversion efficiencies, nutrients in the residual material, greenhouse gas and ammonia emissions. Waste Manag. 2021, 126, 674–683. [Google Scholar] [CrossRef]
- Pang, W.; Hou, D.; Nowar, E.E.; Chen, H.; Zhang, J.; Zhang, G.; Li, Q.; Wang, S. The influence on carbon, nitrogen recycling, and greenhouse gas emissions under different C/N ratios by black soldier fly. Environ. Sci. Pollut. R. 2020, 27, 42767–42777. [Google Scholar] [CrossRef]
- Sandrock, C.; Walter, C.; Wohlfahrt, J.; Krauss, M.; Amelchanka, S.L.; Berard, J.; Leiber, F.; Kreuzer, M. Effects of feeding substrate on greenhouse gas emissions during black soldier fly larval development. In Proceedings of the 70th Annual Meeting of the European Federation of Animal Science, Ghent, Belgium, 26–30 August 2019; p. 156. [Google Scholar]
- Klammsteiner, T.; Walter, A.; Bogataj, T.; Heussler, C.D.; Stres, B.; Steiner, F.M.; Schlick-Steiner, B.C.; Arthofer, W.; Insam, H. The core gut microbiome of black soldier fly (Hermetia illucens) larvae raised on low-bioburden diets. Front. Microbiol. 2020, 11, 993. [Google Scholar] [CrossRef]
- Wynants, E.; Frooninckx, L.; Crauwels, S.; Verreth, C.; De Smet, J.; Sandrock, C.; Wohlfahrt, J.; Van Schelt, J.; Depraetere, S.; Lievens, B.; et al. Assessing the microbiota of black soldier fly larvae (Hermetia illucens) reared on organic waste streams on four different locations at laboratory and large scale. Microb. Ecol. 2019, 77, 913–930. [Google Scholar] [CrossRef]
- Bruno, D.; Bonelli, M.; De Filippis, F.; Di Lelio, I.; Tettamanti, G.; Casartelli, M.; Ercolini, D.; Caccia, S. The intestinal microbiota of Hermetia illucens larvae is affected by diet and shows a diverse composition in the different midgut regions. Appl. Environ. Microb. 2019, 85, e01864-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanga, C.M.; Waweru, J.W.; Tola, Y.H.; Onyoni, A.A.; Khamis, F.M.; Ekesi, S.; Paredes, J.C. Organic waste substrates induce important shifts in gut microbiota of black soldier fly (Hermetia illucens L.): Coexistence of conserved, variable, and potential pathogenic microbes. Front. Microbiol. 2021, 12, 24. [Google Scholar] [CrossRef] [PubMed]
- Shumo, M.; Khamis, F.M.; Ombura, F.L.; Tanga, C.M.; Fiaboe, K.K.M.; Subramanian, S.; Ekesi, S.; Schlüter, O.K.; van Huis, A.; Borgemeister, C. A molecular survey of bacterial species in the guts of black soldier fly larvae (Hermetia illucens) reared on two urban organic waste streams in Kenya. Front. Microbiol. 2021, 12, 687103. [Google Scholar] [CrossRef] [PubMed]
- Khamis, F.M.; Ombura, F.L.O.; Akutse, K.S.; Subramanian, S.; Mohamed, S.A.; Fiaboe, K.K.M.; Saijuntha, W.; Van Loon, J.J.A.; Dicke, M.; Dubois, T.; et al. Insights in the global genetics and gut microbiome of black soldier fly, Hermetia illucens: Implications for animal feed safety control. Front. Microbiol. 2020, 11, 1538. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, M.P.; Hull, K.L.; Brink-Hull, M.; Lloyd, M.; Rhode, C. Feed and host genetics drive microbiome diversity with resultant consequences for production traits in mass-reared black soldier fly (Hermetia illucens) larvae. Insects 2021, 12, 1082. [Google Scholar] [CrossRef]
- Zheng, L.Y.; Crippen, T.L.; Holmes, L.; Singh, B.; Pimsler, M.L.; Benbow, M.E.; Tarone, A.M.; Dowd, S.; Yu, Z.N.; Vanlaerhoven, S.L.; et al. Bacteria mediate oviposition by the black soldier fly, Hermetia illucens (L.), (Diptera: Stratiomyidae). Sci. Rep. 2013, 3, 2563. [Google Scholar] [CrossRef] [Green Version]
- De Smet, J.; Wynants, E.; Cos, P.; Van Campenhout, L. Microbial community dynamics during rearing of black soldier fly larvae (Hermetia illucens) and impact on exploitation potential. Appl. Environ. Microb. 2018, 84, e02722-17. [Google Scholar] [CrossRef] [Green Version]
- Tegtmeier, D.; Hurka, S.; Mihajlovic, S.; Bodenschatz, M.; Schlimbach, S.; Vilcinskas, A. Culture-independent and culture-dependent characterization of the black soldier fly gut microbiome reveals a large proportion of culturable bacteria with potential for industrial applications. Microorganisms 2021, 9, 1642. [Google Scholar] [CrossRef]
- Gold, M.; Fowles, T.; Fernandez-Bayo, J.D.; Miner, L.P.; Zurbrügg, C.; Nansen, C.; Bischel, H.N.; Mathys, A. Effects of rearing system and microbial inoculation on black soldier fly larvae growth and microbiota when reared on agri-food by-products. J. Ins. Food Feed 2021, 8, 113–127. [Google Scholar] [CrossRef]
- Schreven, S.J.J.; de Vries, H.; Hermes, G.D.A.; Smidt, H.; Dicke, M.; van Loon, J.J.A. Relative contributions of egg-associated and substrate-associated microorganisms to black soldier fly larval performance and microbiota. FEMS Microbiol. Ecol. 2021, 97, fiab054. [Google Scholar] [CrossRef]
- Vorburger, C.; Sandrock, C.; Gouskov, A.; Castaneda, L.; Ferrari, J. Genotypic variation and the role of defensive endosymbionts in an all-parthenogenetic host-parasitoid interaction. Evolution 2009, 63, 1439–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castañeda, L.E.; Sandrock, C.; Vorburger, C. Variation and covariation of life history traits in aphids are related to infection with the facultative bacterial endosymbiont Hamiltonella defensa. Biol. J. Linn. Soc. 2010, 100, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Vogel, M.; Shah, P.N.; Voulgari-Kokota, A.; Maistrou, S.; Aartsma, Y.; Beukeboom, L.W.; Falcao Salles, J.; Van Loon, J.J.A.; Dicke, M.; Wertheim, B. Health of the black soldier fly and house fly under mass-rearing conditions: Innate immunity and the role of the microbiome. J. Ins. Food Feed 2022, in press. [Google Scholar] [CrossRef]
- Bonelli, M.; Bruno, D.; Brilli, M.; Gianfranceschi, N.; Tian, L.; Tettamanti, G.; Caccia, S.; Casartelli, M. Black soldier fly larvae adapt to different food substrates through morphological and functional responses of the midgut. Int. J. Mol. Sci. 2020, 21, 4955. [Google Scholar] [CrossRef]
- Paz, A.S.P.; Carrejo, N.S.; Rodriguez, C.H.G. Effects of larval density and feeding rates on the bioconversion of vegetable waste using black soldier fly larvae Hermetia illucens (L.), (Diptera: Stratiomyidae). Waste Biomass Valori. 2015, 6, 1059–1065. [Google Scholar]
- Pastor, B.; Velasquez, Y.; Gobbi, P.; Rojo, S. Conversion of organic wastes into fly larval biomass: Bottlenecks and challenges. J. Ins. Food Feed 2015, 1, 179–193. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Influence of larval density and dietary nutrient concentration on performance, body protein, and fat contents of black soldier fly larvae (Hermetia illucens). Entomol. Exp. Appl. 2018, 166, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Moyers, B.T.; Morrell, P.L.; McKay, J.K. Genetic Costs of domestication and improvement. J. Hered. 2018, 109, 103–116. [Google Scholar] [CrossRef] [Green Version]
- Vijendravarma, R.K.; Kraaijeveld, A.R.; Godfray, H.C.J. Experimental evolution shows Drosophila melanogaster resistance to a microsporidian pathogen has fitness costs. Evolution 2009, 63, 104–114. [Google Scholar] [CrossRef]
- Van der Most, P.J.; de Jong, B.; Parmentier, H.K.; Verhulst, S. Trade-off between growth and immune function: A meta-analysis of selection experiments. Funct. Ecol. 2011, 25, 74–80. [Google Scholar] [CrossRef]
- Schwenke, R.A.; Lazzaro, B.P.; Wolfner, M.F. Reproduction–immunity trade-offs in insects. Annu. Rev. Entomol. 2016, 61, 239–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruno, D.; Montali, A.; Mastore, M.; Brivio, M.F.; Mohamed, A.; Tian, L.; Grimaldi, A.; Casartelli, M.; Tettamanti, G. Insights into the immune response of the black soldier fly larvae to bacteria. Front. Immunol. 2021, 12, 4866. [Google Scholar] [CrossRef] [PubMed]
- Ho, P.N.; Klanrit, P.; Hanboonsong, Y.; Yordpratum, U.; Suksawat, M.; Kulthawatsiri, T.; Jirahiranpat, A.; Deewai, S.; Mackawan, P.; Sermswan, R.W.; et al. Bacterial challenge-associated metabolic phenotypes in Hermetia illucens defining nutritional and functional benefits. Sci. Rep. 2021, 11, 23316. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Ge, C.; Yao, H. Antimicrobial peptides from black soldier fly (Hermetia illucens) as potential antimicrobial factors representing an alternative to antibiotics in livestock Farming. Animals 2021, 11, 1937. [Google Scholar] [CrossRef]
- Vogel, H.; Müller, A.; Heckel, D.G.; Gutzeit, H.; Vilcinskas, A. Nutritional immunology: Diversification and diet-dependent expression of antimicrobial peptides in the black soldier fly Hermetia illucens. Dev. Comp. Immunol. 2018, 78, 141–148. [Google Scholar] [CrossRef]
- Sheppard, D.C.; Tomberlin, J.K.; Joyce, J.A.; Kiser, B.C.; Sumner, S.M. Rearing methods for the black soldier fly (Diptera: Stratiomyidae). J. Med. Entomol. 2002, 39, 695–698. [Google Scholar] [CrossRef] [Green Version]
- Bertinetti, C.; Samayoa, A.C.; Hwang, S.Y. Effects of feeding adults of Hermetia illucens (Diptera: Stratiomyidae) on longevity, oviposition, and egg hatchability: Insights into optimizing egg production. J. Insect Sci. 2019, 19, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gobbi, P.; Martinez-Sanchez, A.; Rojo, S. The effects of larval diet on adult life-history traits of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Eur. J. Entomol. 2013, 110, 461–468. [Google Scholar] [CrossRef]
- Cammack, J.A.; Tomberlin, J.K. The Impact of diet protein and carbohydrate on select life-history traits of the black soldier fly Hermetia illucens (L.) (Diptera: Stratiomyidae). Insects 2017, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Barragan-Fonseca, K.; Pineda-Mejia, J.; Dicke, M.; van Loon, J.J.A. Performance of the black soldier fly (Diptera: Stratiomyidae) on vegetable residue-based diets formulated based on protein and carbohydrate contents. J. Econ. Entomol. 2018, 111, 2676–2683. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.; Gort, G.; Dicke, M.; van Loon, J. Nutritional plasticity of the black soldier fly (Hermetia illucens) in response to artificial diets varying in protein and carbohydrate concentrations. J. Ins. Food Feed 2021, 7, 51–61. [Google Scholar] [CrossRef]
- Bellezza Oddon, S.; Biasato, I.; Gasco, L. Isoenergetic-practical and semi-purified diets for protein requirement determination in Hermetia illucens larvae: Consequences on life history traits. J. Anim. Sci. Biotechnol. 2022, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, A.C.; Montali, A.; Bruno, D.; Tettamanti, G. Metabolic adjustment of the larval fat body in Hermetia illucens to dietary conditions. J. Asia-Pac. Entomol. 2017, 20, 1307–1313. [Google Scholar] [CrossRef]
- Zhu, Z.; Rehman, K.U.; Yu, Y.; Liu, X.; Wang, H.; Tomberlin, J.K.; Sze, S.-H.; Cai, M.; Zhang, J.; Yu, Z.; et al. De novo transcriptome sequencing and analysis revealed the molecular basis of rapid fat accumulation by black soldier fly (Hermetia illucens, L.) for development of insectival biodiesel. Biotechnol. Biofuels 2019, 12, 194. [Google Scholar] [CrossRef]
- Hoc, B.; Genva, M.; Fauconnier, M.L.; Lognay, G.; Francis, F.; Caparros Megido, R. About lipid metabolism in Hermetia illucens (L. 1758): On the origin of fatty acids in prepupae. Sci. Rep. 2020, 10, 11916. [Google Scholar] [CrossRef]
- Giannetto, A.; Oliva, S.; Ceccon Lanes, C.F.; de Araújo Pedron, F.; Savastano, D.; Baviera, C.; Parrino, V.; Lo Paro, G.; Spanò, N.C.; Cappello, T.; et al. Hermetia illucens (Diptera: Stratiomydae) larvae and prepupae: Biomass production, fatty acid profile and expression of key genes involved in lipid metabolism. J. Biotechnol. 2020, 307, 44–54. [Google Scholar] [CrossRef]
- Rozkosný, R. A Biosystematic Study of the European Stratiomyidae (Diptera), vol. 2. In Series Entomologica vol. 25: Clitellariinae, Hermetiinae, Pachygasterinae and Bibliography; Spencer, K.A., Ed.; Dr W. Junk Publishers: Hague, The Netherlands, 1983; pp. 172–176. [Google Scholar]
- Nylin, S.; Gotthard, K. Plasticity in life-history traits. Annu. Rev. Entomol. 1998, 43, 63–83. [Google Scholar] [CrossRef] [Green Version]
- Blanckenhorn, W.U. Causes and consequences of phenotypic plasticity in body size: The case of the yellow dung fly Scathophaga stercoraria (Diptera: Scathophagidae). In Phenotypic Plasticity of Insects: Mechanisms and Consequences; Whitman, D.W., Ananthakrishnan, T.N., Eds.; Science Publishers, Inc.: Enfield, Australia, 2009; pp. 369–422. [Google Scholar]
- Freda, P.J.; Ali, Z.M.; Heter, N.; Ragland, G.J.; Morgan, T.J. Stage-specific genotype-by-environment interactions for cold and heat hardiness in Drosophila melanogaster. Heredity 2019, 123, 479–491. [Google Scholar] [CrossRef]
- Samayoa, A.C.; Hwang, S.Y. Degradation capacity and diapause effects on oviposition of Hermetia illucens (Diptera: Stratiomyidae). J. Econ. Entomol. 2018, 111, 1682–1690. [Google Scholar] [CrossRef]
- Gao, Z.; Wang, W.; Lu, X.; Zhu, F.; Liu, W.; Wang, X.; Lei, C. Bioconversion performance and life table of black soldier fly (Hermetia illucens) on fermented maize straw. J. Clean. Prod. 2019, 230, 974–980. [Google Scholar] [CrossRef]
- Yaşar, B.; Çirik, T. Life tables of Hermetia illucens (Linnaeus, 1758)(Diptera: Stratiomyidae) on different foods. J. Nat. Appl. Sci. 2018, 22, 392–398. [Google Scholar] [CrossRef]
- Brede, N.; Sandrock, C.; Straile, D.; Spaak, P.; Jankowski, T.; Streit, B.; Schwenk, K. The impact of human-made ecological changes on the genetic architecture of Daphnia species. Proc. Natl. Acad. Sci. USA 2009, 106, 4758–4763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandrock, C.; Schirrmeister, B.E.; Vorburger, C. Evolution of reproductive mode variation and host associations in a sexual-asexual complex of aphid parasitoids. BMC Evol. Biol. 2011, 11, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roffeis, M.; Wakefield, M.E.; Almeida, J.; Alves Valada, T.R.; Devic, E.; Koné, N.G.; Kenis, M.; Nacambo, S.; Fitches, E.C.; Koko, G.K.D.; et al. Life cycle cost assessment of insect based feed production in West Africa. J. Clean. Prod. 2018, 199, 792–806. [Google Scholar] [CrossRef] [Green Version]
- Ewusie, E.A.; Kwapong, P.K.; Ofosu-Budu, G.; Sandrock, C.; Akumah, A.M.; Nartey, E.K.; Tetegaga, C.; Agyakwah, S.K. The black soldier fly, Hermetia illucens (Diptera:Stratiomyidae): Trapping and culturing of wild colonies in Ghana. Sci. Afr. 2019, 5, e00134. [Google Scholar]
- Rhode, C.; Badenhorst, R.; Hull, K.; Greenwood, M.; Bester, A.; Andere, A.; Picard, C.J.; Richards, C. Genetic and phenotypic consequences of early domestication in black soldier flies (Hermetia illucens). Anim. Genet. 2020, 51, 752–762. [Google Scholar] [CrossRef]
- Bang, A.; Courchamp, F. Industrial rearing of edible insects could be a major source of new biological invasions. Ecol. Lett. 2021, 24, 393–397. [Google Scholar] [CrossRef]
- Berggren, Å.; Jansson, A.; Low, M. Approaching ecological sustainability in the emerging insects-as-food industry. Trends Ecol. Evol. 2019, 34, 132–138. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Item | Poultry Feed (PF) | Food Waste (FW) | Poultry Manure (PM) |
|---|---|---|---|
| Total ash | 132.4 | 35.3 | 257.5 |
| Carbon | 440.5 | 468.2 | 381.0 |
| Nitrogen | 32.3 | 24.5 | 46.4 |
| Ether extract | 50.0 | 52.2 | 23.0 |
| Neutral detergent fibre | 299.1 | 317.3 | 399.3 |
| Acid-detergent fibre | 132.2 | 102.1 | 298.6 |
| Hemicellulose | 166.9 | 215.2 | 100.7 |
| Strain | Pairwise FST | FIS | Hobs | Hexp | AR | AU | Ne | ||
|---|---|---|---|---|---|---|---|---|---|
| S1 | S2 | S3 | |||||||
| S1 | 0.129 | 0.496 | 0.563 | 4.000 | 0.108 | 110 | |||
| S2 | 0.330 | 0.135 | 0.432 | 0.494 | 3.867 | 0.074 | 193 | ||
| S3 | 0.307 | 0.112 | 0.021 | 0.551 | 0.557 | 4.467 | 0.114 | 267 | |
| S4 | 0.132 | 0.349 | 0.335 | −0.013 | 0.528 | 0.516 | 3.667 | 0.066 | 147 |
| Diet | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Statistics | Poultry Feed (PF) | Food Waste (FW) | Poultry Manure (PM) | |||||||||||||
| Parameter | Model; Test | Strain (DF = 3) | Diet (DF = 2) | S × D (DF = 6) | Strain S1 | Strain S2 | Strain S3 | Strain S4 | Strain S1 | Strain S2 | Strain S3 | Strain S4 | Strain S1 | Strain S2 | Strain S3 | Strain S4 |
| Larval live weight (mg) at harvest | i; F: | 634.2 *** | 10497.2 *** | 27.3 *** (10) | 253.0 a | 181.5 c | 201.8 b | 199.7 b | 226.4 e | 168.9 h | 216.6 f | 207.5 g | 120.1 i | 89.4 l | 100.0 k | 102.6 j |
| Larval mortality (%) | ii; F: | 41.1 *** | 15.9 *** | 6.6 *** (3) | 5.14 a | 2.03 ab | 0.67 b | 0.65 b | 5.00 e | 0.64f | 0.20f | 0.83f | 0.41 | 0.70 | 0.36 | 0.24 |
| Larval biomass (g DM) | i; F: | 848.6 *** | 17312.8 *** | 29.7 *** (11) | 107.5 a | 80.7 b | 78.0 b | 77.5 b | 97.6 e | 72.4 h | 88.6 f | 83.3 g | 34.0 i | 26.9 j | 27.9 j | 27.5 j |
| Estimated protein (EP; g N × 4.76/kg larval DM) | iv; χ2: | 135.5 *** | 1037.5 *** | 42.3 *** (6) | 303 c | 322 a | 313 b | 319 ab | 272 g | 300 e | 290 f | 307 e | 339 j | 347 j | 358 i | 356 i |
| Total estimated protein (EP; g) | i; F: | 505.0 *** | 21,638.8 *** | 37.6 *** (12) | 32.6 a | 26.0 b | 24.4 c | 24.7 bc | 26.5 e | 21.7 f | 25.6 e | 25.5 e | 11.5 i | 9.3 k | 10.0 j | 9.8 jk |
| Ether extract (EE; g/kg larval DM) | iv, χ2: | 20.7 *** | 1439.7 *** | 101.7 *** (11) | 279 b | 232 b | 381 a | 395 a | 500 e | 441 f | 454 ef | 485 ef | 78 j | 140 i | 90 j | 71 j |
| Total ether extract (EE; g) | i; F: | 99.1 *** | 6925.8 *** | 8.8 *** (8) | 29.7 a | 18.7 b | 29.7 a | 30.7 a | 48.8 e | 31.9 g | 40.2 f | 40.4 f | 2.7 i | 3.8 i | 2.5 i | 2.0 j |
| Estimated protein: ether extract ratio (EP:EE) | i; F: | 686.3 *** | 3432.5 *** | 14.5 *** (13) | 1.13 ab | 1.41 a | 0.82 b | 0.81 b | 0.55 f | 0.68 e | 0.64 e | 0.63 e | 4.38 ij | 2.66 j | 3.97 j | 5.06 i |
| Total ash (TA; g/kg larval DM) | iii; χ2: | 59.5 *** | 43,845.0 *** | 172.4 *** (12) | 112 a | 101 bc | 102 b | 9.7 c | 24 f | 28 e | 30 e | 31 e | 213 k | 201 l | 217 jk | 223 i |
| Total ash (TA; g) | i; F: | 912.3 *** | 10372.4 *** | 48.9 *** (15) | 12.0 a | 8.2 b | 8.0 b | 7.6 b | 2.3 g | 2.1 h | 2.7 e | 2.6 f | 7.3 i | 5.4 k | 6.1 j | 6.2 j |
| Bioconversion efficiency (BE; g larvae/kg diet; DM-based) | ii; F: | 990.3 *** | 18997.2 *** | 42.7 *** (12) | 343 a | 256 b | 248 b | 247 b | 225 e | 166 h | 204 f | 192 g | 84 i | 66 j | 69 j | 68 j |
| Nitrogen efficiency (N-BE; g N larvae/kg N diet) | ii; F: | 2510.0 *** | 42850.3 *** | 58.5 *** (12) | 674 a | 535 b | 503 c | 510 c | 522 e | 424 f | 505 e | 488 e | 128 i | 102 k | 111 j | 109 j |
| Dry matter (DM) reduction (g/kg diet) | ii; F: | 5807.6 *** | 23536.8 *** | 13.4 *** (10) | 638 a | 527 c | 631 a | 600 b | 771 g | 697 h | 786 f | 802 e | 359 j | 344 k | 366 j | 375 i |
| Neutral detergent fibre (NDF) reduction (g/kg diet) | ii; F: | 533.7 *** | 2118.8 *** | 1.5 (0) | 593 | 598 | 599 | 585 | 833 | 814 | 840 | 832 | 189 ij | 175 ij | 233 i | 166 j |
| Acid detergent fibre (ADF) reduction (g/kg diet) | ii; F: | 48.9 *** | 368.6 *** | 0.9 (0) | 479 | 456 | 499 | 461 | 531 | 525 | 552 | 587 | 146 | 154 | 215 | 179 |
| Hemicellulose (HC) reduction (g/kg diet) | ii; F: | 255.0 *** | 2469.0 *** | 11.6 *** (8) | 684 | 710 | 678 | 683 | 976 ef | 952 f | 977 e | 960 ef | 316 i | 237 i | 285 i | 128 j |
| Dry matter (DM) losses (g emissions/kg diet) | iv, χ2: | 297.7 *** | 5986.1 *** | 97.0 *** (7) | 294 c | 270 d | 381 a | 353 b | 546 g | 545 g | 582 f | 610 e | 274 k | 278 jk | 297 ij | 307 i |
| Amino Acid (g/100 g protein) | Effect | Poultry Feed (PF) | Food Waste (FW) | Poultry Manure (PM) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BSFL | BSFL | BSFL | ||||||||||||||||
| Strain (DF = 3) | Diet (DF = 2) | S × D (DF = 6) | Strain S1 | Strain S2 | Strain S3 | Strain S4 | Strain S1 | Strain S2 | Strain S3 | Strain S4 | Strain S1 | Strain S2 | Strain S3 | Strain S4 | ||||
| Essential | ||||||||||||||||||
| His 1 | 13.72 *** | 321.47 *** | 4.51 *** (2) | 2.61 | 3.37 c A | 3.45 c D | 3.53 b G | 3.65 a J | 2.62 | 3.37 g A | 3.46 f D | 3.43 fg H | 3.57 e J | 2.09 | 2.90 j B | 3.14 i E | 3.03 ij I | 3.10 i K |
| Ile 1 | 0.72 | 1.92 | 0.74 | 4.10 | 4.57 | 4.53 | 4.57 | 4.60 JK | 4.18 | 4.52 | 4.50 | 4.58 | 4.47 K | 4.68 | 4.57 | 4.55 | 4.58 | 4.74 J |
| Leu1 | 12.80 *** | 3.40 * | 2.46 * (1) | 9.21 | 7.67 a B | 7.54 b | 7.60 ab | 7.71 ab J | 7.46 | 7.88 e A | 7.63 f | 7.64 f | 7.56 f JK | 8.10 | 7.65 i B | 7.58 ij | 7.56 ij | 7.49 j K |
| Lys | 32.95 *** | 174.45 *** | 6.68 *** (4) | 5.06 | 6.62 b B | 7.07 a E | 7.04 a H | 6.81 b K | 4.28 | 6.81 f B | 7.19 e E | 6.62 f I | 6.63 f K | 5.83 | 7.35 j A | 7.76 i D | 7.40 j G | 7.47 j J |
| Met1 | 3.53 * | 13.19 *** | 18.19 *** (11) | 2.39 | 2.05 b B | 2.19 a D | 2.11 b G | 2.13 ab | 2.27 | 2.28 e A | 2.15 f D | 1.95 g H | 2.10 f | 1.85 | 1.95 k B | 2.04 jk E | 2.12 ij G | 2.15 i |
| Phe1 | 42.64 *** | 68.75 *** | 7.30 *** (5) | 5.45 | 4.92 b A | 5.15 a D | 4.91 b G | 4.89 b J | 5.31 | 4.87 ef A | 4.89 e E | 4.72 fg H | 4.72 g K | 4.44 | 4.69 j B | 4.77 ij F | 4.84 ij GH | 4.86 i J |
| Thr1 | 58.51 *** | 33.09 *** | 8.32 *** (4) | 3.73 | 4.08 B | 4.07 D | 4.06 H | 4.06 K | 3.59 | 4.30 A | 4.19 D | 4.20 G | 4.18 JK | 5.21 | 4.09 j B | 3.97 k E | 4.15 ij GH | 4.26 i J |
| Val 1 | 8.36 *** | 15.30 *** | 4.91 *** (3) | 4.78 | 6.22 c | 6.47 b D | 6.71 a G | 6.59 a J | 4.99 | 6.45 ef | 6.13 f E | 6.40 ef H | 6.55 e JK | 6.28 | 6.21 | 6.46 D | 6.27 H | 6.36 K |
| Non-essential | ||||||||||||||||||
| Ala1 | 580.40 *** | 142.26 *** | 16.57 *** (9) | 5.67 | 8.29 a B | 7.75 b E | 7.22 c H | 7.20 c K | 4.37 | 9.34 e A | 8.12 f D | 8.12 f G | 7.80 g J | 9.69 | 7.95 i B | 7.46 j F | 7.36 j H | 7.58 j J |
| Arg1 | 63.38 *** | 425.00 *** | 48.50 *** (15) | 6.64 | 5.39 bc A | 5.37 c E | 5.57 ab G | 5.65 a J | 5.76 | 4.65 g B | 5.77 e D | 5.59 f G | 5.70 ef J | 4.38 | 4.81 j B | 5.16 i F | 4.90 j H | 4.77 j K |
| Asx | 18.71 *** | 2.01 | 9.38 *** (4) | 8.17 | 9.17 a | 9.14 a D | 8.75 b G | 8.74 b | 6.34 | 9.06 e | 8.64 f E | 9.02 e H | 8.67 f | 9.25 | 9.00 i | 8.59 j E | 9.28 i H | 8.59 j |
| Cys | 0.97 | 832.00 *** | 11.77 *** (9) | 1.95 | 0.96 a B | 0.98 a E | 1.00 a H | 0.90 b L | 2.56 | 0.97 B | 1.00 E | 0.99 H | 0.97 K | 2.70 | 1.36 i A | 1.29 j D | 1.27 j G | 1.37 i J |
| Glx1 | 198.53 *** | 784.70 *** | 13.53 *** (9) | 19.63 | 11.26 B | 11.18 E | 11.15 H | 11.11 L | 23.57 | 10.31 g C | 11.33 ef E | 11.16 f H | 11.42 e K | 13.68 | 13.38 i A | 12.76 jk D | 12.52 k G | 13.10 j L |
| Gly1 | 37.30 *** | 65.37 *** | 10.29 *** (6) | 4.39 | 5.54 a B | 5.39 b E | 5.41 b H | 5.59 a K | 3.75 | 5.87 e A | 5.44 f E | 5.48 f H | 5.45 f L | 6.95 | 5.90 i A | 5.66 k D | 5.69 jk G | 5.86 ij J |
| Pro1 | 24.52 *** | 87.01 *** | 5.43 *** (3) | 7.23 | 6.14 B | 6.13 DE | 6.07 H | 6.48 J | 9.73 | 6.90 e A | 6.27 f D | 6.68 e G | 6.54 ef J | 5.48 | 5.69 ij C | 5.88 i E | 5.85 i H | 5.51 j K |
| Ser | 1.94 | 11.45 *** | 1.05 | 4.98 | 4.34 | 4.13 | 4.20 H | 4.10 K | 5.75 | 4.42 | 4.37 | 4.46 G | 4.52 J | 5.19 | 4.46 | 4.30 | 4.34 GH | 4.29 JK |
| Tyr1 | 219.90 *** | 293.08 *** | 10.83 *** (6) | 2.89 | 7.75 b A | 7.82 b D | 8.41 a G | 8.18 a J | 2.36 | 6.58 f B | 7.33 e E | 7.30 e H | 7.48 e K | 3.10 | 6.51 k B | 7.02 j F | 7.19 i H | 6.86 j L |
| Specific protein (g/kg DM) 1 | 26.40 *** | 230.63 *** | 11.83 *** (12) | 196.4 | 362.6 c A | 384.2 a D | 373.8 b H | 375.7 b J | 139.8 | 311.3 g B | 351.2 e E | 338.4 f I | 355.0 e K | 93.9 | 360.3 k A | 383.8 i D | 389.4 i G | 372.0 j J |
| Kp (×N) 1 | 23.77 *** | 360.48 *** | 2.52 * (0) | 6.08 | 5.70 ab A | 5.68 ab D | 5.69 a G | 5.61 b J | 5.70 | 5.46 B | 5.58 D | 5.56 H | 5.51 K | 2.02 | 5.07 j C | 5.28 i E | 5.18 i I | 4.97 j L |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandrock, C.; Leupi, S.; Wohlfahrt, J.; Kaya, C.; Heuel, M.; Terranova, M.; Blanckenhorn, W.U.; Windisch, W.; Kreuzer, M.; Leiber, F. Genotype-by-Diet Interactions for Larval Performance and Body Composition Traits in the Black Soldier Fly, Hermetia illucens. Insects 2022, 13, 424. https://doi.org/10.3390/insects13050424
Sandrock C, Leupi S, Wohlfahrt J, Kaya C, Heuel M, Terranova M, Blanckenhorn WU, Windisch W, Kreuzer M, Leiber F. Genotype-by-Diet Interactions for Larval Performance and Body Composition Traits in the Black Soldier Fly, Hermetia illucens. Insects. 2022; 13(5):424. https://doi.org/10.3390/insects13050424
Chicago/Turabian StyleSandrock, Christoph, Simon Leupi, Jens Wohlfahrt, Cengiz Kaya, Maike Heuel, Melissa Terranova, Wolf U. Blanckenhorn, Wilhelm Windisch, Michael Kreuzer, and Florian Leiber. 2022. "Genotype-by-Diet Interactions for Larval Performance and Body Composition Traits in the Black Soldier Fly, Hermetia illucens" Insects 13, no. 5: 424. https://doi.org/10.3390/insects13050424