
Enantiomeric Discrimination in Insects: The Role of OBPs and ORs


1
Biointeractions and Crop Protection Department, Rothamsted Research, Harpenden, Hertfordshire AL5 2JQ, UK
2
School of Chemistry, University of Nottingham, University Park, Nottingham NG7 2RD, UK
*
Author to whom correspondence should be addressed.
Insects 2022, 13(4), 368; https://doi.org/10.3390/insects13040368
Submission received: 1 March 2022
/
Revised: 25 March 2022
/
Accepted: 30 March 2022
/
Published: 8 April 2022
(This article belongs to the Special Issue Recent Advances in Physiology of Insect Olfaction)
Abstract
:Simple Summary
Insects use olfaction, i.e., their sense of smell, to detect odors that elicit behavioral responses, with structurally similar compounds eliciting different responses. The roles of specific recognition proteins, i.e., odorant-binding proteins (OBPs) and odorant receptors (ORs), located in insect antennae, in discriminating between structurally similar compounds are not fully understood. Here, we explore current research in understanding the role of OBPs and ORs in discriminating between enantiomers—mirror image structures—in insect chemical ecology and chemoperception.
Abstract
Olfaction is a complex recognition process that is critical for chemical communication in insects. Though some insect species are capable of discrimination between compounds that are structurally similar, little is understood about how this high level of discrimination arises. Some insects rely on discriminating between enantiomers of a compound, demonstrating an ability for highly selective recognition. The role of two major peripheral olfactory proteins in insect olfaction, i.e., odorant-binding proteins (OBPs) and odorant receptors (ORs) has been extensively studied. OBPs and ORs have variable discrimination capabilities, with some found to display highly specialized binding capability, whilst others exhibit promiscuous binding activity. A deeper understanding of how odorant-protein interactions induce a response in an insect relies on further analysis such as structural studies. In this review, we explore the potential role of OBPs and ORs in highly specific recognition, specifically enantiomeric discrimination. We summarize the state of research into OBP and OR function and focus on reported examples in the literature of clear enantiomeric discrimination by these proteins.
1. Introduction
Insects use chemistry to communicate in sexual reproduction, prey and natural enemy location/avoidance, oviposition and host location. Insect chemical communication occurs mainly via olfaction, i.e., the recognition and discrimination of olfactory ligands by olfactory proteins mainly located in the antennae. Although insects possess other chemosensory systems, such as the gustatory (taste) system, olfaction remains one of the most critical senses in insects [1,2]. Elucidating the chemistry and physiology underpinning insect olfaction provides an opportunity to better understand insect behavior [3]. Furthermore, as many insect species are either agricultural pests or harmful to human health, understanding insect olfaction also provides an opportunity to develop novel pest management approaches that are effective, economically viable and sustainable [4].
Chirality is common in nature and many insect species utilize chiral olfactory ligands for communication [5,6]. Furthermore, in contrast to hormones or other internal chemical communication pathways, insect olfaction is an external recognition process, meaning that olfactory ligands need to be recognized amongst a plethora of background signals. This implies that the olfactory recognition system in insects involving olfactory proteins must be highly sophisticated [5].
Though enantiomeric discrimination is a critical biochemical process, the mechanisms by which olfactory proteins can discriminate between structurally similar compounds with high levels of specificity remain unclear. The complex, membrane-bound nature of some olfactory proteins provides a challenge for their study in vitro [7,8], meaning that very few membrane-bound protein structures have been reported to date [7,9]. Furthermore, although extensive studies have been conducted on soluble olfactory proteins and their interaction with ligands [10,11,12,13,14,15], the specific role of such proteins remains largely debatable, with some proteins demonstrating high levels of specificity and others being highly promiscuous [16,17,18]. Where extensive functional analysis of olfactory proteins has been conducted, studies have frequently neglected to screen multiple stereoisomers or enantiomers of an olfactory ligand, most likely due to the expense and difficulty of synthesizing non-naturally occurring enantiomers, compared to naturally-occurring enantiomers which are often easily derived from plant sources [19].
Understanding how enantiomeric discrimination arises, specifically the role of membrane-bound and soluble olfactory proteins, will provide a greater fundamental understanding of how sophisticated insect olfactory recognition systems function. This could lead to novel practical applications, such as monitoring and management of significant agricultural pests [4].
2. Chirality in Insect Olfaction
2.1. Semiochemistry
Chemical communication between organisms is mediated by semiochemicals that can be categorized into either pheromones, which are compounds that are released by an organism and induce a response in individuals of the same species, or allelochemicals, which induce responses in individuals of different species [20,21]. Different types of pheromones include sex pheromones, aggregation pheromones and alarm pheromones [20]. Pheromones can be further categorized into releasers, which induce an immediate behavioral change, and primers, which initiate a complex set of physiological or developmental changes but may result in no immediate behavioral change [21]. Allelochemicals can also be further categorized depending on whether the emitter or receiver is the beneficiary [21]. Semiochemicals are used in sexual reproduction, prey and natural enemy avoidance, oviposition and host location. Though widely employed by insects for communication, semiochemicals have been identified in many other organisms including mammals, birds and fish [22,23,24]. Many semiochemical-based interactions, especially pheromone interactions, are highly specific. This specificity may arise from the use of species-specific compounds, or the use of the same compounds but at either different ratios/concentrations or with different stereochemistries [6,25].
2.2. Chiral Pheromones
Insect species across a broad range of taxa utilize chiral pheromone components. Identification of the specific stereochemistry and bioactivity of chiral pheromones requires the synthesis of specific enantiomers of pheromone components [6]. Synthesis of enantiomerically pure pheromones was first achieved in the 1970s with the synthesis of compounds such as (S)-4-methyl-3-heptanone 1, the alarm pheromone of leaf-cutting ant Atta texana, (7R,8S)-(+)-disparlure 2, the sex pheromone of the female gypsy moth Lymantria dispar and (1S,5R)-frontalin 3, an aggregation pheromone of the western pine beetle, Dendroctonus brevicomis (Figure 1) [6,26,27,28]. Since then, hundreds of enantiomerically pure pheromone components have been synthesized [6].
Generally, insects utilize a specific enantiomer as a chiral pheromone component and do not respond to the other enantiomer in the same way [6]. However, the other enantiomer may inhibit pheromone activity, may have differing effects on males and females, or have variable levels of bioactivity dependent on their stereochemical similarity to the most active enantiomer [6]. This level of variation of bioactivity of pheromones clearly indicates a complex recognition system, the mechanism and specificity of which may vary between different species. The attractiveness to males of (R)-japonilure 4, the female-produced sex pheromone of the Japanese beetle, Popillia japonica, is inhibited by (S)-japonilure 5, resulting in a lack of biological response when a racemate is used to study beetle behavior [29] (Figure 2). Even minor contamination by (S)-japonilure can inhibit male responses to (R)-japonilure. The olive fruit fly, Bactrocera oleae, produces a racemic mixture of (R) and (S)-malic acid (6 and 7), with the (R)-isomer acting as the male-attractive sex pheromone and the (S)-isomer attracting females [30]. (6R,10R)-Matsuone 8 is a sex pheromone of red pine scale, Matsucoccus resinosae, whereas the (6R,10S) isomer 9 is active but to a lesser degree [31].
Pheromones may also comprise mixtures of different compounds or mixtures of enantiomers. Aphids utilize a sex pheromone, generally consisting of two components (1R,4aS,7S,7aR)-nepetalactol 10 and (4aS,7S,7aR)-nepetalactone 11 [32,33]. Some aphids, such as the rosy apple aphid, Dysaphis plantaginea and the damson-hop aphid, Phorodon humuli, also employ other isomers including (1R,2S,5S)-dolichodial 12, (1S,4aR,7S,7aS)-nepetalactol 13 and (1R,4aR,7S,7aS)-nepetalactol 14 [25,34]. Although the aphid sex pheromone components are ubiquitous across most species, the ratio of the components is species-specific [19,35]. For example, for the pea aphid, Acyrthosiphon pisum, the sex pheromone consists of a 1:1 ratio, whereas the black-bean aphid, Aphis fabae, uses a very high ratio of lactone to lactol [35]. Though some pheromones comprise two different compounds with the same stereochemistry in all shared positions, some pheromones may comprise a mixture where both enantiomers are necessary for a biological response, eg. Sulcatol, the aggregation pheromone of the ambrosia beetle, Gnathotrichus sulcatus, comprises a 35:65 mixture of (R)-sulcatol 15 and (S)-sulcatol 16 respectively [36].
In addition to volatile pheromone components, some insect species, mainly Hymenoptera, utilize cuticular hydrocarbons (CHCs) for intraspecific communication. Stereochemistry is often conserved within a species, eg. in a study of 36 methyl-branched CHCs across 20 species, all CHCs were found to possess (R) stereochemistry [37].
2.3. Other Chiral Semiochemicals
Chirality is also crucial for the bioactivity of other semiochemicals eg. plant volatile organic compounds (VOCs), which play a significant role in plant-insect interactions. Insect host location required for pollination, herbivory or oviposition may require high levels of specificity in semiochemicals similar to that for intraspecific interactions [38,39]. Dependent on the plant-insect relationship, some plant VOCs may be attractive and some may be repellent to insects [38].
Plant VOCs can be identical to pheromone components, such as (4aS,7S,7aR)-nepetalactone 11 which, in addition to its role as an aphid sex pheromone component, is emitted by the catmint plant Nepeta cataria and repellent to many insect species [40,41]. One stereoisomer of α-pinene 17, a plant-produced isoprenoid, is also known as an aggregation pheromone component for some Ips species of bark beetle [42]. Furthermore, other Coleoptera species, such as the red turpentine beetle, Dendroctonus valens, show differential responses to enantiomers of α-pinene [43]. Additional VOCs of isoprenoid origin may have differing effects depending on their chirality. Linalool, which can exist as either the (R)-18 or (S)-19 form, has been frequently shown to have different behavioral or physiological activities depending on which enantiomer is utilized [44,45].
3. Insect Olfactory Proteins
3.1. Introduction to Insect Olfaction
Olfaction (i.e., the recognition and discrimination of olfactory ligands) is critical for the perception of pheromones and other semiochemicals. Prior to the identification of the first insect pheromones, little was understood regarding the mechanisms underlying insect olfaction, with knowledge being mainly based on anecdotal evidence [46].
The first insect pheromone to be characterized was Bombykol ((10E,12Z)-hexadec-10,12-dien-1-ol) 20 (Figure 3), identified from the silkworm moth, Bombyx mori [15,46,47,48]. Due to the large physical size of this insect, a significant quantity of pheromone was isolated and initial structural identification was achieved using infrared (IR) and ultraviolet (UV) spectroscopy. Since the first identification of pheromones, significant advances in the field of olfaction research were made by developments in analytical chemistry. Introduction of techniques for the identification of complex chemical structures, particularly nuclear magnetic resonance (NMR) spectroscopy and coupled gas chromatography-mass spectrometry (GC-MS) for the detection of minute quantities of compounds, were used to confirm the identification of Bombykol 20 [15,46,47], and vastly expanded the known chemical library of pheromones and other semiochemicals [46]. The identification of the previously discussed aphid sex pheromone components, (1R,4aS,7S,7aR)-nepetalactol 10 and (4aS,7S,7aR)-nepetalactone 11, relied on such techniques [32,49].
The main olfactory organs in insects are the antennae, although some insects possess additional olfactory organs, such as maxillary palps [50,51]. Insect antennae generally comprise segments and are covered in small hair-like structures called sensilla [17,51,52,53,54]. Olfactory sensilla are perforated with pores, through which odorant molecules diffuse. Each sensillum can be uniporous (one pore) or multiporous (many pores) [17,52]. Sensilla contain an aqueous fluid, or sensillum lymph, in which olfactory receptor neurons (ORNs) are bathed, and where olfactory proteins (i.e., odorant-binding proteins (OBPs) and olfactory receptors (ORs)) are located [17]. There is no similarity between the OBPs and ORs in structure, but both appear to play a role in olfaction and interact with ligands, including pheromones and semiochemicals. Specific ORs have been mapped to specific sensilla types [17,51].
Diversity in function has been observed across sensilla. There are a few morphological classes of sensilla found in most insects; basionic, trichoid, placoid and coeloconic [17,50,51,54]. Each morphological class is generally responsible for a different function. In Drosophila, the basionic sensilla are responsible for fruit odors, trichoid sensilla for pheromones, and coeloconic sensilla for organic acid and amine-based odors [17]. For vetch aphids, Megoura viciae, placoid sensilla bear pores indicating a chemosensory role [53]. Variation in sensilla abundance and morphology can be observed between different insects that have different chemosensory profiles [54].
Olfactory neuroanatomy is an extensive field of research that focuses on insect olfaction, including ORNs that express ORs [50,51,55,56]. In Mammalia, ORNs are bipolar, allowing dendrites to give rise to numerous specialized cilia and providing a large receptive surface for the binding of odors to ORs [56]. Removal of these specialized cilia in mammals removes associated olfactory responses [56]. ORs are activated and hypothesized to generate an action potential, which travels along an ORN that glomerulates and converges in the brain [51,56,57,58].
Significant developments in the study of odorant perception have been furthered by recent advances in insect genomics. With full genome sequences now available for many insect species, including model organisms such as Drosophila melanogaster, many genes for OBPs and ORs have been identified [59]. Furthermore, by using advanced molecular biology techniques, the function of these receptors and proteins can be more extensively studied [17,18,58,59]. Advances in molecular and structural biology techniques allow for more intensive study of the structure and function of olfactory proteins, via heterologous expression, structural characterization and knockout or knockdown studies with RNAi and CRISPR/Cas9 gene editing [60].
3.2. Odorant-Binding Proteins
OBPs are a unique group of olfactory proteins found in high abundance in the sensillum lymph, the aqueous fluid that can be found within the sensilla of the antennae [61]. These proteins are highly structurally conserved across insects and the high abundance of mRNA encoding for OBPs found in the antennae suggests they play an important role in olfaction [17].
The first OBP to be identified in invertebrates was from an extract of the large moth, Antheraea polyphemus, using a radioactively labeled pheromone in ligand-binding experiments [61]. Initially, OBPs could be categorized by their six conserved cysteines, which results in a similar 3D structure despite diverging amino acid sequences [61]. However, further research showed a greater diversity of 3D structure, and three distinct categories have now been defined; classic OBPs (possessing six highly conserved cysteines that form disulfide bridges), Plus-C OBPs (possessing eight conserved cysteines and one conserved proline) and atypical (possessing nine or ten conserved cysteines) [62]. The conserved cysteines and multiple disulfide bridges lead to the high thermal stability of these proteins. OBPs range in size from approximately 110 to 240 residues, usually resulting in proteins of 10–25 kDa in size [62]. In addition to OBPs, another family of proteins, known as chemosensory proteins or CSPs, have been described. CSPs show similar binding activity to OBPs, but no sequence similarities, and only share four conserved cysteines [63].
Though thermally stable, OBPs are flexible globular proteins that may occur in multiple conformations. Alternate conformations may arise as a result of conditions such as pH change or binding activity. Pheromone-binding proteins (PBPs) found in B. mori and other insect species show pH-dependent conformational changes [12,64,65,66,67,68,69]. This change is often associated with the binding and release of a ligand [64].
Numerous roles have been proposed for OBPs via studies on a range of insect species [16,18,61,70]. Many insect odorant molecules are highly lipophilic and are poorly soluble in aqueous solutions, which indicates these proteins may play a role in solubilizing or transporting the ligands to the odorant receptors in the aqueous sensillum lymph. OBPs have been shown to reversibly bind behaviorally active olfactory ligands, suggesting they play a role in olfactory perception [71,72,73,74,75,76,77]. LUSH, one of the most widely studied OBPs found in D. melanogaster [17,78,79], is thought to play a role in recognition and response to the male sex pheromone (Z)-vaccenyl acetate (VA) 21, and has been shown to bind to VA in vitro, as well as other insect pheromones, short-chain alcohols and phthalates (Figure 4) [17]. Bombyx mori OBPs and PBPs are capable of discriminating between B. mori sex pheromone components and bombykol [74,80,81], whilst OBPs in aphids have been shown to discriminate between the alarm pheromone, (E)-β-farnesene 22, and other ligands (Figure 4) [11,72,82,83]. Though OBPs have been shown to discriminate some ligands from others in binding assays, the specificity of OBPs varies widely and the fluorescence techniques utilized for studying OBP-ligand interactions are not optimal for observing subtle differences in ligand binding.
Expression of the gene encoding for a specific OBP can be suppressed, resulting in lower levels of gene expression, or the DNA encoding for the gene can be knocked out, entirely removing the expression of the gene [60]. Deletion may lead to reductions in electrophysiological responses, spontaneous neuron firing and behavioral effects [50,55,82,84,85,86,87]. However, some studies have shown olfactory neuron responses are still functional after deletion of LUSH, and LUSH-deficient mutants do not show any behavioral defects [17,55]. The existence of such contradicting studies only furthers the need for a thorough understanding of the role of OBPs.
In addition to suggesting roles for OBPs and CSPs in olfaction, genomic and proteomic studies generally show that many OBPs or CSPs are not found within the primary sensory organs (antennae) and are expressed in a wide diversity of spatial patterns, suggesting they possess roles beyond olfaction. For the honeybee, Apis mellifera, only 12 of 21 identified OBPs and two of six CSPs have been identified within the antennae [17,18,88]. Though some OBPs are expressed in the gustatory (taste) system of some species, as well as in larval chemosensory organs (Drosophilia), many OBPs and CSPs may possess entirely non-chemosensory roles [16,17,70,89]. OBPs and PBPs have been found in the pheromonal or ejaculatory glands of insects [70,90,91,92]. CSPs have also been linked to development [70,93]. In addition to pheromone transport and development, OBPs and CSPs play a role in a variety of other biological processes. These include anti-inflammatory action in disease-carrying insects, humidity sensing in Drosophila and other roles in nutrition, vision, migration and insecticide resistance [16,70,89].
3.3. Odorant Receptors
ORs are found across the animal kingdom. For mammals, ORs belong to a group of proteins known as G-coupled protein receptors (GPCRs) [94], a large, diverse family of seven transmembrane (7TM) or heptahelical proteins with an extracellular N-terminus and intracellular C-terminus [56,95]. Although insect ORs are also 7TM proteins, they are distinct from GPCRs and possess an inverse heptahelical topology, with the N-terminus being located in the intracellular section of the transmembrane protein and the C-terminus found extracellularly (Figure 5) [7,9,58]. This suggests that insect ORs are a unique protein family, different from all other chemosensory receptors [58]. Insect ORs also differ in signal transduction and form a unique class of heteromeric cation channels [7,8,9,96].
ORs are a much larger and more diverse group of receptors in mammals. However, most ORs found in insects are co-expressed with another OR, known as ORCO (odorant receptor co-receptor, identified as OR83b in D. melanogaster) [1,58,97]. ORCO is structurally similar to other insect ORs, however, it is highly conserved across different insect species—only 20% conservation is seen between ORs, however approximately 60% shared identity can be seen for ORCO [7]. Basal insects lack an ORCO homolog due to their small number of OR genes, expressing homomeric OR complexes instead [9,98]. There is also no known mammalian ortholog of ORCO [58].
As with OBPs, knockdown or knockout studies contribute to the understanding of the function of ORs in insects. ORCO-deficient mutants show significant loss of olfactory function and related behaviors, indicating that ORCO must play an important role in olfactory signal transduction [99,100,101]. Antennal lobe glomeruli were also reduced in some ORCO-deficient mutants and antennal lobe size was affected by the knock-out of a sex pheromone receptor in Spodoptera littoralis [99,100,101,102]. OR binding activity is generally investigated using electrophysiological techniques, where the receptor is expressed in a membrane, exposed to a ligand and electrophysiological responses measured. Presently, a lack of structural data reduces options for in silico studies due to challenges associated with studying membrane-bound proteins [7,9,103,104,105].
4. Enantiomeric Discrimination by OBPs
OBPs are the first recognition proteins involved in the process of insect olfaction. After entering the antennae from the air via a pore, semiochemicals come into contact with OBPs contained within the aqueous sensillum lymph [18,61,70,106]. Although the specific role of OBPs in olfaction is unclear, they are capable of discrimination between different compounds and should therefore be evaluated for enantioselectivity [71,72,73,74,75,76,77]. OBPs exist in both insects and mammals, with similar structures. Mammalian PBPs have been shown to possess chiral discrimination properties, with a single residue conferring specificity to chiral compounds in a pig OBP [107]. However, OBPs are notoriously promiscuous binders, and finding an OBP capable of high levels of discrimination between any compounds is challenging.
4.1. OBP Specificity
Insects are capable of discriminating, at the behavioral and physiological level, between structurally similar compounds, including those with only small modifications or stereochemical differences, such as the sex pheromone components of aphids [32]. OBPs may assist in this process or potentially bind as an OBP-ligand complex to ORs. Supporting evidence for this hypothesis would include the occurrence of a conformational change, induced in OBPs when biologically active ligands bind. D. melanogaster OBP LUSH possesses a salt bridge between Lys87 and Asp118 that is only present in the apo (unbound) structure (non-VA-bound 3D- structure). When this salt bridge was disrupted in LUSH mutants, the DmOR69d neurons were activated in the absence of VA [55]. This suggests that LUSH is conformationally activated by VA, and in turn activates ORNs, or that a VA/LUSH complex interacts with ORNs [55]. OBPs and PBPs from different insect species also possess comparable salt bridges in their OBPs. B. mori possesses a salt bridge between Lys89 and Glu125 that is structurally analogous to LUSH [74].
Additional evidence for a conformation change, or conformation activation of OBPs generally indicates a C-terminal folding domain [13,64,74,108]. This C-terminus folding may be dictated by pH changes, where the acidic-residue-rich C-terminus loses negative charge at a low pH and forms an additional α-helix [108]. This additional helix can then enter the binding pocket and displace any ligand present [108]. This conformation activation has been observed in a range of species and could be responsible for interactions at the OR, where the OBP may expel the ligand for OR-binding, or the OBP may bind itself in a protein-protein interaction [66,67,68,69]. Some OBPs have also been shown to dimerize, and it is possible this dimerization could be disrupted by conformational changes, suggesting alternative ligand binding and release mechanisms [12].
OBPs generally have large binding pockets and can flexibly adapt to fit a multitude of ligands, often multiple ligands at once, leading to low levels of specificity [63]. It has been proposed that specificity might arise from numerous factors including the conformational changes resulting in active and inactive conformers, in addition to the energy minima and entropic contributions of ligands fitting into the ligand-binding pocket [109]. Ligands with lower conformational constraint, such as long-chain poly-unsaturated fatty acids that are often found as Lepidopteran pheromones, may be more difficult to discriminate compared with more conformationally constrained compounds, such as bicyclic terpenes. The more flexible ligands have more degrees of freedom, allowing them to fit into the available space within the ligand-binding sites. An OBP demonstrating high levels of discrimination will likely have a smaller binding site capable of accepting very specific, highly conformationally constrained compounds.
4.2. Enantiomeric Discrimination by Insect OBPs
OBPs are not thought to be good candidates for enantiomeric discrimination [110]. Olfactory responses to different enantiomers occur within the same sensilla; two species of a scarab beetle, i.e., the Osaka beetle, Anomala osakana and the Japanese beetle, Popillia japonica, utilize different enantiomers of japonilure 4 and 5, yet possess identical OBPs in the activated sensilla [110]. However, some OBPs and PBPs, have been identified that exhibit some degree of enantiomeric discrimination ability. PBP1 and PBP2, from the gypsy moth, Lymantria dispar, show differential responses to the sex pheromone component (7R,8S)-(+)-disparlure 2, and its behaviorally antagonist enantiomer (7S,8R)-(−)-disparlure 23 (Figure 6) [10,111]. In a radioactive ligand-binding assay, LdisPBP1 was shown to have a higher affinity for (7S,8R)-(−)-disparlure 23 whereas LdisPBP2 responded with a higher affinity to (7R,8S)-(+)-disparlure 2. Additionally, Apol-3, the PBP from the wild silk moth, Antheraea polyphemus, bound to both enantiomers with a low affinity, capable of discriminating both enantiomers [111]. Structural studies confirmed this binding activity and the conformational changes that these PBPs undergo at different pH, although LdisPBP1 was seen to bind both enantiomers of disparlure in NMR experiments, it bound the (-)-enantiomer with a higher affinity and interacts with different residues [10,109].
In the case of LdisPBPs and other examples, the kinetic activity of the protein, specifically the binding and release mechanism, may be important in determining specificity [10,12,67,108,112,113]. Furthermore, as the two enantiomers of disparlure result in different behavioral responses in gypsy moths, this would indicate that PBPs not only play a role in enantiomeric discrimination, but also suggests a discriminatory or recognition role in insect olfaction in general. Results thus far suggest that OBPs, in general, are not capable of enantiomeric discrimination, but there may exist PBPs with a high level of specificity. How the discriminatory ability of these peripheral proteins may affect olfactory responses in the insect is also unclear.
5. Enantiomeric Discrimination by ORs
Insect ORs play a significant role in determining insect responses to olfactory ligands. However, limited evidence is available confirming the structure of insect ORs, the specific activation of insect ORs by olfactory ligands and enantioselectivity in insect ORs. Enantioselectivity has been demonstrated in mammalian ORs, however, insect and mammalian ORs are vastly different in their structure and function [9,56,58,114,115]. Activation of ORs results in an electrophysiological response and, as it is clear that enantiomeric discrimination is critical in insect olfaction and unlikely to arise from OBPs, ORs are a better candidate for exploring enantioselectivity.
5.1. OR Specificity
Insect ORs can be broadly tuned or highly specific. Broadly tuned receptors show responses to a broad range of compounds, often with vastly different chemical structures [9,116,117]. For example, when ApisOR4, from the pea aphid, A. pisum, was screened against a panel of 57 odorants, it responded to a range of aromatic compounds, such as 4-ethylacetophenone 24 and salicylaldehyde 25, in addition to bicyclic terpene compounds such as (S)-cis-verbenol 26 [118] (Figure 7). A total of six ligands were found to activate ApisOR4 with similar responses, all plant-derived volatiles [118]. Despite the binding of different ligands activating the same receptor, discrimination between ligands may come at a higher level. In mammals, odor perception has shown to be combinatorially coded (i.e., multiple ORs are activated in unique combinations as a response to a specific odor) [119]. Although little is known about odor coding in insects, evidence for distinctive neuronal perception between attractive and repellent odors has been identified [120].
By contrast, highly specific ORs respond to one or a few compounds, usually with a high level of specificity. This may include discrimination between similar chemistries, isomers or enantiomers. In Lepidoptera, there exist many ORs that are classified as pheromone receptors (PRs) due to their specific response to a single pheromone compound [117]. These receptors have been shown to discriminate between very similar chemical structures, though in Lepidoptera these are generally achiral compounds. Moth pheromones are generally Type I, straight-chain fatty alcohols and corresponding acetates and aldehydes, or Type II, long-chain polyunsaturated hydrocarbons, although there are exceptions (e.g., the sex pheromone (7R,8S)-(+)-disparlure 2 of the female gypsy moth Lymantria dispar and (1S,5R)-frontalin 3). Highly specific receptors have conserved motifs, e.g., moth PRs show a conserved C-terminal region, the functional significance of which is unknown [117]. Additionally, in ants there exists a large family ‘9-exon’ ORs which have been shown to be specific for CHCs and candidate pheromones [120].
Little is known about the origins of specificity in insect ORs, though a range of functional assays including mutation of specific loci within the OR subunit has been performed. Single amino acid residues often dictate specificity—polymorphism or mutation of one specific residue often results in loss of function or different activity in insect ORs [9,105,121,122,123,124,125,126,127]. This suggests a tight and highly specific binding site, though further investigation is required to determine how this may impact enantiomeric discrimination.
5.2. Enantioselective Insect ORs
Due to the low specificity of soluble periphery proteins in insect olfaction (i.e., OBPs or PBPs), it seems logical that ORs comprise the major discriminatory system in insect olfaction. However, as previously stated, not all ORs show high levels of specificity, with very few demonstrating activity with one of the two enantiomers despite the activity of this specificity being observed in electrophysiological and behavioral studies (Table 1). Additionally, few functional studies of insect ORs screen multiple enantiomers of chiral components due to the inaccessibility of non-naturally occurring components.
An enantioselective OR has been identified in the Yellow Fever mosquito, Aedes aegypti [128]. AaegOR8 co-expressed with the A. aegypti ORCO (AaegOR7) was shown to have a significantly higher response to the kairomone (R)-1-octen-3-ol 27 than the respective enantiomer, (S)-1-octen-3-ol 28 (Figure 8) [128,129], although a small response was observed from (S)-1-octen-3-ol 28 and was attributed to a 0.1% enantiomeric impurity [129]. Similar activity has been observed between conserved receptors from other mosquito species (Toxorhynchites amboinensis, Culex quinquefasciatus and Anopheles gambiae) [122,130,131,132]. An OR from Anopheles gambiae, AgamOR29, also shows differential responses to the racemate (R/S)-linalool and the specific enantiomer (R)-linalool 18, suggesting enantiomeric specificity [45]. Furthermore, two ORs from the Eurasian spruce bark beetle, Ips typographus, were shown to possess enantio-discriminatory ability. After initial responses to the racemic mixtures of pheromone components, (R/S)-ipsenol (29 and 30) and (R/S)-ipsdienol (31 and 32) (Figure 8), were observed, enantiospecific functional assays were conducted. ItypOR46 was shown to be most responsive to (S)-ipsenol 29 and ItypOR49 to (R)-ipsdienol 31 [104]. Some ORs have also been shown to respond to multiple enantiomers, though some enantiomers induce slightly higher responses. ORs from the cerambycid beetle, Megacyllene caryae, display flexibility when responding to various enantiomers of pheromonal components 2-methyl-1-butanol (33 and 34) and 2,3-hexanediol (35, 36, 37 and 38) [133].
In most cases of enantioselective insect ORs, a racemic mixture of the two components was screened before enantiospecific functional assays were conducted. This further highlights the need for additional exploration where responses are seen from a racemic mixture. Chirality holds biological importance, and it is unlikely two enantiomers of a compound will have an identical activity to one another.
Observed enantiomeric discrimination by insect ORs indicates either a highly specific binding site, or multiple potential sites with only one site activating the receptor. Multiple binding sites have been predicted in insect ORs. Molecular modeling and docking identified two potential binding sites for ligands in ItypOR46 [104]. Other studies of ligand-binding interactions with insect ORs have suggested the existence of multiple binding sites. A study investigating potential allosteric agonist interactions with A. gambiae ORCO demonstrated that different ligands may bind simultaneously to different sites [134]. Additionally, mutations in the C-terminal helix (7) of CquiOR8 altered enantioselectivity, indicating the importance of specific loci within the OR unit [122]. Studies with achiral ligands, such as (Z)-11-tetradecenyl acetate and Lepidopteran ORs also suggested there may exist multiple ORs for the same compound that recognize different conformations or stereochemistries [135]. Presently, there appear to be few examples of enantiomeric discrimination by ORs, though this may be due to difficulty functionally screening with multiple enantiomers or focus on insect species with no chiral pheromone components.
5.3. Structural Studies of Insect ORs
Despite extensive functional studies, the structure of insect OR-ORCO complexes remains elusive. The structure of the ORCO from the fig wasp, Apocrypta bakeri was determined as a heteromeric tetramer by Butterwick et al. (2018) using cryo-EM techniques [7]. This has led to large advances in knowledge of the assembly and functionality of ORCO-OR tetramers, including their properties as an ion channel for olfactory signal transduction. Structurally, ORCO can be divided into two domains including four loose peripheral transmembrane domains and a single central anchor domain [7]. Cryo-EM data also confirmed the inverse heptahelical topology [7,9]. Comprehensive structural data can be used to sufficiently model and assess the binding activity of ORs and explain the range of critical residues that have been identified [9,104,124,125,126,127].
Following elucidation of the structure of ORCO from A. bakeri, a structure for a homomeric broadly tuned insect OR was elucidated [9]. The structure of OR5 from the jumping bristletail, Machilidae harabi, was elucidated as a homomeric OR (MharOR5) activated by a broad range of ecologically relevant olfactory ligands, including the mosquito repellent DEET and eugenol [9]. The ligand binding and channel opening of the OR was resolved, revealing a clear ligand-binding pocket and hydrophobic interactions between the ligands and critical residues. The ligand-binding site was a 15Å-deep pocket within the extracellular portion of the complex, which was shown to be highly conserved across different insect species [9]. Assessment with other ligands also demonstrated this binding pocket to be highly promiscuous, but unable to accommodate all ligands which are known to activate or inhibit the receptor, suggesting the potential for multiple binding sites or additional allosteric sites [9].
The architecture of the odorant-binding pocket of the M. harabi OR5 included highly conserved residues that have previously been identified as important in ligand binding [9]. Before a full structure of an insect OR was available, data relied on mutagenesis studies. Maps of critical loci within the OR could be generated by mutating suspected key residues and observing functional changes. Multiple mutagenesis experiments have been performed with ORCO and ORs across a variety of species, many of which aligned with key sections of the final ORCO structure [7]. Mutations in both extracellular loop 2 (EL2) and transmembrane domain 3 (TM3) both generally resulted in responses to olfactory ligands, either altering, reducing or abolishing specificity [105,126,136,137,138]. This functional change can also occur with mutations in the N- and C-termini, transmembrane domains 2,4 and 6 (TM2, TM4 and TM6), extracellular loop 3 (EL3) and intracellular loop 3 (IL3) [123,138,139,140,141,142,143]. Intracellular loop 2 (IL2) is predicted to be a calmodium, a Ca2+ regulator protein, binding site, and IL2, transmembrane domains 5–7 (TM6) all affect current and ion selectivity and permeability due to their location around the ORCO-OR complex ion channel pore (Figure 9) [7,96,124,127,138,139,142,144].
Investigations of OR function can utilize computational techniques, specifically modeling and docking analysis, with the limited data that are available [103,104,105,145]. Additional structural data will only improve the accuracy and viability of these techniques [103,104,105,145]. Unfortunately, no structural data exist to date for either a highly specific OR or a heteromeric complex between an OR and ORCO. Progress in understanding how specificity arises in insect ORs, and the subsequent mode of action, will rely on advances in structural biology studies.
6. Future Research Directions
6.1. Alternative Targets
Enantioselectivity may arise elsewhere in the insect olfactory system. Enantioselective ORNs have been described, such as an ORN from the cabbage moth, Mamestra brassica, that is most responsive to (R)-linalool 18, but it is predicted this selectivity arises from the OR expressed within the neuron [44]. Additionally, pheromone-degrading enzymes (PDEs) have demonstrated chiral discrimination in P. japonica [146].
Another group of proteins commonly studied in the insect olfactory system is ionotropic receptors (IRs). IRs have many roles in insects, including olfactory perception, in addition to taste, hygroperception (humidity sensing) and cool temperature sensing [147]. IRs, like ORs, are ligand-gated cation channels, though they possess a different general structure. IRs are thought to possess a similar domain topology and overall homology with glutamate receptors [2]. IRs and ORs do not overlap in odor specificity, and IRs mainly respond to amines, aldehydes and acids [147]. IRs have the potential to be enantioselective as shown by selective responses to L-amino acids in Drosophila taste perception [148].
6.2. Applications of Enantiomeric Discrimination
Many insect species have indirect and direct detrimental effects on human health. Providing a fundamental understanding of insect olfactory systems and enantioselectivity of pheromones and other semiochemicals has the potential to underpin future pest management approaches [3,4,149]. An understanding of how specificity occurs in insect olfaction will shed light on the overall olfactory recognition process. Presently, it is unclear how this specificity arises, and often olfactory proteins are shown to be broadly tuned to a multitude of ligands. Elucidation of differences between highly specific and broadly tuned OBPs and ORs could provide insights into the evolution of specific olfactory responses and discrimination between complex mixtures of compounds. Beyond fundamental understanding, understanding enantiomeric discrimination in insect olfaction may have potential practical applications. These may include manipulation of the insect olfactory system or design of novel olfactory ligands for pest management, and the use of olfactory proteins or modified olfactory proteins in biosensor systems [5,150,151].
7. Conclusions
Enantiomeric discrimination plays an important role in insect olfaction and insect chemical ecology. However, how highly specific recognition arises is still unclear. First, the specific role or mode of action of OBPs remains unclear. Though multiple studies explore OBP-ligand binding specificity, it is unclear whether conformational changes, induced by ligand binding and other physiological changes, are a critical component of OBPs function. Investigation into OBP activity needs to consider the screening of multiple enantiomers and not only the racemate or biologically active enantiomer. Furthermore, a more in-depth analysis of the structure and conformational changes induced in OBPs is required. Finally, it appears high specificity only arises in PBPs that specifically bind pheromones. The only example of enantiomeric discrimination by OBPs is by PBPs from L. dispar, which show differential responses to the sex pheromone component (7R,8S)-(+)-disparlure 2, and the enantiomer (7S,8R)-(-)-disparlure 3 [10,111]. Refinement of OBP classification may be required to easily divide general or non-specific OBPs and highly specific PBPs and their structure, conformation changes and subsequent role.
ORs are a much more convincing target for enantiomeric discrimination. Examples of enantiomeric discrimination by insect Ors exist, indicating these proteins must play an important role in this important process [45,104,122,128,130,133]. Unfortunately, little is understood about the activation of the receptor and opening of the ion channel by a ligand. With such limited structural data available, predictions and explanations of activity are difficult. Mutagenesis studies provide some insight, but additional structural data are needed for progress in understanding these proteins.
Overall, there are very few examples of enantiomeric discrimination by peripheral olfactory proteins in insects. Additional structural and functional data are required for better understanding, and future studies investigating the ligand-binding ability of OBPs and ORs should include both enantiomers of a chiral ligand where possible, especially in the case of highly specific pheromones.
Author Contributions
All authors contributed equally to conception of the review. Research and writing, C.S.; supervision and funding, D.M.W. and M.A.B. All authors contributed to proofreading and editing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
CS was supported by a Biotechnology and Biological Sciences (BBSRC) Doctoral Training Partnership studentship (reference 1804053). The work at Rothamsted formed part of the Smart Crop Protection (SCP) strategic programme (BBS/OS/CP/000001) funded through the Biotechnology and Biological Sciences Research Council’s Industrial Strategy Challenge Fund.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors would like to acknowledge the Biotechnology and Biological Sciences Research Council for funding contributions (BBSRC).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sánchez-Gracia, A.; Vieira, F.G.; Rozas, J. Molecular evolution of the major chemosensory gene families in insects. Heredity 2009, 103, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Rytz, R.; Croset, V.; Benton, R. Ionotropic Receptors (IRs): Chemosensory ionotropic glutamate receptors in Drosophila and beyond. Insect Biochem. Mol. Biol. 2013, 43, 888–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reisenman, C.E.; Lei, H.; Guerenstein, P.G. Neuroethology of olfactory-guided behavior and its potential application in the control of harmful insects. Front. Physiol. 2016, 7, 271. [Google Scholar] [CrossRef] [PubMed]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex pheromones and their impact on pest management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef] [PubMed]
- Touchet, S.; Chamberlain, K.; Woodcock, C.M.; Miller, D.J.; Birkett, M.A.; Pickett, J.A.; Allemann, R.K. Novel olfactory ligands via terpene synthases. Chem. Commun. 2015, 51, 7550–7553. [Google Scholar] [CrossRef] [Green Version]
- Mori, K. Significance of chirality in pheromone science. Bioorganic Med. Chem. 2007, 15, 7505–7523. [Google Scholar] [CrossRef]
- Butterwick, J.A.; del Mármol, J.; Kim, K.H.; Kahlson, M.A.; Rogow, J.A.; Walz, T.; Ruta, V. Cryo-EM structure of the insect olfactory receptor Orco. Nature 2018, 560, 447–452. [Google Scholar] [CrossRef]
- Sato, K.; Pellegrino, M.; Nakagawa, T.; Nakagawa, T.; Vosshall, L.B.; Touhara, K. Insect olfactory receptors are heteromeric ligand-gated ion channels. Nature 2008, 452, 1002–1006. [Google Scholar] [CrossRef]
- Del Mármol, J.; Yedlin, M.A.; Ruta, V. The structural basis of odorant recognition in insect olfactory receptors. Nature 2021, 597, 126–131. [Google Scholar] [CrossRef]
- Terrado, M.; Okon, M.; Mcintosh, L.P.; Plettner, E. Ligand- and pH-Induced Structural Transition of Gypsy Moth Lymantria dispar Pheromone-Binding Protein 1 (LdisPBP1). Biochemistry 2020, 59, 3411–3426. [Google Scholar] [CrossRef]
- Northey, T.; Venthur, H.; De Biasio, F.; Chauviac, F.X.; Cole, A.; Ribeiro, K.A.L.; Grossi, G.; Falabella, P.; Field, L.M.; Keep, N.H.; et al. Crystal structures and binding dynamics of odorant-binding protein 3 from two aphid species Megoura viciae and Nasonovia ribisnigri. Sci. Rep. 2016, 6, 24739. [Google Scholar] [CrossRef] [PubMed]
- Pesenti, M.E.; Spinelli, S.; Bezirard, V.; Briand, L.; Pernollet, J.C.; Tegoni, M.; Cambillau, C. Structural Basis of the Honey Bee PBP Pheromone and pH-induced Conformational Change. J. Mol. Biol. 2008, 380, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Ziemba, B.P.; Murphy, E.J.; Edlin, H.T.; Jones, D.N.M. A novel mechanism of ligand binding and release in the odorant binding protein 20 from the malaria mosquito Anopheles gambiae. Protein Sci. 2013, 22, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campanacci, V.; Lartigue, A.; Hällberg, B.M.; Jones, T.A.; Giudici-Orticoni, M.T.; Tegoni, M.; Cambillau, C. Moth chemosensory protein exhibits drastic conformational changes and cooperativity on ligand binding. Proc. Natl. Acad. Sci. USA 2003, 100, 5069–5074. [Google Scholar] [CrossRef] [Green Version]
- Sandler, B.H.; Nikonova, L.; Leal, W.S.; Clardy, J. Sexual attraction in the silkworm moth: Structure of the pheromone-binding-protein-bombykol complex. Chem. Biol. 2000, 7, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Rihani, K.; Ferveur, J.F.; Briand, L. The 40-year mystery of insect odorant-binding proteins. Biomolecules 2021, 11, 509. [Google Scholar] [CrossRef]
- Larter, N.K.; Sun, J.S.; Carlson, J.R. Organization and function of Drosophila odorant binding proteins. eLife 2016, 5, e20242. [Google Scholar] [CrossRef]
- Pelosi, P.; Zhou, J.J.; Ban, L.P.; Calvello, M. Soluble proteins in insect chemical communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef]
- Birkett, M.A.; Pickett, J.A. Aphid sex pheromones: From discovery to commercial production. Phytochemistry 2003, 62, 651–656. [Google Scholar] [CrossRef]
- Howard, R.W.; Blomquist, G.J. Ecological, Behavoiral, and Biochemical Aspects of Insect Hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef]
- Nordlund, D.A.; Lewis, W.J. Terminology of chemical releasing stimuli in intraspecific and interspecific interactions. J. Chem. Ecol. 1976, 2, 211–220. [Google Scholar] [CrossRef]
- Burger, B.V. Mammalian semiochemicals. In The Chemistry of Pheromones and Other Semiochemicals II; Springer: Berlin/Heidelberg, Germany, 2004; pp. 231–278. [Google Scholar]
- Campagna, S.; Mardon, J.; Celerier, A.; Bonadonna, F. Potential semiochemical molecules from birds: A practical and comprehensive compilation of the last 20 years studies. Chem. Senses 2012, 37, 3–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magurran, A.E.; Irving, P.W.; Henderson, P.A. Is there a fish alarm pheromone? A wild study and critique. Proc. R. Soc. B Biol. Sci. 1996, 263, 1551–1556. [Google Scholar] [CrossRef]
- Pickett, J.A.; Allemann, R.K.; Birkett, M.A. The semiochemistry of aphids. Nat. Prod. Rep. 2013, 30, 1277. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.G.; Silverstein, R.M.; Moser, J.C. Biological Responses of Atta texana to Its Alarm Pheromone and the Enantiomer of the Pheromone. Science 1974, 183, 760–762. [Google Scholar] [CrossRef]
- Marumo, S.; Iwaki, S.; Saito, T.; Yamada, M.; Katagiri, K. Synthesis and Activity of Optically Active Disparlure. J. Am. Chem. Soc. 1974, 21, 7842–7844. [Google Scholar]
- Mori, K. Synthesis of optically active forms of frontalin: The pheromone of dendroctonus bark beetles. Tetrahedron 1975, 31, 1381–1384. [Google Scholar] [CrossRef]
- Tumlinson, J.H.; Klein, M.G.; Doolittle, R.E.; Ladd, T.L.; Proveaux, A.T. Identification of the Female Japanese Beetle Sex Pheromone: Inhibition of Male Response by an Enantiomer. Science 1977, 197, 789–792. [Google Scholar] [CrossRef]
- Haniotakis, G.; Francke, W.; Mori, K.; Redlich, H.; Schurig, V. Sex-specific activity of (R)-(-)- and (S)- (+)-1,7-dioxaspiro[5.5]undecane, the major pheromone of Dacus oleae. J. Chem. Ecol. 1986, 12, 1559–1568. [Google Scholar] [CrossRef]
- Shi, X.; Webster, F.X.; Meinwald, J. Synthesis and the determination of the absolute configuration of matsuone, sex pheromone of female Matsucoccus pine scales. Tetrahedron Lett. 1995, 36, 7201–7204. [Google Scholar] [CrossRef]
- Dawson, G.; Griffiths, D.; Janes, N. Identification of an aphid sex pheromone. Nature 1987, 325, 614–616. [Google Scholar] [CrossRef]
- Marsh, D. Sex pheromone in the aphid Megoura viciae. Nature 1972, 238, 31–32. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.A.M.; Dawson, G.W.; Griffiths, D.C.; Pettersson, J.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Sex attractant pheromone of damson-hop aphid Phorodon humuli (Homoptera, aphididae). J. Chem. Ecol. 1990, 16, 3455–3465. [Google Scholar] [CrossRef]
- Dawson, G.W.; Griffiths, D.C.; Merritt, L.A.; Mudd, A.; Pickett, J.A.; Woodcock, C.M. Aphid semiochemicals—A review, and recent advances on the sex pheromone. J. Chem. Ecol. 1990, 16, 3019–3030. [Google Scholar] [CrossRef] [PubMed]
- Borden, J.H.; Hong, C.L.; McLean, J.A.; Slessor, K.N.; Mori, K. Gnathotrichus sulcatus: Synergistic response to enantiomers of the aggregation pheromone sulcatol. Science 1976, 192, 894–896. [Google Scholar] [CrossRef]
- Bello, J.E.; McElfresh, J.S.; Millar, J.G. Isolation and determination of absolute configurations of insect-produced methyl-branched hydrocarbons. Proc. Natl. Acad. Sci. USA 2015, 112, 1077–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szendrei, Z.; Rodriguez-Saona, C. A meta-analysis of insect pest behavioral manipulation with plant volatiles. Entomol. Exp. Appl. 2010, 134, 201–210. [Google Scholar] [CrossRef]
- De Vega, C.; Herrera, C.M.; Dötterl, S. Floral volatiles play a key role in specialized ant pollination. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Bellesia, F.; Grandi, R.M.; Pagnoni, U.; Pinetti, A.; Trave, R. Biosynthesis of nepetalactone in Nepeta cataria. Phytochemistry 1984, 23, 83–87. [Google Scholar] [CrossRef]
- Lichman, B.R.; Godden, G.T.; Hamilton, J.P.; Palmer, L.; Kamileen, M.O.; Zhao, D.; Vaillancourt, B.; Wood, J.C.; Sun, M.; Kinser, T.J.; et al. The evolutionary origins of the cat attractant nepetalactone in catnip. Sci. Adv. 2020, 6, eaba0721. [Google Scholar] [CrossRef]
- Bakke, A.; Frøyen, P.; Skattebøl, L. Field response to a new pheromonal compound isolated from Ips typographus. Naturwissenschaften 1977, 64, 98–99. [Google Scholar] [CrossRef]
- Erbilgin, N.; Mori, S.R.; Sun, J.H.; Stein, J.D.; Owen, D.R.; Merrill, L.D.; Bolaños, R.C.; Raffa, K.F.; Montiel, T.M.; Wood, D.L.; et al. Response to host volatiles by native and introduced populations of Dendroctonus valens (Coleoptera: Curculionidae, Scolytinae) in North America and China. J. Chem. Ecol. 2007, 33, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Ulland, S.; Ian, E.; Borg-Karlson, A.K.; Mustaparta, H. Discrimination between enantiomers of linalool by olfactory receptor neurons in the cabbage moth Mamestra brassicae (L.). Chem. Senses 2006, 31, 325–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huff, R.M.; Jason Pitts, R. An odorant receptor from Anopheles gambiae that demonstrates enantioselectivity to the plant volatile, linalool. PLoS ONE 2019, 14, e0225637. [Google Scholar] [CrossRef]
- Schneider, D. 100 years of pheromone research—An essay on lepidoptera. Naturwissenschaften 1992, 79, 241–250. [Google Scholar] [CrossRef]
- Hansson, B.S. Olfaction in Lepidoptera. Experientia 1995, 51, 1003–1027. [Google Scholar] [CrossRef]
- Von Butenandt, A.; Beckmann, R.; Stamm, D.; Hecker, E. Über den Sexual-Lockstoff des Seidenspinners Bombyx mori. Reindarstellung Konstitution. Z. Nat. B 1959, 14, 283–284. [Google Scholar]
- Dawson, G.W.; Pickett, J.A.; Smiley, D.W.M. The aphid sex pheromone cyclopentanoids: Synthesis in the elucidation of structure and biosynthetic pathways. Bioorgan. Med. Chem. 1996, 4, 351–361. [Google Scholar] [CrossRef]
- Clyne, P.J.; Warr, C.G.; Freeman, M.R.; Lessing, D.; Kim, J.; Carlson, J.R. A novel family of divergent seven-transmembrane proteins: Candidate odorant receptors in Drosophila. Neuron 1999, 22, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Dobritsa, A.A.; Van Der Goes Van Naters, W.; Warr, C.G.; Steinbrecht, R.A.; Carlson, J.R. Integrating the molecular and cellular basis of odor coding in the Drosophila antenna. Neuron 2003, 37, 827–841. [Google Scholar] [CrossRef] [Green Version]
- De Biasio, F.; Riviello, L.; Bruno, D.; Grimaldi, A.; Congiu, T.; Sun, Y.F.; Falabella, P. Expression pattern analysis of odorant-binding proteins in the pea aphid Acyrthosiphon pisum. Insect Sci. 2015, 22, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Bruno, D.; Grossi, G.; Salvia, R.; Scala, A.; Farina, D.; Grimaldi, A.; Zhou, J.J.; Bufo, A.; Vogel, H.; Grosse-Wilde, E.; et al. Sensilla Morphology and Complex Expression Pattern of Odorant Binding Proteins in the Vetch Aphid Megoura viciae (Hemiptera: Aphididae). Front. Phys. 2018, 9, 777. [Google Scholar] [CrossRef] [Green Version]
- Yuvaraj, J.; Andersson, M.; Anderbant, O.; Löfstedt, C. Diversity of olfactory structures: A comparative study of antennal sensilla in Trichoptera and Lepidoptera. Micron 2018, 111, 9–18. [Google Scholar] [CrossRef]
- Gomez-Diaz, C.; Reina, J.H.; Cambillau, C.; Benton, R. Ligands for Pheromone-Sensing Neurons Are Not Conformationally Activated Odorant Binding Proteins. PLoS Biol. 2013, 11, e1001546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Kristoffersen, L.; Hansson, B.S.; Anderbrant, O.; Larsson, M.C. Aglomerular hemipteran antennal lobes—Basic neuroanatomy of a small nose. Chem. Senses 2008, 33, 771–778. [Google Scholar] [CrossRef] [Green Version]
- Benton, R. On the ORigin of smell: Odorant receptors in insects. Cell. Mol. Life Sci. 2006, 63, 1579–1585. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.; Celniker, S.; Holt, R.; Evans, C.; Gocyane, J.; Amanatides, P.; Scherer, S.; Li, P.; Hoskins, R. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Guo, Z.; Liu, Y.; Zhang, Y. Progress and prospects of CRISPR/Cas systems in insects and other arthropods. Front. Physiol. 2017, 8, 608. [Google Scholar] [CrossRef]
- Pelosi, P.; Maida, R. Odorant-binding proteins in insects. Comp. Biochem. Physiol. 1995, 111, 503–514. [Google Scholar] [CrossRef]
- Zhou, J.J.; Vieira, F.G.; He, X.L.; Smadja, C.; Liu, R.; Rozas, J.; Field, L.M. Genome annotation and comparative analyses of the odorant-binding proteins and chemosensory proteins in the pea aphid Acyrthosiphon pisum. Insect Mol. Biol. 2010, 19, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosbah, A.; Campanacci, V.; Lartigue, A.; Tegoni, M.; Cambillau, C.; Darbon, H. Solution structure of a chemosensory protein from the moth Mamestra brassicae. Biochem. J. 2003, 369, 39–44. [Google Scholar] [CrossRef]
- Leal, W.S.; Chen, A.M.; Ishida, Y.; Chiang, V.P.; Erickson, M.L.; Morgan, T.I.; Tsuruda, J.M. Kinetics and molecular properties of pheromone binding and release. Proc. Natl. Acad. Sci. USA 2005, 102, 5386–5391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowcun, A.; Honson, N.; Plettner, E. Olfaction in the gypsy moth, Lymantria dispar: Effect of pH, ionic strength, and reductants on pheromone transport by pheromone-binding proteins. J. Biol. Chem. 2001, 276, 44770–44776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leite, N.R.; Krogh, R.; Xu, W.; Ishida, Y.; Iulek, J.; Leal, W.S.; Oliva, G. Structure of an Odorant-Binding Protein from the Mosquito Aedes aegypti Suggests a Binding Pocket Covered by a pH-Sensitive “Lid”. PLoS ONE 2009, 4, e8006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubkov, S.; Gronenborn, A.M.; Byeon, I.J.L.; Mohanty, S. Structural consequences of the pH-induced conformational switch in A. polyphemus pheromone-binding protein: Mechanisms of ligand release. J. Mol. Biol. 2005, 354, 1081–1090. [Google Scholar] [CrossRef]
- Wogulis, M.; Morgan, T.; Ishida, Y.; Leal, W.S.; Wilson, D.K. The crystal structure of an odorant binding protein from Anopheles gambiae: Evidence for a common ligand release mechanism. Biochem. Biophys. Res. Commun. 2006, 339, 157–164. [Google Scholar] [CrossRef]
- Han, L.; Zhang, Y.J.; Zhang, L.; Cui, X.; Yu, J.; Zhang, Z.; Liu, M.S. Operating mechanism and molecular dynamics of pheromone-binding protein ASP1 as influenced by pH. PLoS ONE 2014, 9, e110565. [Google Scholar] [CrossRef]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Francesca, R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. 2018, 93, 184–200. [Google Scholar] [CrossRef] [Green Version]
- Leal, W.S. Odorant Reception in Insects: Roles of Receptors, Binding Proteins, and Degrading Enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Tuccori, E.; He, X.; Gazzano, A.; Field, L.; Zhou, J.J.; Pelosi, P. Discrimination of alarm pheromone (E)-β-farnesene by aphid odorant-binding proteins. Insect Biochem. Mol. Biol. 2009, 39, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, P.; He, X.L.; Woodcock, C.; Pickett, J.A.; Field, L.M.; Birkett, M.A.; Kalinova, B.; Gomulski, L.M.; Scolari, F.; Gasperi, G.; et al. Identification of pheromone components and their binding affinity to the odorant binding protein CcapOBP83a-2 of the Mediterranean fruit fly, Ceratitis capitata. Insect Biochem. Mol. Biol. 2014, 48, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.J.; Robertson, G.; He, X.; Dufour, S.; Hooper, A.M.; Pickett, J.A.; Keep, N.H.; Field, L.M. Characterisation of Bombyx mori Odorant-binding Proteins Reveals that a General Odorant-binding Protein Discriminates Between Sex Pheromone Components. J. Mol. Biol. 2009, 389, 529–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Pelosi, P.; Liu, Y.; Lin, K.J.; Yuan, H.B.; Wang, G.R. Ligand-binding properties of three odorant-binding proteins of the diamondback moth Plutella xylostella. J. Integr. Agric. 2016, 15, 580–590. [Google Scholar] [CrossRef] [Green Version]
- Leal, G.M.; Leal, W.S. Binding of a fluorescence reporter and a ligand to an odorant-binding protein of the yellow fever mosquito, Aedes aegypti. F1000Research 2015, 3, 305. [Google Scholar] [CrossRef]
- Zhang, T.T.; Mei, X.D.; Feng, J.N.; Berg, B.G.; Zhang, Y.J.; Guo, Y.Y. Characterization of three pheromone-binding proteins (PBPs) of Helicoverpa armigera (Hübner) and their binding properties. J. Insect Physiol. 2012, 58, 941–948. [Google Scholar] [CrossRef]
- Laughlin, J.D.; Ha, T.S.; Jones, D.N.M.; Smith, D.P. Activation of Pheromone-Sensitive Neurons Is Mediated by Conformational Activation of Pheromone-Binding Protein. Cell 2008, 133, 1255–1265. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Repp, A.; Smith, D.P. LUSH odorant-binding protein mediates chemosensory responses to alcohols in Drosophila melanogaster. Genetics 1998, 150, 711–721. [Google Scholar] [CrossRef]
- Hooper, A.M.; Dufour, S.; He, X.; Muck, A.; Zhou, J.-J.; Almeida, R.; Field, L.M.; Svatos, A.; Pickett, J. High-throughput ESI-MS analysis of binding between the Bombyx mori pheromone-binding protein BmorPBP1, its pheromone components and some analogues. Chem. Commun. 2009, 38, 5725–5727. [Google Scholar] [CrossRef]
- Oldham, N.J.; Krieger, J.; Breer, H.; Fischedick, A.; Hoskovec, M.; Svatos, A. Analysis of the Silkworm Moth Pheromone. Angew. Chemie Int. Ed. 2000, 39, 4341–4343. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, B.; Grossi, G.; Falabella, P.; Liu, Y.; Yan, S.; Lu, J.; Xi, J.; Wang, G. Molecular Basis of Alarm Pheromone Detection in Aphids. Curr. Biol. 2017, 27, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Liu, J.-T.; Zhang, Y.-J.; Chen, J.-L.; Li, X.-C.; Liang, P.; Gao, X.-W.; Zhou, J.-J.; Gu, S.-H. Coordinative mediation of the response to alarm pheromones by three odorant binding proteins in the green peach aphid Myzus persicae. Insect Biochem. Mol. Biol. 2021, 130, 103528. [Google Scholar] [CrossRef]
- Dong, X.-T.; Liao, H.; Zhu, G.-H.; Khuhro, S.A.; Ye, Z.-F.; Yan, Q.; Dong, S.-L. CRISPR/Cas9 mediated PBP1 and PBP3 mutagenesis induced significant reduction in electrophysiological response to sex pheromones in male Chilo suppressalis. Insect Sci. 2017, 26, 388–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.; Liu, Y.; Yang, T.; Pelosi, P.; Dong, S.; Wang, G. Pheromone binding proteins enhance the sensitivity of olfactory receptors to sex pheromones in Chilo suppressalis. Sci. Rep. 2015, 5, 13093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, G.H.; Xu, J.; Cui, Z.; Dong, X.T.; Ye, Z.F.; Niu, D.J.; Huang, Y.P.; Dong, S.L. Functional characterization of SlitPBP3 in Spodoptera litura by CRISPR/Cas9 mediated genome editing. Insect Biochem. Mol. Biol. 2016, 75, 1–9. [Google Scholar] [CrossRef]
- Matsuo, T.; Sugaya, S.; Yasukawa, J.; Aigaki, T.; Fuyama, Y. Odorant-binding proteins OBP57d and OBP57e affect taste perception and host-plant preference in Drosophila sechellia. PLoS Biol. 2007, 5, e118. [Google Scholar] [CrossRef] [Green Version]
- Dani, F.R.; Iovinella, I.; Felicioli, A.; Niccolini, A.; Calvello, M.A.; Carucci, M.G.; Qiao, H.; Pieraccini, G.; Turillazzi, S.; Moneti, G.; et al. Mapping the expression of soluble olfactory proteins in the honeybee. J. Proteome Res. 2010, 9, 1822–1833. [Google Scholar] [CrossRef]
- Wang, S.; Minter, M.; Homem, R.A.; Michaelson, L.V.; Venthur, H.; Lim, K.S.; Withers, A.; Xi, J.; Jones, C.M.; Zhou, J.-J. Odorant binding proteins promote flight activity in the migratory insect, Helicoverpa armigera. Mol. Ecol. 2020, 29, 3795–3808. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Zhu, X.Y.; Fang, L.P.; He, P.; Wang, Z.Q.; Chen, G.; Sun, L.; Ye, Z.F.; Deng, D.G.; Li, J.B. Identification and expression profiles of sex pheromone biosynthesis and transport related genes in Spodoptera litura. PLoS ONE 2015, 10, e0120270. [Google Scholar] [CrossRef]
- Benton, R. Sensitivity and specificity in Drosophila pheromone perception. Trends Neurosci. 2007, 30, 512–519. [Google Scholar] [CrossRef]
- Ban, L.; Napolitano, E.; Serra, A.; Zhou, X.; Iovinella, I.; Pelosi, P. Identification of pheromone-like compounds in male reproductive organs of the oriental locust Locusta migratoria. Biochem. Biophys. Res. Commun. 2013, 437, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Forêt, S.; Wanner, K.W.; Maleszka, R. Chemosensory proteins in the honey bee: Insights from the annotated genome, comparative analyses and expressional profiling. Insect Biochem. Mol. Biol. 2007, 37, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Spehr, M.; Munger, S.D. Olfactory receptors: GPCRs and beyond. J. Neurochem. 2009, 109, 1570–1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenbaum, D.M.; Rasmussen, S.G.; Kobilka, B.K. The structure and function of G-protein-coupled receptors. Nature 2009, 459, 356–363. [Google Scholar] [CrossRef] [Green Version]
- Wicher, D.; Schafer, R.; Bauernfeind, R.; Stensmyr, M.C.; Heller, R.; Heinemann, S.H.; Hansson, B.S. Drosophila odorant receptors are both ligand-gated and cyclic-nucleotide-activated cation channels. Nature 2008, 452, 1007–1011. [Google Scholar] [CrossRef] [PubMed]
- Larsson, M.C.; Domingos, A.I.; Jones, W.D.; Chiappe, M.E.; Amrein, H.; Vosshall, L.B. Or83b encodes a broadly expressed odorant receptor essential for Drosophila olfaction. Neuron 2004, 43, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Brand, P.; Robertson, H.M.; Lin, W.; Pothula, R.; Klingeman, W.E.; Jurat-Fuentes, J.L.; Johnson, B.R. The origin of the odorant receptor gene family in insects. eLife 2018, 7, e38340. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.; Chen, D.; Yang, P.; Jiang, F.; Wang, X.; Kang, L. CRISPR/Cas9 in locusts: Successful establishment of an olfactory deficiency line by targeting the mutagenesis of an odorant receptor co-receptor (Orco). Insect Biochem. Mol. Biol. 2016, 79, 27–35. [Google Scholar] [CrossRef]
- Fandino, R.A.; Haverkamp, A.; Bisch-Knaden, S.; Zhang, J.; Bucks, S.; Nguyen, T.A.T.; Schröder, K.; Werckenthin, A.; Rybak, J.; Stengl, M.; et al. Mutagenesis of odorant coreceptor Orco fully disrupts foraging but not oviposition behaviors in the hawkmoth Manduca sexta. Proc. Natl. Acad. Sci. USA 2019, 116, 15677–15685. [Google Scholar] [CrossRef] [Green Version]
- Trible, W.; Olivos-Cisneros, L.; McKenzie, S.K.; Saragosti, J.; Chang, N.C.; Matthews, B.J.; Oxley, P.R.; Kronauer, D.J.C. Orco Mutagenesis Causes Loss of Antennal Lobe Glomeruli and Impaired Social Behavior in Ants. Cell 2017, 170, 727–735. [Google Scholar] [CrossRef]
- Koutroumpa, F.; Monsempès, C.; Anton, S.; François, M.C.; Montagné, N.; Jacquin-Joly, E. Pheromone Receptor Knock-Out Affects Pheromone Detection and Brain Structure in a Moth. Biomolecules 2022, 12, 341. [Google Scholar] [CrossRef] [PubMed]
- Batra, S.; Corcoran, J.; Zhang, D.D.; Pal, P.; Umesh, K.P.; Kulkarni, R.; Löfstedt, C.; Sowdhamini, R.; Olsson, S.B. A functional agonist of insect olfactory receptors: Behavior, physiology and structure. Front. Cell. Neurosci. 2019, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Yuvaraj, J.; Roberts, R.; Sonntag, Y.; Hou, X.; Grosse-Wilde, E.; Machara, A.; Hansson, B.; Johanson, U.; Löfstedt, C.; Andersson, M. Putative ligand binding sites of two functionally characterized bark beetle odorant receptors. BMC Biol. 2021, 19, 16. [Google Scholar] [CrossRef] [PubMed]
- Radadiya, A.; Pickett, J.A. Characterizing human odorant signals: Insights from insect semiochemistry and in silico modelling. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190263. [Google Scholar] [CrossRef] [Green Version]
- Field, L.M.; Pickett, J.A.; Wadhams, L.J. Molecular studies in insect olfaction. Insect Mol. Biol. 2000, 9, 545–551. [Google Scholar] [CrossRef]
- Zaremska, V.; Tan, J.; Lim, S.; Knoll, W.; Pelosi, P. Isoleucine Residues Determine Chiral Discrimination of Odorant-Binding Protein. Chem. Eur. J. 2020, 26, 8720–8724. [Google Scholar] [CrossRef]
- Damberger, F.; Horst, R.; Wüthrich, K.; Peng, G.; Nikonova, L.; Leal, W.S. NMR characterization of a pH-dependent equilibrium between two folded solution conformations of the pheromone-binding protein from Bombyx mori. Protein Sci. 2000, 9, 1038–1041. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Plettner, E. Enantiomer and conformer recognition of (+) and (−)-disparlure and their analogs by the pheromone binding proteins of the gypsy moth, Lymantria dispar. Bioorgan. Med. Chem. 2013, 21, 1811–1822. [Google Scholar] [CrossRef]
- Wojtasek, H.; Hansson, B.S.; Leal, W.S. Attracted or repelled?—A matter of two neurons, one pheromone binding protein, and a chiral center. Biochem. Biophys. Res. Commun. 1998, 250, 217–222. [Google Scholar] [CrossRef]
- Plettner, E.; Lazar, J.; Prestwich, E.G.; Prestwich, G.D. Discrimination of pheromone enantiomers by two pheromone binding proteins from the gypsy moth Lymantria dispar. Biochemistry 2000, 39, 8953–8962. [Google Scholar] [CrossRef]
- Drakou, C.E.; Tsitsanou, K.E.; Potamitis, C.; Fessas, D.; Zervou, M.; Zographos, S.E. The crystal structure of the AgamOBP1•Icaridin complex reveals alternative binding modes and stereo-selective repellent recognition. Cell. Mol. Life Sci. 2016, 74, 319–338. [Google Scholar] [CrossRef] [PubMed]
- Lartigue, A.; Gruez, A.; Spinelli, S.; Rivière, S.; Brossut, R.; Tegoni, M.; Cambillau, C. The crystal structure of a cockroach pheromone-binding protein suggests a new ligand binding and release mechanism. J. Biol. Chem. 2003, 278, 30213–30218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geithe, C.; Protze, J.; Kreuchwig, F.; Krause, G.; Krautwurst, D. Structural determinants of a conserved enantiomer-selective carvone binding pocket in the human odorant receptor OR1A1. Cell. Mol. Life Sci. 2017, 74, 4209–4229. [Google Scholar] [CrossRef] [PubMed]
- Takai, Y.; Touhara, K. Enantioselective recognition of menthol by mouse odorant receptors. Biosci. Biotechnol. Biochem. 2015, 79, 1980–1986. [Google Scholar] [CrossRef] [Green Version]
- Bohbot, J.D.; Dickens, J.C. Selectivity of odorant receptors in insects. Front. Cell. Neurosci. 2012, 6, 2010–2013. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.D.; Löfstedt, C. Moth pheromone receptors: Gene sequences, function, and evolution. Front. Ecol. Evol. 2015, 3, 105. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.B.; Liu, Y.; Yan, S.C.; Wang, G.R. Identification and functional characterization of an odorant receptor in pea aphid, Acyrthosiphon pisum. Insect Sci. 2019, 26, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Malnic, B.; Hirono, J.; Sato, T.; Buck, L.B. Combinatorial receptor codes for odors. Cell 1999, 96, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Seki, Y.; Dweck, H.K.M.; Rybak, J.; Wicher, D.; Sachse, S.; Hansson, B.S. Olfactory coding from the periphery to higher brain centers in the Drosophila brain. BMC Biol. 2017, 15, 18. [Google Scholar] [CrossRef]
- Pask, G.M.; Slone, J.D.; Millar, J.G.; Das, P.; Moreira, J.A.; Zhou, X.; Bello, J.; Berger, S.L.; Bonasio, R.; Desplan, C.; et al. Specialized odorant receptors in social insects that detect cuticular hydrocarbon cues and candidate pheromones. Nat. Commun. 2017, 8, 297. [Google Scholar] [CrossRef]
- Hill, S.R.; Majeed, S.; Ignell, R. Molecular basis for odorant receptor tuning: A short C-terminal sequence is necessary and sufficient for selectivity of mosquito Or8. Insect Mol. Biol. 2015, 24, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, M.; Steinbach, N.; Stensmyr, M.C.; Hansson, B.S.; Vosshall, L.B. A natural polymorphism alters odour and DEET sensitivity in an insect odorant receptor. Nature 2011, 478, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.N.; Taylor, R.W.; Pask, G.M.; Zwiebel, L.J.; Newcomb, R.D.; Christie, D.L. A Conserved Aspartic Acid Is Important for Agonist (VUAA1) and Odorant/Tuning Receptor-Dependent Activation of the Insect Odorant Co-Receptor (Orco). PLoS ONE 2013, 8, e70218. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Leal, W.S. Probing insect odorant receptors with their cognate ligands: Insights into structural features. Biochem. Biophys. Res. Commun. 2013, 435, 477–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, A.S.; Luetje, C.W. Transmembrane segment 3 of Drosophila melanogaster odorant receptor subunit 85b contributes to ligand-receptor interactions. J. Biol. Chem. 2010, 285, 11854–11862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, T.; Pellegrino, M.; Sato, K.; Vosshall, L.B.; Touhara, K. Amino acid residues contributing to function of the heteromeric insect olfactory receptor complex. PLoS ONE 2012, 7, e32372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohbot, J.D.; Dickens, J.C. Characterization of an enantioselective odorant receptor in the yellow fever mosquito Aedes aegypti. PLoS ONE 2009, 4, e7032. [Google Scholar] [CrossRef] [Green Version]
- Dickens, J.C.; Bohbot, J.D. Mini review: Mode of action of mosquito repellents. Pestic. Biochem. Physiol. 2013, 106, 149–155. [Google Scholar] [CrossRef]
- Dekel, A.; Pitts, R.J.; Yakir, E.; Bohbot, J.D. Evolutionarily conserved odorant receptor function questions ecological context of octenol role in mosquitoes. Sci. Rep. 2016, 6, 37330. [Google Scholar] [CrossRef] [Green Version]
- Bohbot, J.D.; Pitts, R.J.; Kwon, H.-W.; Rutzler, M.; Robertson, H.M.; Zwiebel, L.J. Molecular characterization of the Aedes aegypti odorant receptor gene family. Insect Mol. Biol. 2007, 16, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.A.; Fox, A.N.; Pitts, R.J.; Kent, L.B.; Tan, P.L.; Chrystal, M.A.; Cravchik, A.; Collins, F.H.; Robertson, H.M.; Zwiebel, L.J. G protein-coupled receptors in Anopheles gambiae. Science 2002, 298, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.F.; Hughes, D.T.; Luetje, C.W.; Millar, J.G.; Soriano-Agatón, F.; Hanks, L.M.; Robertson, H.M. Sequencing and characterizing odorant receptors of the cerambycid beetle Megacyllene caryae. Insect Biochem. Mol. Biol. 2012, 42, 499–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kythreoti, G.; Sdralia, N.; Tsitoura, P.; Papachristos, D.P.; Michaelakis, A.; Karras, V.; Ruel, D.M.; Yakir, E.; Bohbot, J.D.; Schulz, S.; et al. Volatile allosteric antagonists of mosquito odorant receptors inhibit human-host attraction. J. Biol. Chem. 2021, 296, 100172. [Google Scholar] [CrossRef]
- Chapman, O.L.; Mattes, K.C.; Sheridan, R.S.; Klun, J.A. Stereochemical Evidence of Dual Chemoreceptors for an Achiral Sex Pheromone in Lepidoptera1. J. Am. Chem. Soc. 1978, 100, 4878–4884. [Google Scholar] [CrossRef]
- Yang, K.; Huang, L.Q.; Ning, C.; Wang, C.Z. Two single-point mutations shift the ligand selectivity of a pheromone receptor between two closely related moth species. eLife 2017, 6, e29100. [Google Scholar] [CrossRef]
- Leary, G.P.; Allen, J.E.; Bunger, P.L.; Luginbill, J.B.; Linn, C.E.; Macallister, I.E.; Kavanaugh, M.P.; Wanner, K.W. Single mutation to a sex pheromone receptor provides adaptive specificity between closely related moth species. Proc. Natl. Acad. Sci. USA 2012, 109, 14081–14086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, D.T.; Wang, G.; Zwiebel, L.J.; Luetje, C.W. A determinant of odorant specificity is located at the extracellular loop 2-transmembrane domain 4 interface of an anopheles gambiae odorant receptor subunit. Chem. Senses 2014, 39, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Sargsyan, V.; Getahun, M.N.; Llanos, S.L.; Olsson, S.B.; Hansson, B.S.; Wicher, D. Phosphorylation via PKC regulates the function of the Drosophila odorant co-receptor. Front. Cell. Neurosci. 2011, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hopf, T.A.; Morinaga, S.; Ihara, S.; Touhara, K.; Marks, D.S.; Benton, R. Amino acid coevolution reveals three-dimensional structure and functional domains of insect odorant receptors. Nat. Commun. 2015, 6, 6077. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Ha, T.S.; Smith, D.P. SNMP is a signaling component required for pheromone sensitivity in Drosophila. Proc. Natl. Acad. Sci. USA 2008, 105, 10996–11001. [Google Scholar] [CrossRef] [Green Version]
- Ray, A.; van Naters, W.G.; Carlson, J.R. Molecular determinants of odorant receptor function in insects. J. Biosci. 2014, 39, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Turner, R.M.; Derryberry, S.L.; Kumar, B.N.; Brittain, T.; Zwiebel, L.J.; Newcomb, R.D.; Christie, D.L. Mutational analysis of cysteine residues of the insect odorant co-receptor (Orco) from Drosophila melanogaster reveals differential effects on agonist- and odorant-tuning receptor-dependent activation. J. Biol. Chem. 2014, 289, 31837–31845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukunda, L.; Miazzi, F.; Kaltofen, S.; Hansson, B.S.; Wicher, D. Calmodulin modulates insect odorant receptor function. Cell Calcium 2014, 55, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Antony, B.; Johny, J.; Montagné, N.; Jacquin-Joly, E.; Capoduro, R.; Cali, K.; Persaud, K.; Al-Saleh, M.A.; Pain, A. Pheromone receptor of the globally invasive quarantine pest of the palm tree, the red palm weevil (Rhynchophorus ferrugineus). Mol. Ecol. 2021, 30, 2025–2039. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Leal, W.S. Chiral discrimination of the Japanese beetle sex pheromone and a behavioral antagonist by a pheromone-degrading enzyme. Proc. Natl. Acad. Sci. USA 2008, 105, 9076–9080. [Google Scholar] [CrossRef] [Green Version]
- Wicher, D.; Miazzi, F. Functional properties of insect olfactory receptors: Ionotropic receptors and odorant receptors. Cell Tissue Res. 2021, 383, 7–19. [Google Scholar] [CrossRef]
- Croset, V.; Schleyer, M.; Arguello, J.R.; Gerber, B.; Benton, R. A molecular and neuronal basis for amino acid sensing in the Drosophila larva. Sci. Rep. 2016, 6, 34871. [Google Scholar] [CrossRef]
- Pelosi, P.; Mastrogiacomo, R.; Iovinella, I.; Tuccori, E.; Persaud, K.C. Structure and biotechnological applications of odorant-binding proteins. Appl. Microbiol. Biotechnol. 2014, 98, 61–70. [Google Scholar] [CrossRef]
- Wang, S.; Duan, H.; Yang, X.; Sun, Y.F.; Du, S.; Qin, Y.; Duan, H.; Yang, X. Computer-aided rational design of novel EBF analogues with an aromatic ring. J. Mol. Model. 2016, 22, 144. [Google Scholar] [CrossRef]
- Scorsone, E.; Manai, R.; Cali, K.; Ricatti, M.J.; Farno, S.; Persaud, K.; Mucignat, C. Biosensor array based on ligand binding proteins for narcotics and explosives detection. Sensors Actuators B Chem. 2021, 334, 129587. [Google Scholar] [CrossRef]
Figure 1.
Early synthesized enantiomerically pure pheromones, Atta texana alarm pheromone (S)-4-methyl-3-heptanone 1, Lymantria dispar sex pheromone (7R,8S)-(+)-disparlure 2 and Dendroctonus brevicomis aggregation pheromone (1S,5R)-frontalin 3 .
Figure 1.
Early synthesized enantiomerically pure pheromones, Atta texana alarm pheromone (S)-4-methyl-3-heptanone 1, Lymantria dispar sex pheromone (7R,8S)-(+)-disparlure 2 and Dendroctonus brevicomis aggregation pheromone (1S,5R)-frontalin 3 .

Figure 2.
The chemical structures of a selection of chiral insect pheromones and semiochemicals involved in insect olfaction.(R)-Japonilure 4, (S)-japonilure 5, (R)-malic acid 6, (S)-malic acid 7, (6R,10R)-matsuone 8, (6R,10S)-matsuone 9, (1R,4aS,7S,7aR)-nepetalactol 10, (4aS,7S,7aR)-nepetalactone 11, (1R,2S,5S)-dolichodial 12, (1S,4aR,7S,7aS)-nepetalactol 13 and (1R,4aR,7S,7aS)-nepetalactol 14, (R)-sulcatol 15, (S)-sulcatol 16, α-pinene 17, (R)-linalool 18 and (S)-linalool 19.
Figure 2.
The chemical structures of a selection of chiral insect pheromones and semiochemicals involved in insect olfaction.(R)-Japonilure 4, (S)-japonilure 5, (R)-malic acid 6, (S)-malic acid 7, (6R,10R)-matsuone 8, (6R,10S)-matsuone 9, (1R,4aS,7S,7aR)-nepetalactol 10, (4aS,7S,7aR)-nepetalactone 11, (1R,2S,5S)-dolichodial 12, (1S,4aR,7S,7aS)-nepetalactol 13 and (1R,4aR,7S,7aS)-nepetalactol 14, (R)-sulcatol 15, (S)-sulcatol 16, α-pinene 17, (R)-linalool 18 and (S)-linalool 19.

Figure 3.
The chemical structure of Bombykol 20, the first insect pheromone isolated from the silkworm moth, Bombyx mori.
Figure 3.
The chemical structure of Bombykol 20, the first insect pheromone isolated from the silkworm moth, Bombyx mori.

Figure 4.
The chemical structure of (Z)-vaccenyl acetate (VA) 21, the male sex pheromone of D. melanogaster, and (E)-β-farnesene 22, the aphid alarm pheromone.
Figure 4.
The chemical structure of (Z)-vaccenyl acetate (VA) 21, the male sex pheromone of D. melanogaster, and (E)-β-farnesene 22, the aphid alarm pheromone.

Figure 5.
General olfactory processing in insects. Within the antennal sensilla, odorant-binding proteins (OBPs) play a role in allowing odorants to activate odorant receptors (ORs) which are co- expressed with the olfactory receptor co-receptor (ORCO). Once activated, an action potential travels along the odorant receptor neuron (ORN) to the antennal lobe.
Figure 5.
General olfactory processing in insects. Within the antennal sensilla, odorant-binding proteins (OBPs) play a role in allowing odorants to activate odorant receptors (ORs) which are co- expressed with the olfactory receptor co-receptor (ORCO). Once activated, an action potential travels along the odorant receptor neuron (ORN) to the antennal lobe.

Figure 6.
The chemical structures of (7R,8S)-(+)-disparlure 2 and (7S,8R)-(−)-disparlure 23.

Figure 7.
The chemical structure of 4-ethylacetophenone 24, salicylaldehyde 25 and (S)-cis-verbenol 26, compounds shown to elicit a response to the broadly tuned OR ApisOR4 from A. pisum.
Figure 7.
The chemical structure of 4-ethylacetophenone 24, salicylaldehyde 25 and (S)-cis-verbenol 26, compounds shown to elicit a response to the broadly tuned OR ApisOR4 from A. pisum.

Figure 8.
The chemical structures of A. aegypti kairomone (R)-1-octen-3-ol 27 and (S)-1-octen-3-ol 28, the pheromone components (S)-ipsenol 29, (R)-ipsenol 30, (R)-ipsdienol 31 and (S)-ipsdienol 32 of the Eurasian spruce bark beetle, Ips typographus and the pheromone components (S)-2-methyl-1-butanol 33, (R)-2-methyl-1-butanol 34, (2R,3R)-2,3,-hexanediol 35, (2R,3S)-2,3-hexanediol 36, (2S,3R)-2,3-hexandiol 37 and (2S,3S)-2,3-hexandiol 38 of the cerambycid beetle Megacyllene caryae.
Figure 8.
The chemical structures of A. aegypti kairomone (R)-1-octen-3-ol 27 and (S)-1-octen-3-ol 28, the pheromone components (S)-ipsenol 29, (R)-ipsenol 30, (R)-ipsdienol 31 and (S)-ipsdienol 32 of the Eurasian spruce bark beetle, Ips typographus and the pheromone components (S)-2-methyl-1-butanol 33, (R)-2-methyl-1-butanol 34, (2R,3R)-2,3,-hexanediol 35, (2R,3S)-2,3-hexanediol 36, (2S,3R)-2,3-hexandiol 37 and (2S,3S)-2,3-hexandiol 38 of the cerambycid beetle Megacyllene caryae.

Figure 9.
Topology of an ORCO subunit with key loci for activity highlighted [1,7,9,104,124,126,127,136–142]. Not included are transmembrane domains 2,4,5 and 6, extracellular loop 3 and intracellular loop 3, which all possess additional roles in affecting sensitivity to ligands, ion selectivity and response ratios [9,96,123,127,138,143].
Figure 9.
Topology of an ORCO subunit with key loci for activity highlighted [1,7,9,104,124,126,127,136–142]. Not included are transmembrane domains 2,4,5 and 6, extracellular loop 3 and intracellular loop 3, which all possess additional roles in affecting sensitivity to ligands, ion selectivity and response ratios [9,96,123,127,138,143].


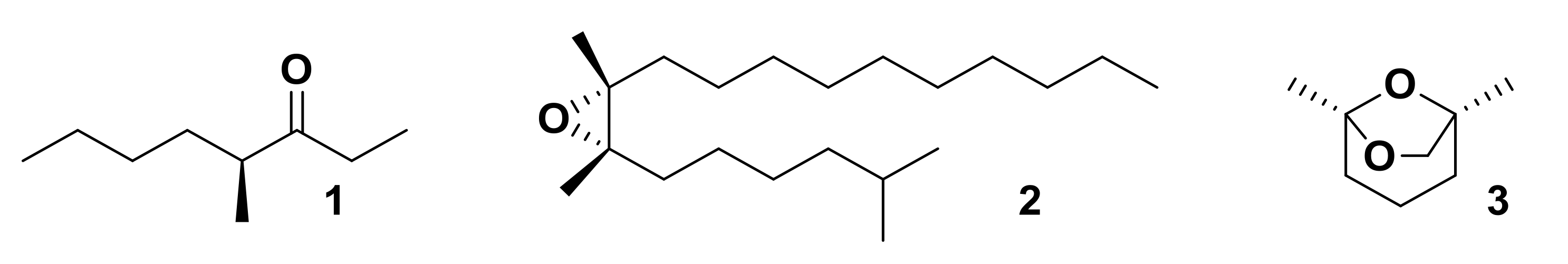{kind=link}
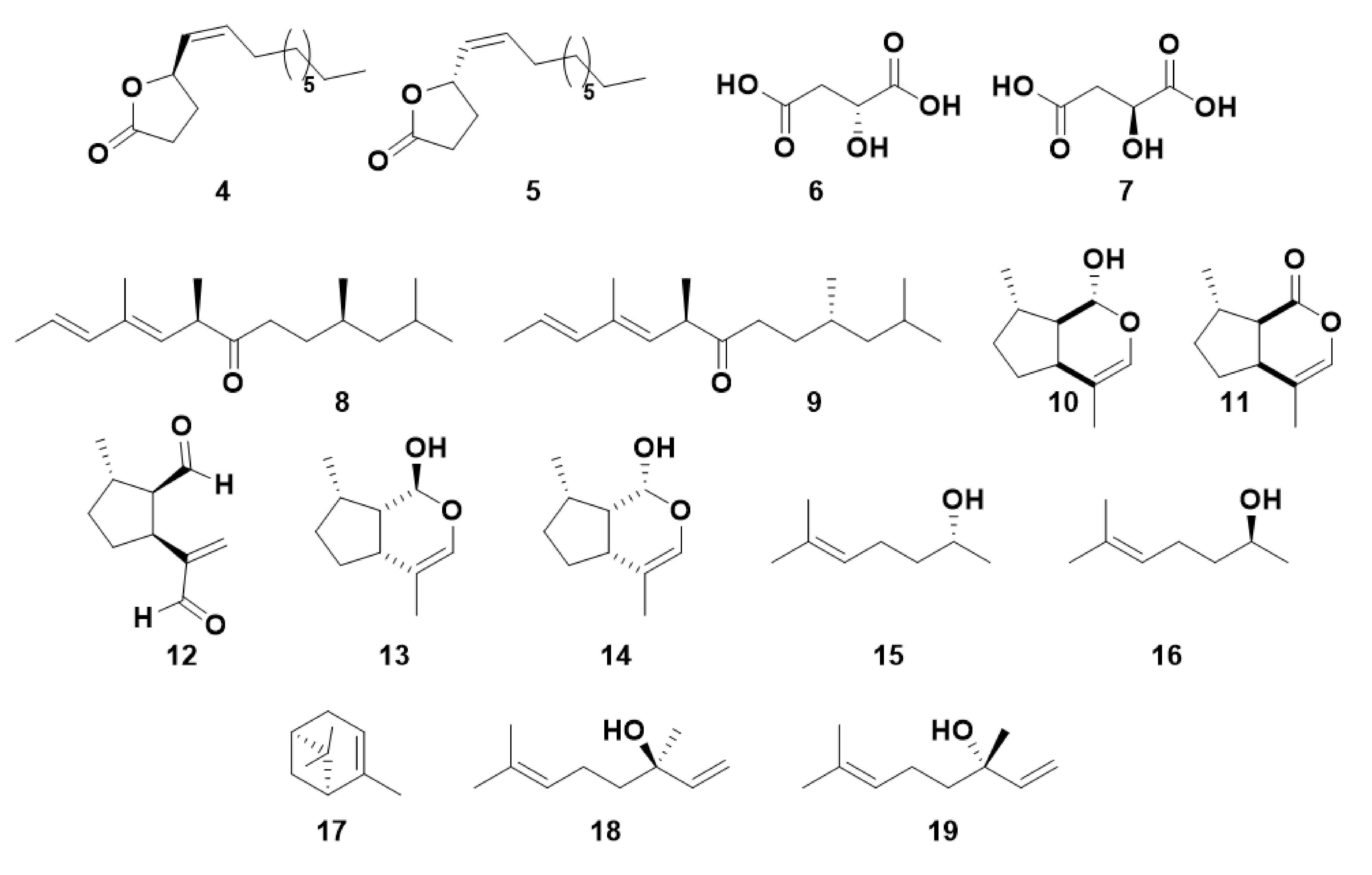{kind=link}
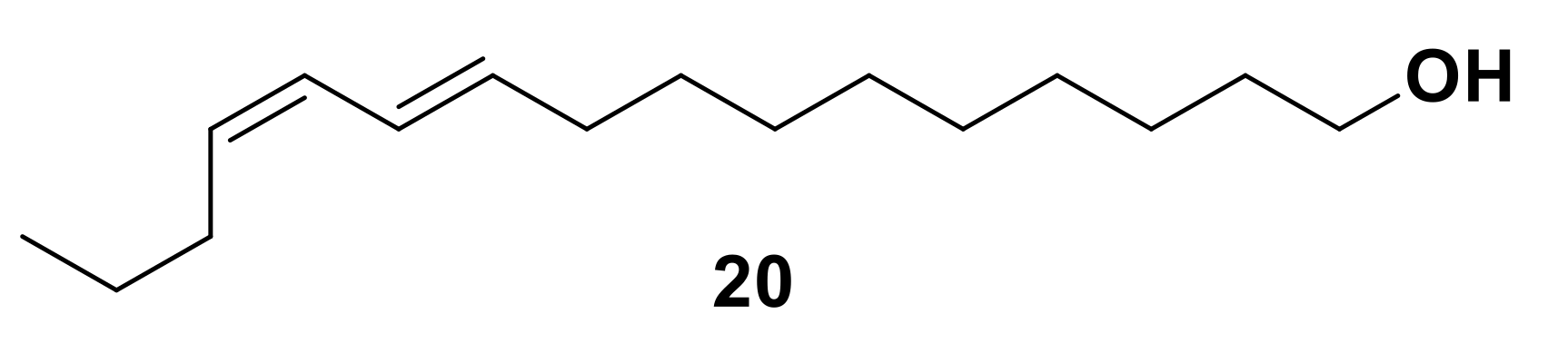{kind=link}
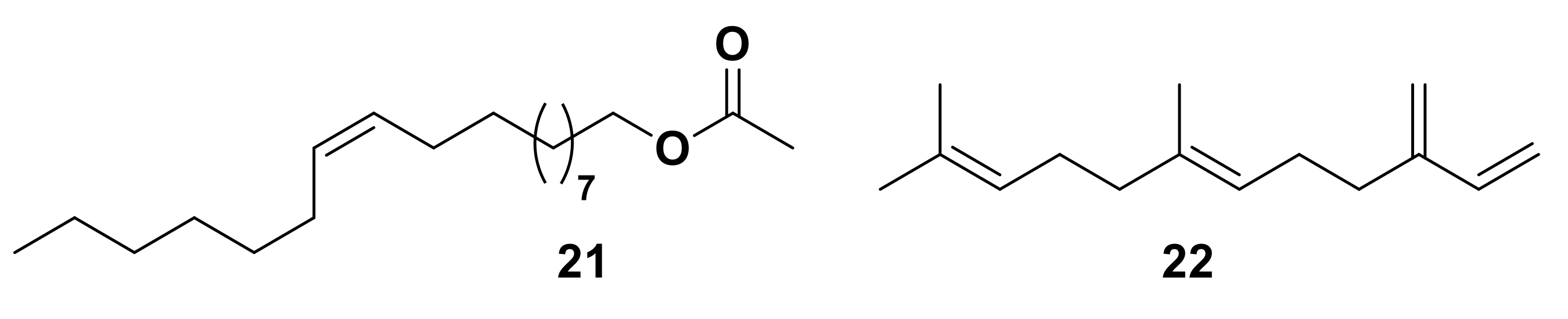{kind=link}
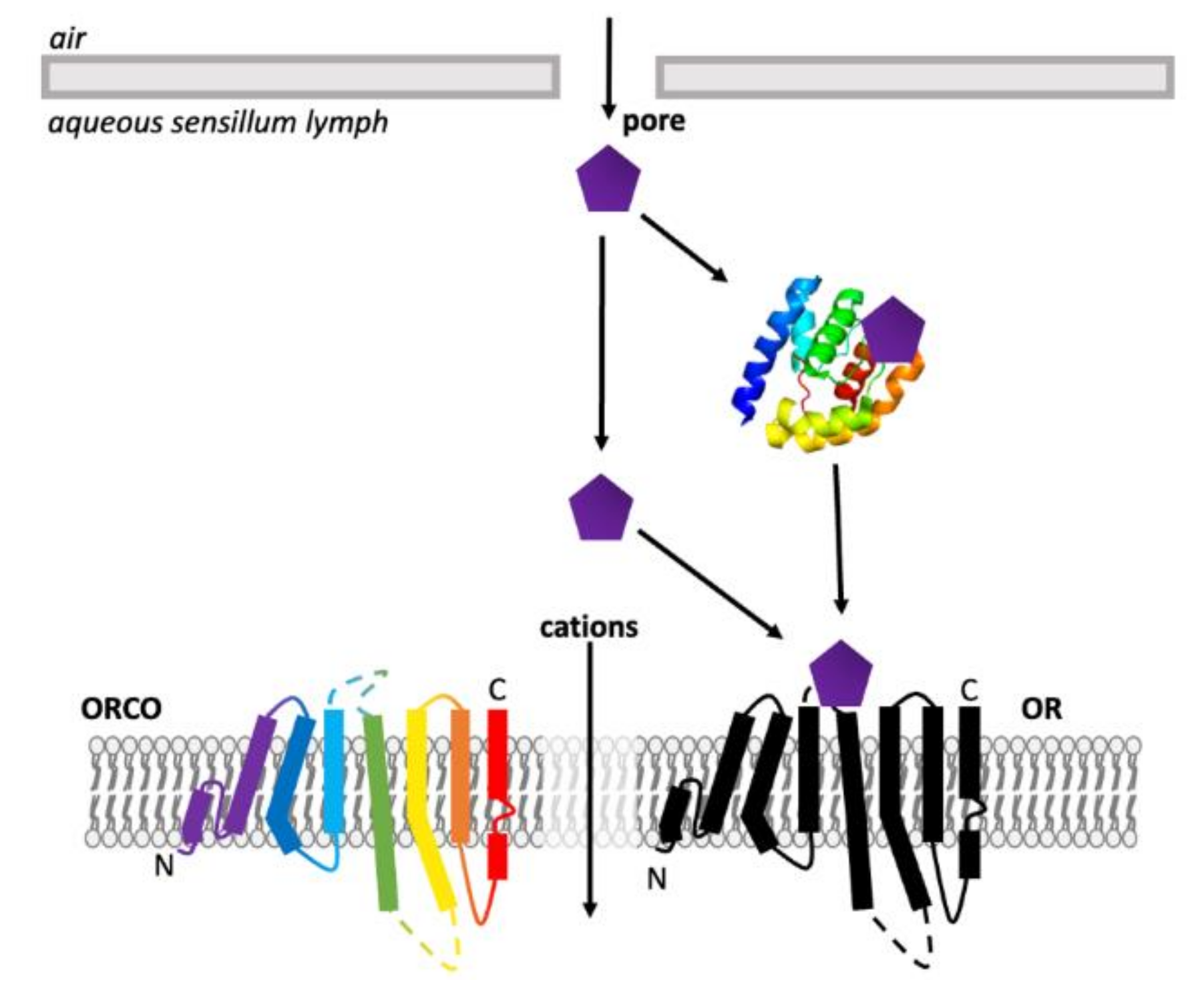{kind=link}
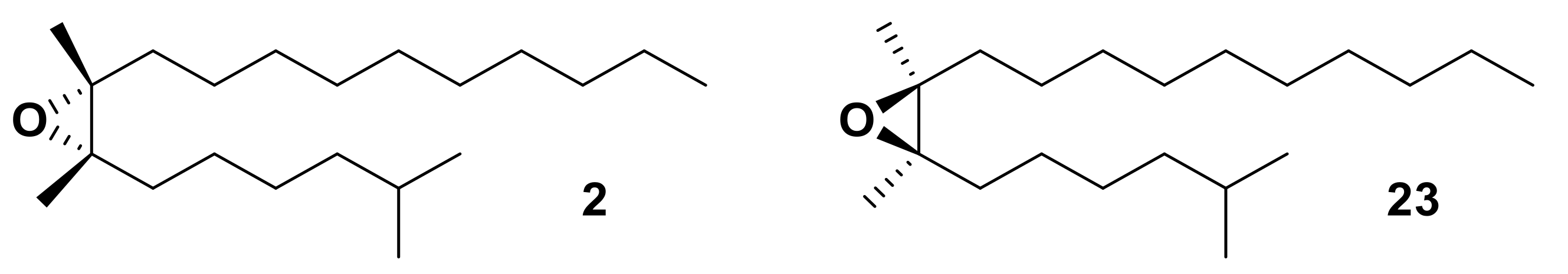{kind=link}
{kind=link}
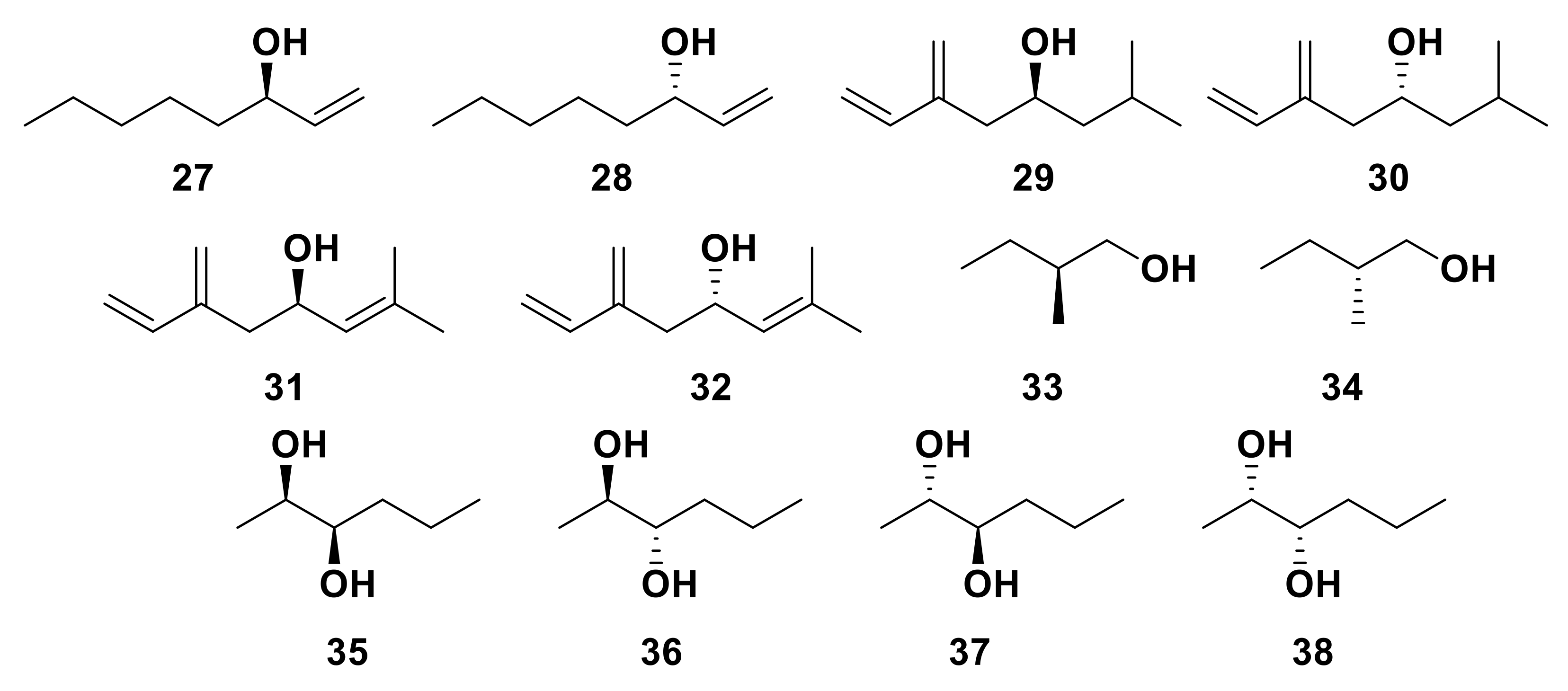{kind=link}
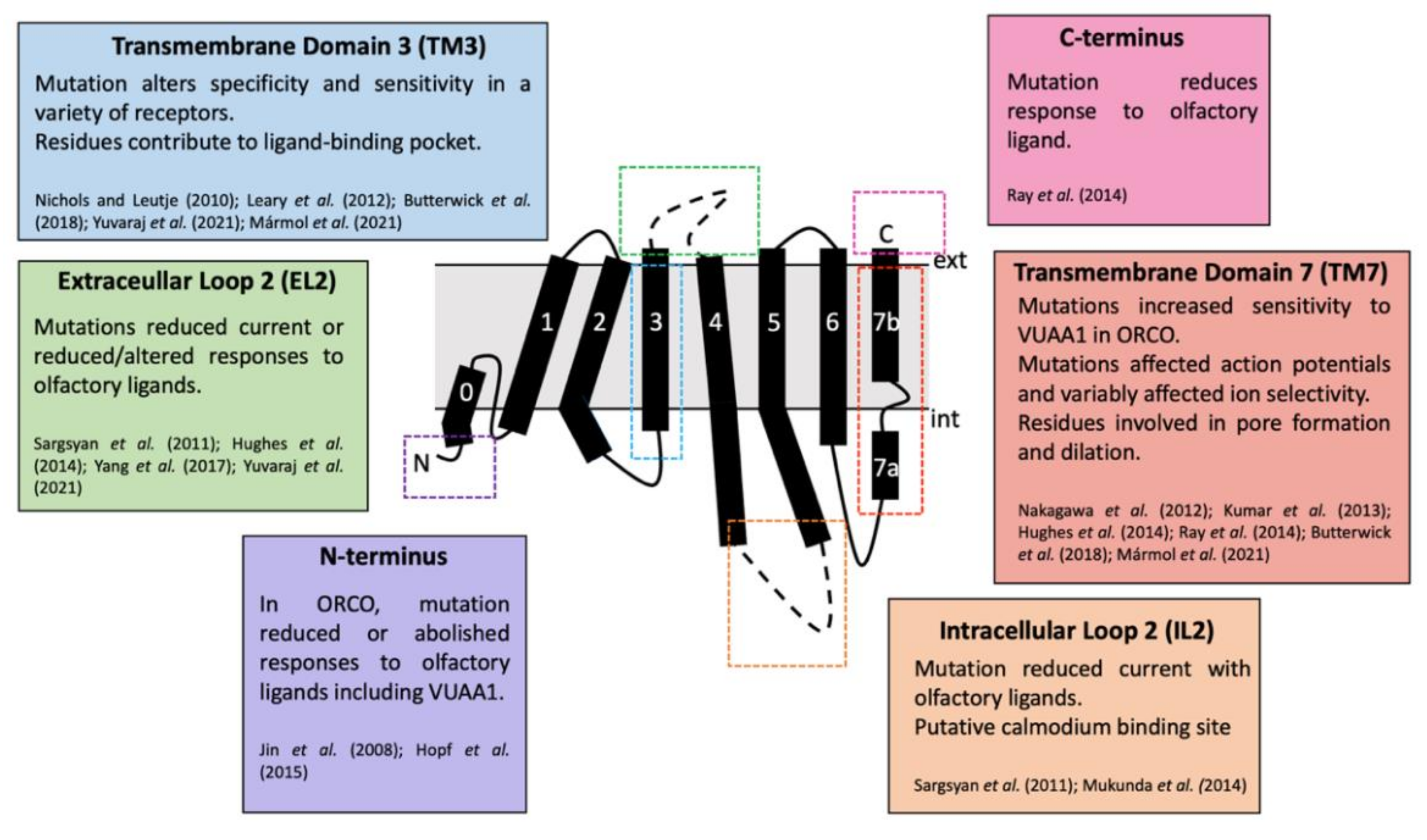{kind=link}
Table 1.
Insect odorant receptors that possess enantioselectivity experimentally. The suggested active enantiomer is highlighted in bold for each receptor.
Table 1.
Insect odorant receptors that possess enantioselectivity experimentally. The suggested active enantiomer is highlighted in bold for each receptor.
| Species | Common Name | Receptor | Ligands | Reference |
|---|---|---|---|---|
| Aedes aegypti | Yellow Fever Mosquito | AaegOR8 | (R)-1-Octen-3-ol (S)-1-Octen-3-ol | Bohbot and Dickens, 2009 [128] |
| Culex quinquefasciatus | Southern House Mosquito | CquiOR8 | Hill et al., 2015 [122] | |
| Toxorhynchites amboinensis | Elephant Mosquito | TambOR8 | Dekel et al., 2016 [130] | |
| Anopheles gambiae | Malaria Vector Mosquito | AgamOR29 | (R)-Linalool (S)-Linalool | Huff and Pitts, 2019 [45] |
| Megacyllene caryae | Hickory Borer | McarOR20 | (2S,3R)-2,3-Hexanediol (2R,3S)-2,3-Hexanediol (2S,3S)-2,3-Hexanediol | Mitchell et al., 2013 [133] |
| Ips typographus | Eurasian spruce Bark Beetle | ItypOR46 | (R)-Ipsenol (S)-Ipsenol | Yuvaraj et al., 2021 [104] |
| ItypOR49 | (R)-Ipsendiol (S)-Ipsendiol |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sims, C.; Birkett, M.A.; Withall, D.M. Enantiomeric Discrimination in Insects: The Role of OBPs and ORs. Insects 2022, 13, 368. https://doi.org/10.3390/insects13040368
AMA Style
Sims C, Birkett MA, Withall DM. Enantiomeric Discrimination in Insects: The Role of OBPs and ORs. Insects. 2022; 13(4):368. https://doi.org/10.3390/insects13040368
Chicago/Turabian StyleSims, Cassie, Michael A. Birkett, and David M. Withall. 2022. "Enantiomeric Discrimination in Insects: The Role of OBPs and ORs" Insects 13, no. 4: 368. https://doi.org/10.3390/insects13040368
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.