
The Addition of Sterols and Cryoprotectants to Optimize a Diet Developed for Eldana saccharina Walker (Lepidoptera: Pyralidae) Using the Carcass Milling Technique


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Eldana saccharina Rearing
2.2.1. Formulation for Sterol Incorporation into Diets
2.2.2. Formulation for Cryoprotectant Incorporation into Diets
2.2.3. Diet Preparation
2.2.4. Diet Dispensing
2.2.5. Inoculation of Neonate Larvae onto the Diet
2.3. Quality Assessment
2.3.1. Development Time to First Pupation
2.3.2. Pupal Harvesting and Pupal Weighing
2.3.3. Adult Emergence and Sex Ratio
2.3.4. Chill Coma Recovery Time
2.4. Oviposition Testing
2.4.1. Male and Female Mating Frequency
2.4.2. Female Fecundity and Fertility
2.5. Statistical Analysis
3. Results
3.1. Insect Production during Quality Control and at Harvest
Diet Incorporation of Sterols
3.2. Population Age Distribution
Diet Incorporation of Sterols
3.3. Male and Female Pupal Weight
3.3.1. Diet Incorporation of Sterols
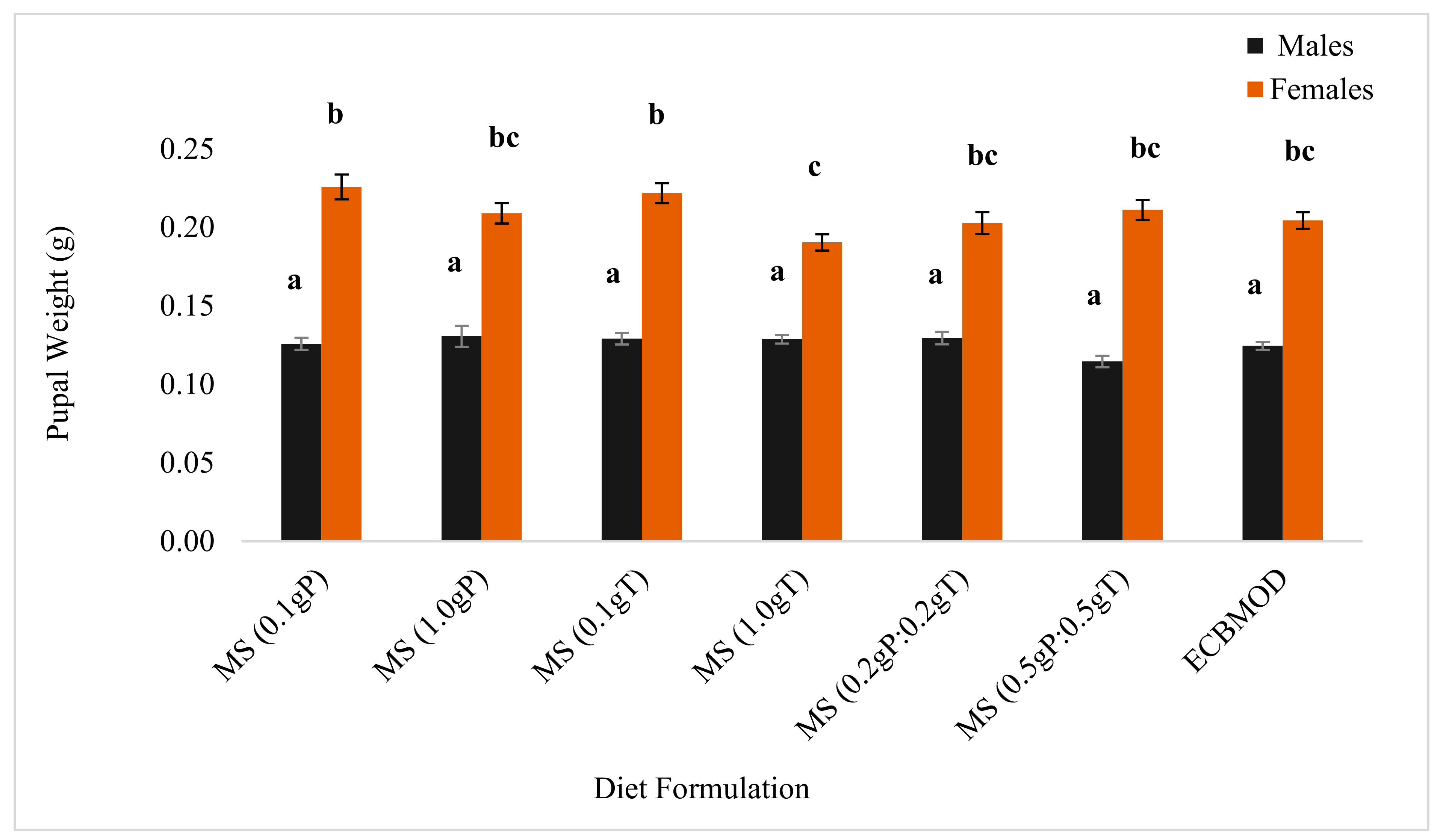
3.3.2. Diet Incorporation of Cryoprotectants
3.3.3. Evaluation of Eldana saccharina Performance between Diets
3.4. Moth Emergence and Sex Ratio
3.4.1. Diet Incorporation of Sterols
3.4.2. Diet Incorporation of Cryoprotectants
3.4.3. Evaluation of Eldana saccharina Performance between Diets
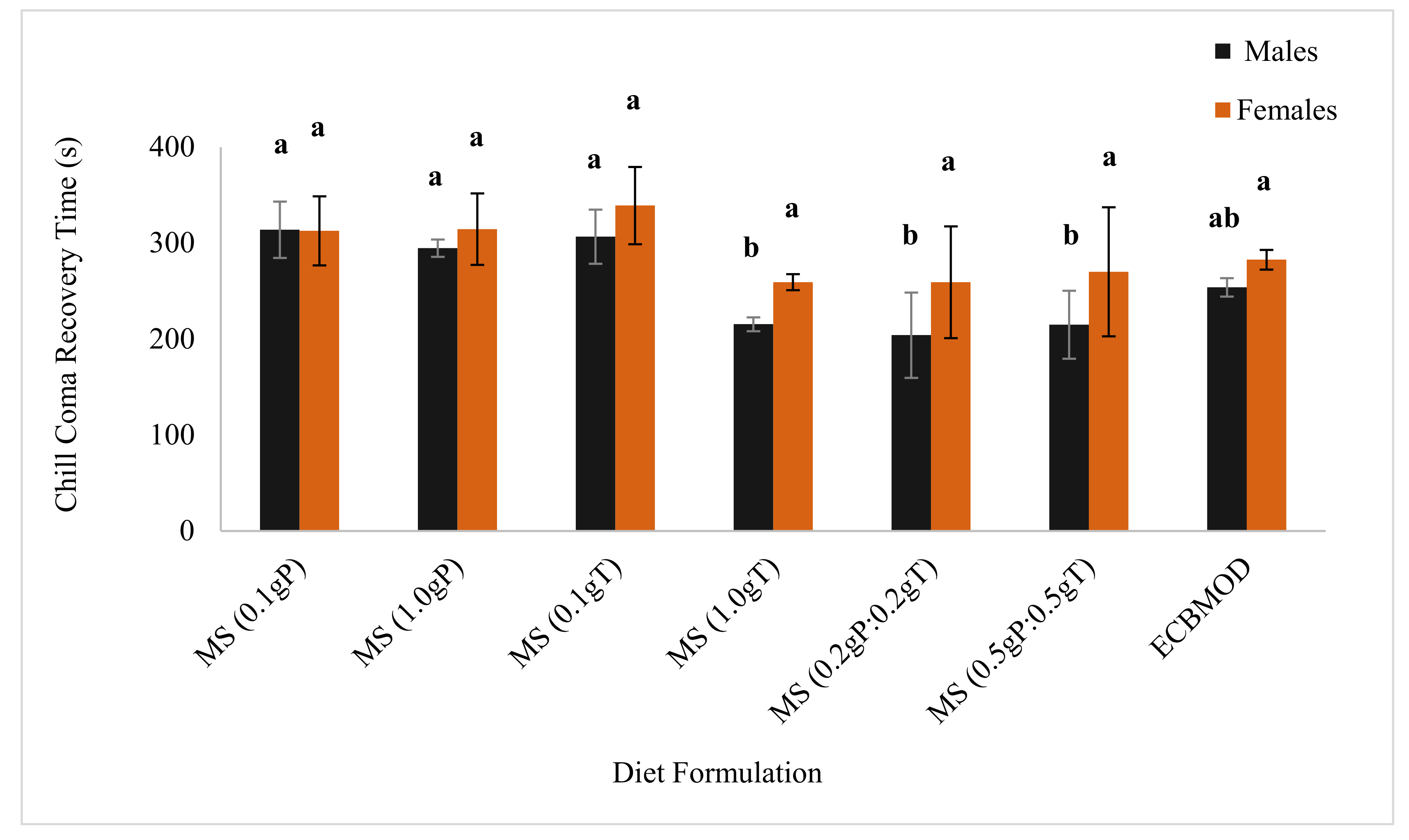
3.5. Chill Coma Recovery Time
Diet Incorporation of Cryoprotectants
3.6. Male and Female Mating Frequency
Diet Incorporation of Cryoprotectants
3.7. Female Fecundity and Fertility
3.7.1. Diet Incorporation of Sterols
3.7.2. Diet Incorporation of Cryoprotectants
3.7.3. Evaluation of Eldana saccharina Performance between Diets
4. Discussion
4.1. Improved Eldana saccharina Growth, Development and Reproduction through the Incorporation of Sterols into the CMT Derived MS Diet Formulation
4.1.1. Insect Development
4.1.2. Pupal Weight
4.2. Enhanced Cold Hardiness of Eldana Saccharina Moths Due to the Incorporation of Cryoprotectants into the CMT Derived MS Diet Formulation
4.2.1. Chill Coma Recovery Time
4.2.2. Fecundity and Fertility
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conlong, D.E. Eldana borer research: The role of the biocontrol insect unit. Proc. S. Afr. Sugar Technol. Assoc. 1992, 64, 106–108. [Google Scholar]
- Babu, G.C.; Sharma, H.C.; Madhumati, T.; Raghavaiah, G.; Murthy, K.V.M.K. Effect of cholesterol, stigmasterol and sitosterol in artificial diet on survival and development of Helicoverpa armigera. IJRSAS 2018, 4, 34. [Google Scholar]
- Behmer, S.T.; Nes, W.D. Insect sterol nutrition and physiology: A global overview. Adv. Insect. Physiol. 2003, 31, 1–72. [Google Scholar]
- Nagata, S.; Omori, Y.; Nagasawa, H. Dietary Sterol Preference in the Silkworm, Bombyx mori. Biosci. Biotechnol. Biochem. 2006, 70, 3094–3098. [Google Scholar] [CrossRef]
- Behmer, S.T.; Grebenok, R.J.; Douglas, A.E. Plant sterols and host plant suitability for a phloem-feeding insect. Funct. Ecol. 2011, 25, 484–491. [Google Scholar] [CrossRef]
- Janson, E.M.; Grebenok, R.J.; Behmer, S.T.; Abbot, P. Same host-plant, different sterols: Variation in sterol metabolism in an insect herbivore community. J. Chem. Ecol. 2009, 35, 1309–1319. [Google Scholar] [CrossRef]
- Bouvaine, S.; Faure, M.L.; Grebenok, R.J.; Behmer, S.T.; Douglas, A.E. A Dietary Test of Putative Deleterious Sterols for the Aphid Myzus persicae. PLoS ONE 2014, 9, e86256. [Google Scholar] [CrossRef] [Green Version]
- Jing, X.; Grebenok, R.J.; Behmer, S.T. Sterol/steroid metabolism and absorption in a generalist and specialist caterpillar: Effects of dietary sterol/steroid structure, mixture and ratio. Insect. Biochem. Mol. Biol. 2013, 43, 580–587. [Google Scholar] [CrossRef]
- Al-Izzi, M.A.J.; Hopkins, T.L. Effects of dietary sterols on development and reproduction of South Western corn borer Diatraea grandiosella Dyar. J. Insect. Physiol. 1982, 28, 189–192. [Google Scholar] [CrossRef]
- Terblanche, J.S. Physiological performance of field-released insects. Curr. Opin. Insect. Sci. 2014, 4, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Chidawanyika, F.; Terblanche, J.S. Rapid thermal responses and thermal tolerance in adult codling moth Cydia pomonella (Lepidoptera: Tortricidae). J. Insect. Physiol. 2011, 57, 108–117. [Google Scholar] [CrossRef]
- Sorensen, J.G.; Addison, M.F.; Terblanche, J.S. Mass-rearing of insects for pest management: Challenges, synergies and advances from evolutionary physiology. J. Crop. Prot. 2012, 38, 87–94. [Google Scholar] [CrossRef]
- Terblanche, J.S.; Mitchell, K.A.; Uys, W.; Short, C.; Boardman, L. Thermal limits to survival and activity in two life stages of false codling moth Thaumatotibia leucotreta (Lepidoptera, Tortricidae). Physiol. Entomol. 2017, 42, 379–388. [Google Scholar] [CrossRef]
- Kleynhans, E.; Mitchell, K.A.; Conlong, D.E.; Terblanche, J.S. Evolved variation in cold tolerance among populations of Eldana saccharina (Lepidoptera: Pyralidae) in South Africa. J. Evol. Biol. 2014, 27, 1149–1159. [Google Scholar] [CrossRef]
- Lee, R.E., Jr.; Denlinger, D.L. (Eds.) Insects at Low Temperature; Chapman & Hall: London, UK, 1991; 513p, ISBN-13: 978-1475701920; ISBN-10: 1475701926. [Google Scholar]
- Kostal, V.; Zahradnickova, H.; Simek, P. Hyperprolinemic larvae of the drosophilid fly, Chymomyza costata, survive cryopreservation in liquid nitrogen. Proc. Natl. Acad. Sci. USA 2011, 108, 13041–13046. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, P.R. Mass rearing and artificial infestation methods for Eldana saccharina Walker. Proc. S. Afr. Sugar Technol. Assoc. 1978, 52, 143–145. [Google Scholar]
- Graham, D.Y.; Conlong, D.E. Improved laboratory rearing of Eldana saccharina (Lepidoptera: Pyralidae) and its indigenous parasitoid Goniozus natalensis (Hymenoptera: Bethylidae). Proc. S. Afr. Sugar Technol. Assoc. 1988, 62, 116–119. [Google Scholar]
- Rutherford, R.S.; Van Staden, J. Development of defined synthetic diets for the culturing of Eldana saccharina Walker (Lepidoptera: Pyralidae). Proc. S. Afr. Sugar Technol. Assoc. 1991, 65, 80–86. [Google Scholar]
- Gillespie, D.Y. Development of mass-rearing methods for the sugarcane borer Eldana saccharina Walker (Lepidoptera: Pyralidae) II: Diet gelling agents. Proc. S. Afr. Sugar Technol. Assoc. 1993, 67, 127–130. [Google Scholar]
- Walton, A.J.; Conlong, D.E. General biology of Eldana saccharina Walker (Lepidoptera: Pyralidae): A target for the sterile insect technique. Fla. Entomol. 2016, 99, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Ngomane, N.C.; Gillespie, D.Y.; Conlong, D.E. The effect of an improved artificial diet formulation on Eldana saccharina Walker rearing, growth and development. Proc. S. Afr. Sugar Technol. Assoc. 2017, 90, 66–77. [Google Scholar]
- Woods, M.J.; Conlong, D.E.; Ngomane, N.; Gillespie, D.; Hoffman, L.C.; Pieterse, E. The development of an improved artificial diet for the mass-rearing of Eldana saccharina Walker (Lepidoptera: Pyralidae). J. Sci. Food. Agric. 2019, 100, 4678–4687. [Google Scholar] [CrossRef]
- Ngomane, N.C.; Pieterse, E.; Woods, M.J.; Conlong, D.E. Formulation of artificial diets for rearing Eldana saccharina Walker (Lepidoptera: Pyralidae) using the Carcass Milling Technique. Insects 2022, 13. Accepted for publication 11 March 2022. [Google Scholar]
- Atkinson, P.R. On the biology, distribution and natural host-plants of Eldana saccharina Walker (Lepidoptera: Pyralidae). Proc. S. Afr. Sugar Technol. Assoc. 1980, 43, 171–194. [Google Scholar]
- MacMillan, H.A.; Williams, C.M.; Staples, J.F.; Sinclair, B.J. Re-establishment of ion homeostasis during chill-coma recovery in the cricket Gryllus pennsylvanicus. Proc. Natl. Acad. Sci. USA 2012, 109, 20750–20755. [Google Scholar] [CrossRef] [Green Version]
- Goebel, R. The effect of temperature on development and reproduction of the sugarcane stalk borer, Chilo sacchariphagus (Bojer 1856) (Lepidoptera: Crambidae). Afr. Entomol. 2006, 14, 103–111. [Google Scholar]
- Sinclair, B.J.; Vernon, P.; Klok, C.J.; Chown, S.L. Insects at low temperatures: An ecological perspective. Trends. Ecol. Evol. 2003, 18, 257–262. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Ross, P.A. Rates and patterns of laboratory adaptation in (mostly) insects. J. Econ. Entomol. 2018, 20, 1–9. [Google Scholar] [CrossRef]
- Daniel, C.A. Improving the Cold Tolerance of False Codling Moth, Thaumatotibia leucotreta, for Better Performance in a Sterile Insect Release Programme. Master of Science Thesis, Rhodes University, Grahamstown, South Africa, 2016; pp. 13–19. [Google Scholar]
- Boersma, N. Impacts of Temperature Variation on Performance, Life History and Flight Ability of the False Codling Moth, Thaumatotibia leucotreta (Lepidoptera: Tortricidae). Master of Science Thesis, Stellenbosch University, Stellenbosch, South Africa, 2018; 89p. [Google Scholar]
- Assefa, Y.; Conlong, D.E.; Mitchell, A. Status of Eldana saccharina (Lepidoptera: Pyralidae), its host plants and natural enemies in Ethiopia. Bull. Entomol. Res. 2006, 96, 497–504. [Google Scholar]
- Colinet, H.; Rinehart, J.P.; Yocum, G.D.; Greenlee, K.J. Mechanisms underpinning the beneficial effects of fluctuating thermal regimes in insect cold tolerance. J. Exp. Biol. 2018, 221, jeb164806. [Google Scholar] [CrossRef] [Green Version]
- Gibert, P.; Moreteau, B.; Petavy, G.; Karan, D.; David, J.R. Chill-coma tolerance, a major climatic adaptation among Drosophila species. Evol. 2001, 55, 1063–1068. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Alvarado, L.E.C.; Ferguson, L.V. An invitation to measure insect cold tolerance: Methods, approaches, and workflow. Biol. Public. 2015, 68, 1–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Findsen, A.; Pedersen, T.H.; Petersen, A.G.; Nielsen, O.B.; Overgaard, J. Why do insects enter and recover from chill coma? Low temperature and high extracellular potassium compromise muscle function in Locusta migratoria. J. Exp. Biol. 2014, 217, 1297–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, L.H.; Kristensen, T.; Loeschcke, V.; Toft, S.; Mayntz, D. Protein and carbohydrate composition of larval food affects tolerance to thermal stress and desiccation in adult Drosophila melanogaster. J. Insect. Physiol. 2009, 56, 336–340. [Google Scholar] [CrossRef]
- Nikolouli, K.; Colinet, H.; Renault, D.; Enriquez, T.; Mouton, L.; Gibert, P.; Sassu, F.; Cáceres, C.; Stauffer, C.; Pereira, R.; et al. Sterile insect technique and Wolbachia symbiosis as potential tools for the control of the invasive species Drosophila suzukii. J. Pest. Sci. 2018, 91, 489–503. [Google Scholar] [CrossRef] [Green Version]
- Mensch, J.; Hurtado, J.; Zermoglio, P.F.; de la Vega, J.; Rolandi, C.; Schilman, P.E.; Markow, T.A.; Hasson, E. Enhanced fertility and chill tolerance after cold-induced reproductive arrest in females of temperate species of the Drosophila buzzatii complex. J. Exp. Biol. 2017, 220, 713–721. [Google Scholar] [CrossRef] [Green Version]
- Mudavanhu, P.; Addison, P.; Conlong, D.E. Effect of mass rearing and gamma irradiation on the mating behaviour of Eldana saccharina. Entomol. Exp. Appl. 2016, 162, 159–167. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| MS Diets Containing Sterols vs. Control Diet (ECBMOD) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Diet Name | MS (0.1 gC) | MS (1.0 gC) | MS (0.1 gS) | MS (1.0 gS) | MS (0.2 gC:0.2 gS) | MS (0.5 gC:0.5 gS) | ECBMOD (Control) | |
| Ingredients | Unit | Wt. | Wt. | Wt. | Wt. | Wt. | Wt. | Wt. |
| Carrageenan gel | g | 15.00 | 15.00 | 15.00 | 15.00 | 15.00 | 15.00 | |
| Agar powder | g | 4.60 | ||||||
| Lucerne meal | g | 250.00 | 250.00 | 250.00 | 250.00 | 250.00 | 250.00 | |
| Rabbit meal | g | 226.40 | ||||||
| Wheat bran | g | 55.80 | 55.80 | 55.80 | 55.80 | 55.80 | 55.80 | 56.60 |
| Yeast extract | g | 3.40 | ||||||
| Ground chickpea | g | 53.00 | 53.00 | 53.00 | 53.00 | 53.00 | 53.00 | 56.60 |
| Full cream milk powder | g | 7.60 | 7.60 | 7.60 | 7.60 | 7.60 | 7.60 | 22.60 |
| Whole egg powder | g | 28.40 | 28.40 | 28.40 | 28.40 | 28.40 | 28.40 | 28.20 |
| Sucrose | g | 66.20 | 66.20 | 66.20 | 66.20 | 66.20 | 66.20 | 64.60 |
| Sodium chloride | g | 0.60 | ||||||
| Nipagin | g | 6.40 | 6.40 | 6.40 | 6.40 | 6.40 | 6.40 | 6.40 |
| Sodium propionate | g | 10.40 | 10.40 | 10.40 | 10.40 | 10.40 | 10.40 | 10.40 |
| Oxytetracycline | g | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| Ascorbic acid | g | 6.40 | 6.40 | 6.40 | 6.40 | 6.40 | 6.40 | 6.40 |
| Acetic acid | mL | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 |
| Citric acid | g | 2.60 | 2.60 | 2.60 | 2.60 | 2.60 | 2.60 | 2.60 |
| Tri-sodium citrate | g | 2.60 | 2.60 | 2.60 | 2.60 | 2.60 | 2.60 | 2.60 |
| Vit + min premix | g | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 | |
| Cholesterol | g | 0.10 | 1.00 | 0.20 | 0.50 | |||
| Stigmasterol | g | 0.10 | 1.00 | 0.20 | 0.50 | |||
| Total | 515.30 | 516.20 | 515.30 | 515.60 | 515.60 | 516.20 | 502.00 | |
| Water for agar | mL | 500.00 | ||||||
| Water balance | mL | 1500.00 | 1500.00 | 1500.00 | 1500.00 | 1500.00 | 1500.00 | 1000.00 |
| Total diet volume | mL | 2015.30 | 2016.20 | 2015.30 | 2016.20 | 2015.60 | 2016.20 | 2002.00 |
| MS Diets Containing Cryoprotectants vs. Control Diet (ECBMOD). | ||||||||
|---|---|---|---|---|---|---|---|---|
| Diet Name | MS (0.1 gP) | MS (1.0 gP) | MS (0.1 gT) | MS (1.0 gT) | MS (0.2 gP:0.2 gT) | MS (0.5 gP:0.5 gT) | ECBMOD (Control) | |
| Ingredients | Unit | Wt. | Wt. | Wt. | Wt. | Wt. | Wt. | Wt. |
| Carrageenan gel | g | 15.00 | 15.00 | 15.00 | 15.00 | 15.00 | 15.00 | |
| Agar powder | g | 4.60 | ||||||
| Lucerne meal | g | 250.00 | 250.00 | 250.00 | 250.00 | 250.00 | 250.00 | |
| Rabbit meal | g | 226.40 | ||||||
| Wheat bran | g | 55.80 | 55.80 | 55.80 | 55.80 | 55.80 | 55.80 | 56.60 |
| Yeast extract | g | 3.40 | ||||||
| Ground chickpea | g | 53.00 | 53.00 | 53.00 | 53.00 | 53.00 | 53.00 | 56.60 |
| Full cream milk powder | g | 7.60 | 7.60 | 7.60 | 7.60 | 7.60 | 7.60 | 22.60 |
| Whole egg powder | g | 28.40 | 28.40 | 28.40 | 28.40 | 28.40 | 28.40 | 28.20 |
| Sucrose | g | 66.20 | 66.20 | 66.20 | 66.20 | 66.20 | 66.20 | 64.60 |
| Sodium chloride | g | 0.60 | ||||||
| Nipagin | g | 6.40 | 6.40 | 6.40 | 6.40 | 6.40 | 6.40 | 6.40 |
| Sodium propionate | g | 10.40 | 10.40 | 10.40 | 10.40 | 10.40 | 10.40 | 10.40 |
| Oxytetracycline | g | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| Ascorbic acid | g | 6.40 | 6.40 | 6.40 | 6.40 | 6.40 | 6.40 | 6.40 |
| Acetic acid | mL | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 |
| Citric acid | g | 2.60 | 2.60 | 2.60 | 2.60 | 2.60 | 2.60 | 2.60 |
| Tri-sodium citrate | g | 2.60 | 2.60 | 2.60 | 2.60 | 2.60 | 2.60 | 2.60 |
| Vit + min premix | g | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 | |
| L-Proline | g | 0.10 | 1.00 | 0.20 | 0.50 | |||
| Trehalose | g | 0.10 | 1.00 | 0.20 | 0.50 | |||
| Total | 515.30 | 516.20 | 515.30 | 515.60 | 515.60 | 516.20 | 502.00 | |
| Water for agar | mL | 500.00 | ||||||
| Water balance | mL | 1500.00 | 1500.00 | 1500.00 | 1500.00 | 1500.00 | 1500.00 | 1000.00 |
| Total diet volume | mL | 2015.30 | 2016.20 | 2015.30 | 2016.20 | 2015.60 | 2016.20 | 2002.00 |
| Diet Formulation | % Survival at First Pupation (Day 20) | % Survival at Full Pupal Development (Day 27) |
|---|---|---|
| MS (0.1 gC) | 100.00 ± 0 | 98.00 ± 1.01 |
| MS (1.0 gC) | 100.00 ± 0 | 100.00 ± 0 |
| MS (0.1 gS) | 93.00 ± 4.54 | 97.00 ± 1.28 |
| MS (1.0 gS) | 100.00 ± 0 | 96.00 ± 1.40 |
| MS (0.2 gC:0.2 gS) | 100.00 ± 0 | 99.00 ± 0.59 |
| MS (0.5 gC:0.5 gS) | 93.00 ± 4.54 | 99.00 ± 0.59 |
| ECBMOD (Control) | 100.00 ± 0 | 100.00 ± 0 |
| Life Stage Distribution (%) | ||||||
|---|---|---|---|---|---|---|
| Diet Formulation | 1st/2nd Instar * | 3rd/4th Instar | 5th/6th Instar | Pre-Pupae | Pupae | Mortality |
| MS (0.1 gC) | 0 ± 0 | 0 ± 0 b | 27.0 ± 0.6 d | 9.0 ± 0.6 b | 64.0 ± 0.6 b | 0 ± 0 c |
| MS (1.0 gC) | 0 ± 0 | 0 ± 0 b | 30.0 ± 0.6 c | 7.0 ± 0.6 bc | 63.0 ± 0.6 b | 0 ± 0 c |
| MS (0.1 gS) | 0 ± 0 | 0 ± 0 b | 29.0 ± 0.6 cd | 2.0 ± 0.6 d | 62.0 ± 0.6 b | 7.0 ± 0.6 a |
| MS (1.0 gS) | 0 ± 0 | 0 ± 0 b | 16.0 ± 0.6 f | 12.0 ± 0.6 a | 72.0 ± 0.6 a | 0 ± 0 c |
| MS (0.2 gC:0.2 gS) | 0 ± 0 | 0 ± 0 b | 24.0 ± 0.6 e | 6.0 ± 0.6 c | 70.0 ± 0.6 a | 0 ± 0 c |
| MS (0.5 gC:0.5 gS) | 0 ± 0 | 0 ± 0 b | 33.0 ± 0.6 b | 7.0 ± 0.6 bc | 55.0 ± 0.6 c | 5.0 ± 0.6 b |
| ECBMOD (Control) | 0 ± 0 | 35.0 ± 0.6 a | 50.0 ± 0.6 a | 0 ± 0.0 d | 15.0 ± 0.6 d | 0 ± 0 c |
| Life Stage Distribution (%) | |||||
|---|---|---|---|---|---|
| Diet Formulation | Larvae | Pre-Pupae | Pupae | Moths | Mortality |
| MS (0.1 gC) | 2.0 ± 0.6 c | 12.0 ± 0.6 b | 82.0 ± 0.6 c | 4.0 ± 0.6 a | 0 ± 0 a |
| MS (1.0 gC) | 0 ± 0 c | 0 ± 0 d | 96.0 ± 0.6 a | 4.0 ± 0.6 a | 0 ± 0 a |
| MS (0.1 gS) | 2.0 ± 0.6 c | 11.0 ± 0.6 bc | 86.0 ± 0.6 b | 1.0 ± 0.6 b | 0 ± 0 a |
| MS (1.0 gS) | 6.0 ± 0.6 b | 9.0 ± 0.6 c | 82.0 ± 0.6 c | 2.0 ± 0.6 ab | 1.0 ± 0.6 a |
| MS (0.2 gC:0.2 gS) | 6.0 ± 0.6 b | 11.0 ± 0.6 bc | 82.0 ± 0.6 c | 0 ± 0 b | 1.0 ± 0.6 a |
| MS (0.5 gC:0.5 gS) | 7.0 ± 0.6 b | 19.0 ± 0.6 a | 73.0 ± 0.6 d | 0 ± 0 b | 1.0 ± 0.6 a |
| ECBMOD (Control) | 19.0 ± 0.6 a | 9.0 ± 0.6 c | 70.0 ± 0.6 e | 0 ± 0 b | 2.0 ± 0.6 a |
| Diet Formulation | Moth Emergence (%) | Sex Ratio (M:F) |
|---|---|---|
| MS (0.1 gC) | 96.0 ± 0.6 b | 1.3 ± 0.1 a |
| MS (1.0 gC) | 96.0 ± 0.6 b | 1.1 ± 0.1 a |
| MS (0.1 gS) | 100.0 ± 0.3 a | 0.8 ± 0.1 b |
| MS (1.0 gS) | 94.0 ± 0.6 c | 0.9 ± 0.1 b |
| MS (0.2 gC:0.2 gS) | 100.0 ± 0.3 a | 1.2 ± 0.1 a |
| MS (0.5 gC:0.5 gS) | 92.0 ± 0.6 c | 1.1 ± 0.1 a |
| ECBMOD (Control) | 97.0 ± 0.6 b | 0.8 ± 0.1 b |
| Diet Formulation | Moth Emergence (%) | Sex Ratio (M:F) |
|---|---|---|
| MS (0.1 gP) | 95.0 ± 0.6 b | 0.9 ± 0.1 c |
| MS (1.0 gP) | 96.0 ± 0.6 b | 1.3 ± 0.1 b |
| MS (0.1 gT) | 89.0 ± 0.6 c | 1.3 ± 0.1 b |
| MS (1.0 gT) | 100.0 ± 0.3 a | 1.2 ± 0.1 b |
| MS (0.2 gP:0.2 gT) | 91.0 ± 0.6 c | 1.0 ± 0.1 bc |
| MS (0.5 gP:0.5 gT) | 96.0 ± 0.6 b | 1.0 ± 0.1 bc |
| ECBMOD (Control) | 89.0 ± 0.6 c | 1.7 ± 0.1 a |
| Male Mating Frequency | Female Mating Frequency | |||
|---|---|---|---|---|
| Diet Formulation | Chill Coma Exposed | Non-Chill Exposed | Chill Coma Exposed | Non-Chill Exposed |
| MS (0.1 gP) | 3.00 ± 1.00 | 3.00 ± 0.67 | 3.00 ± 0.58 | 2.00 ± 0.33 |
| MS (1.0 gP) | 3.00 ± 0.33 | 3.00 ± 0.33 | 3.00 ± 0.33 | 3.00 ± 0.33 |
| MS (0.1 gT) | 3.00 ± 0.67 | 4.00 ± 0.33 | 3.00 ± 0.33 | 2.00 ± 0.33 |
| MS (1.0 gT) | 4.00 ± 0.33 | 4.00 ± 0.58 | 2.00 ± 0.33 | 2.00 ± 0.33 |
| MS (0.2 gP:0.2 gT) | 3.00 ± 0.67 | 4.00 ± 0.33 | 2.00 ± 0.68 | 2.00 ± 0.33 |
| MS (0.5 gP:0.5 gT) | 3.00 ± 0.58 | 3.00 ± 0.33 | 2.00 ± 0.6 | 2.00 ± 0.33 |
| ECBMOD (Control) | 3.00 ± 0.33 | 4.00 ± 0.33 | 2.00 ± 0.33 | 2.00 ± 0.33 |
| Diet Formulation | Fecundity (n) | Fertility (%) |
|---|---|---|
| MS (0.1 gC) | 879.00 ± 40.32 | 94.00 ± 5.65 |
| MS (1.0 gC) | 955.00 ± 28.81 | 96.00 ± 1.57 |
| MS (0.1 gS) | 827.00 ± 90.30 | 98.00 ± 2.03 |
| MS (1.0 gS) | 922.00 ± 105.06 | 98.00 ± 1.34 |
| MS (0.2 gC:0.2 gS) | 891.00 ± 148.86 | 91.00 ± 7.59 |
| MS (0.5 gC:0.5 gS) | 906.00 ± 20.25 | 100.00 ± 0.15 |
| ECBMOD (Control) | 789.00 ± 48.66 | 94.00 ± 1.77 |
| Female Fecundity (n) | Female Fertility (%) | |||
|---|---|---|---|---|
| Diet Formulation | Chill Coma Exposed | Non-Chill Exposed | Chill Coma Exposed | Non-Chill Exposed |
| MS (0.1 gP) | 710.00 ± 7.22 | 936.00 ± 39.50 | 10.00 ± 9.68 b | 90.00 ± 7.49 a |
| MS (1.0 gP) | 776.00 ± 240.01 | 733.00 ± 82.50 | 30.00 ± 2.00 a | 95.00 ± 4.85 a |
| MS (0.1 gT) | 765.00 ± 54.58 | 800.00 ± 46.00 | 42.00 ± 6.28 a | 78.00 ± 21.9 a |
| MS (1.0 gT) | 783.00 ± 47.61 | 786.00 ± 49.19 | 41.00 ± 5.99 a | 80.00 ± 7.73 a |
| MS (0.2 gP:0.2 gT) | 813.00 ± 150.82 | 853.00 ± 85.50 | 34.00 ± 5.82 a | 95.00 ± 1.31 a |
| MS (0.5 gP:0.5 gT) | 723.00 ± 30.84 | 732.00 ± 60.50 | 14.00 ± 7.59 b | 64.00 ± 13.19 a |
| ECBMOD (Control) | 815.00 ± 25.42 | 841.00 ± 41.15 | 31.00 ± 3.72 a | 88.00 ± 6.45 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngomane, N.C.; Terblanche, J.S.; Conlong, D.E. The Addition of Sterols and Cryoprotectants to Optimize a Diet Developed for Eldana saccharina Walker (Lepidoptera: Pyralidae) Using the Carcass Milling Technique. Insects 2022, 13, 314. https://doi.org/10.3390/insects13040314
Ngomane NC, Terblanche JS, Conlong DE. The Addition of Sterols and Cryoprotectants to Optimize a Diet Developed for Eldana saccharina Walker (Lepidoptera: Pyralidae) Using the Carcass Milling Technique. Insects. 2022; 13(4):314. https://doi.org/10.3390/insects13040314
Chicago/Turabian StyleNgomane, Nomalizo C., John S. Terblanche, and Des E. Conlong. 2022. "The Addition of Sterols and Cryoprotectants to Optimize a Diet Developed for Eldana saccharina Walker (Lepidoptera: Pyralidae) Using the Carcass Milling Technique" Insects 13, no. 4: 314. https://doi.org/10.3390/insects13040314