
Plant-Based Bioinsecticides for Mosquito Control: Impact on Insecticide Resistance and Disease Transmission


Abstract
:Simple Summary
Abstract
1. Introduction
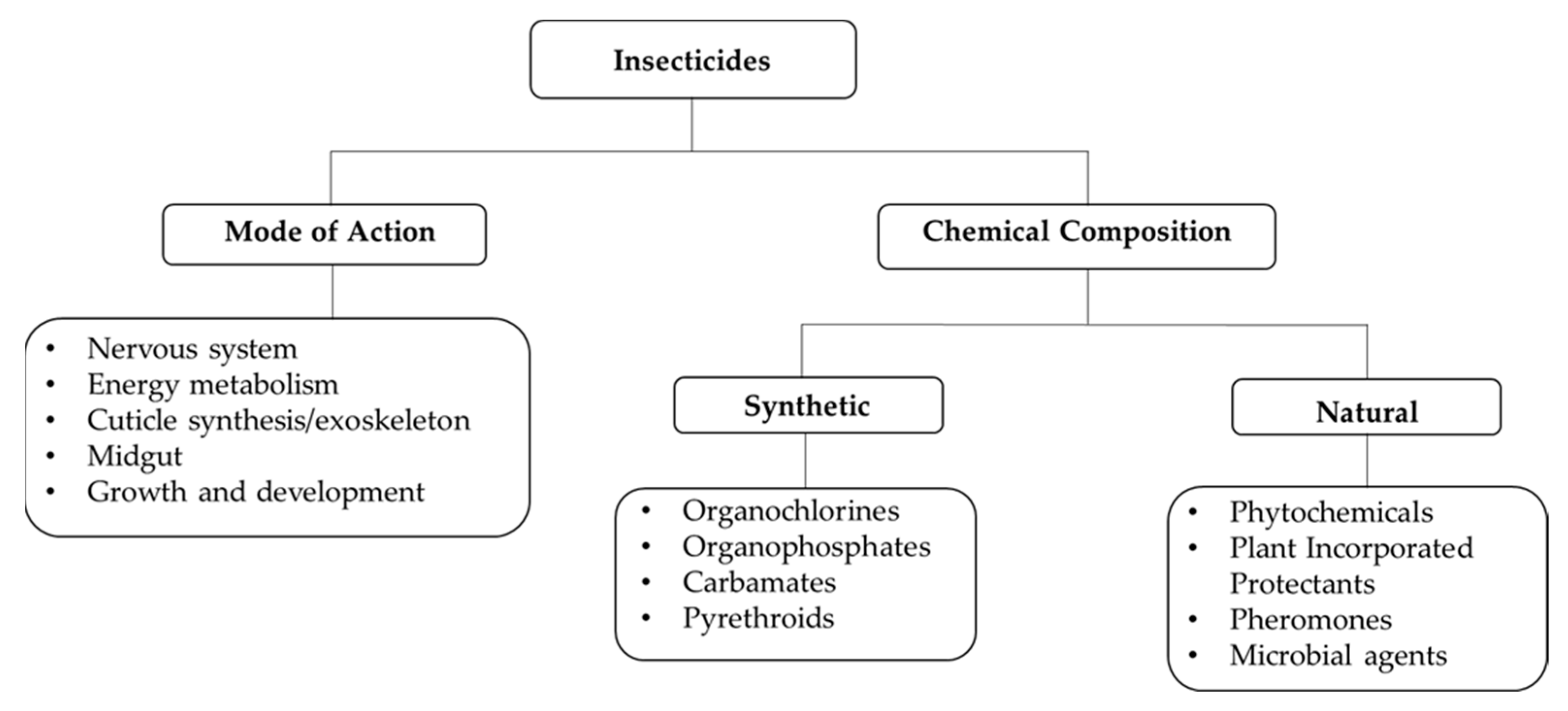
2. Insecticide-Based Mosquito Control Strategies
3. Insecticide Resistance in Mosquitoes
4. Plant-Based Bioinsecticides
5. Plant-Based Compounds and Mosquito Control
5.1. Essential Oils

{kind=link}
| Type of Botanical Product | Plant Family | Activity | Mechanism of Action | Mosquito Species | References |
|---|---|---|---|---|---|
| Essential Oils Monoterpenes: linalool, cuminaldehyde, 1,8-cineole, limonene, fenchone, eugenol, γ-terpineol, cinnamic alcohol, geraniol, β-citronellol, P-menthane-3,8 diol, α-pinene, β-pinene, p-cymene, thymol, terpinolene, camphor, citronellal, sabinene, carvacrol Sesquiterpenes: guaiol, α-bisabolol, α-cadinol, germacrene D, β-caryophyllene, nootkatone Diterpenoids: diterpene alcohol, phytol Aromatic phenol Coumarin | Anacardiaceae Annonaceae Apiaceae Asteraceae Geraniaceae Lamiaceae Lauraceae Poaceae Rutaceae Myrtaceae Verbenaceae | larvicidal, pupaecidal, ovicidal, adulticidal, repellent, antifeedant, growth and reproduction inhibitors | Inhibition of AChE Blockage of GABA-gated chloride channels Agonist of octopamine receptors | Cx. pipiens pallens Cx. quinquefasciatus Cx. pipiens biotype molestus Ae. aegypti Ae. albopictus An. gambiae An. stephensi | [120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146] |
| Neem oil azadirachtin, meliantriol, salannin, desacetyl salannin, nimbin, desacetyl nimbin, nimbidin, nimbolide, deacetylgedunin, gedunin, 17-hydroxyazadiradione, deacetylnimbin | Meliaceae | repellent, ovicidal, larvicidal, feeding deterrence, fecundity suppression, toxicity, growth regulation, oviposition deterrence | growth inhibitors, hormonal disruption (ecdysone blocker), molting aberrations, interference with phagostimulants | An. gambiae Ae. aegypti Ae. albopictus An. stephensi Cx. quinquefasciatus | [147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165] |
| Pyrethrum esters of chrysanthemic acid: pyrethrin I, cinerin I, jasmolin I esters of pyrethric acid: pyrethrin II, cinerin II, jasmolin II | Asteraceae | repellent, knock-down effect, blood-feding inhibition | voltage-gated sodium channel modulator | An. gambiae | [166,167,168,169,170] |
| Alkaloids alpha-solanin ricinine pyridine nicotine diterpene nornicotine anabasine | Berberidaceae Fabaceae Solanaceae Ranunculaceae Euphorbiaceae | repellent, larvicidal | interfering with cellular and physiological functions, inhibition of AChE activity, regulation of hormone activity, toxicity, agonist of acetycholine receptor | Ae. aegypti An. arabiensis An. gambiae Ae. albopictus An. stephensi Cx. pipiens | [171,172,173,174,175,176,177,178,179,180,181] |
| Flavonoids | Zingiberaceae | larvicidal | inhibition of AChE, degradation of cell membranes acting as stomach poisons | Ae. aegypti | [182,183,184] |
| Rotenone | Fabaceae | larvicidal | inhibitor of the cellular respiration system | Ae. aegypti | [185] |
5.2. Neem
5.3. Pyrethrum
5.4. Alkaloids
5.5. Other Plant Compounds
6. Assessment of Plant-Based Bioinsecticide Efficacy in Mosquito Control
7. Effective Use of Plant-Based Bioinsecticides in Resistant Mosquito Populations
8. How to Improve Plant-Based Bioinsecticide Efficacy in Mosquito Control Strategies?
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kilpatrick, A.M. Globalization, land use and the invasion of West Nile virus NIH Public Access. Science 2011, 334, 323–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. World Malaria Report 2020: 20 Years of Global Progress and Challenges; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Du, M.; Jing, W.; Liu, M.; Liu, J. The Global Trends and Regional Differences in Incidence of Dengue Infection from 1990 to 2019: An Analysis from the Global Burden of Disease Study 2019. Infect. Dis. Ther. 2021, 10, 1625–1643. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Stanaway, J.D.; Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Coffeng, L.E.; Brady, O.J.; Hay, S.I.; Bedi, N.; Bensenor, I.M.; Castañeda-Orjuela, C.A.; et al. The global burden of dengue: An analysis from the Global Burden of Disease Study 2013. Lancet Infect. Dis. 2016, 16, 712–723. [Google Scholar] [CrossRef] [Green Version]
- Paixão, E.S.; Teixeira, M.G.; Rodrigues, L.C. Zika, chikungunya and dengue: The causes and threats of new and re-emerging arboviral diseases. BMJ Glob. Health 2018, 3, e000530. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, M.U.G.; Reiner, R.C.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef]
- Altizer, S.; Ostfeld, R.S.; Johnson, P.T.J.; Kutz, S.; Harvell, C.D. Climate change and infectious diseases: From evidence to a predictive framework. Science 2013, 341, 514–519. [Google Scholar] [CrossRef] [Green Version]
- Mordecai, E.A.; Ryan, S.J.; Caldwell, J.M.; Shah, M.M.; LaBeaud, A.D. Climate change could shift disease burden from malaria to arboviruses in Africa. Lancet Planet Health 2020, 4, e416–e423. [Google Scholar] [CrossRef]
- WHO. Global Report on Insecticide Resistance in Malaria Vectors: 2010–2016; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Richards, S.L.; Byrd, B.D.; Reiskind, M.H.; White, A.V. Assessing Insecticide Resistance in Adult Mosquitoes: Perspectives on Current Methods. Environ. Health Insights 2020, 14, 1–7. [Google Scholar] [CrossRef]
- Lines, J.D.; Myamba, J.; Curtis, C.F. Experimental hut trials of permethrin-impregnated mosquito nets and eave curtains against malaria vectors in Tanzania. Med. Vet. Entomol. 1987, 1, 37–51. [Google Scholar] [CrossRef]
- Lindsay, S.W.; Adiamah, J.H.; Miller, J.E.; Armstrong, J.R.M. Pyrethroid-treated bednet effects on mosquitoes of the Anopheles gambiae complex in The Gambia. Med. Vet. Entomol. 1991, 5, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.E.; Lindsay, S.W.; Armstrong, J.R. Experimental hut trials of bednets impregnated with synthetic pyrethroid or organophosphate insecticide for mosquito control in The Gambia. Med. Vet. Entomol. 1991, 5, 465–476. [Google Scholar] [CrossRef]
- Gimnig, J.E.; Vulule, J.M.; Lo, T.Q.; Kamau, L.; Kolczak, M.S.; Phillips-Howard, P.A.; Mathenge, E.M.; Ter Kuile, F.O.; Nahlen, B.L.; Hightower, A.W. Impact of permethrin-treated bed nets on the entomologic indices in an area of intense year-round malaria transmission. Am. J. Trop. Med. Hyg. 2003, 68, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Lengeler, C. Insecticide-treated bed nets and curtains for preventing malaria. Cochrane Database Syst. Rev. 2004, 2, CD000363. [Google Scholar] [CrossRef]
- Killeen, G.F. Characterizing, controlling and eliminating residual malaria transmission. Malar. J. 2014, 13, 330. [Google Scholar] [CrossRef] [Green Version]
- Killeen, G.F.; Smith, T.A.; Ferguson, H.M.; Mshinda, H.; Abdulla, S.; Lengeler, C.; Kachur, S.P. Preventing Childhood Malaria in Africa by Protecting Adults from Mosquitoes with Insecticide-Treated Nets. PLoS Med. 2007, 4, e229. [Google Scholar] [CrossRef] [Green Version]
- Pluess, B.; Tanser, F.C.; Lengeler, C.; Sharp, B.L. Indoor residual spraying for preventing malaria. Cochrane Database Syst. Rev. 2010, 4, CD006657. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cibulskis, R.E.; Alonso, P.; Aponte, J.; Aregawi, M.; Barrette, A.; Bergeron, L.; Fergus, C.A.; Knox, T.; Lynch, M.; Patouillard, E.; et al. Malaria: Global progress 2000–2015 and future challenges. Infect. Dis. Poverty 2016, 5, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soderlund, D.M.; Bloomquist, J.R. Neurotoxic actions of pyrethroid insecticides. Annu. Rev. Entomol. 1989, 34, 77–96. [Google Scholar] [CrossRef]
- Lund, A.E.; Narahashi, T. Kinetics of sodium channel modification as the basis for the variation in the nerve membrane effects of pyrethroids and DDT analogs. Pest Biochem. Physiol. 1983, 20, 203–216. [Google Scholar] [CrossRef]
- Soderlund, D.M. Mode of action of pyrethrins and pyrethroids. In Pyrethrum Flowers: Production, Chemistry, Toxicology, and Uses; Casida, J.E., Quistad, G.B., Eds.; Oxford University Press: New York, NY, USA, 1995; pp. 217–233. [Google Scholar]
- Narahashi, T. Neuronal ion channels as the target sites of insecticides. Pharmacol. Toxicol. 1996, 78, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bowman, N.M.; Akialis, K.; Cave, G.; Barrera, R.; Apperson, C.S.; Meshnick, S.R. Pyrethroid insecticides maintain repellent effect on knock-down resistant populations of Aedes aegypti mosquitoes. PLoS ONE 2018, 13, e0196410. [Google Scholar] [CrossRef] [Green Version]
- Ranson, H.; Jensen, B.; Vulule, J.M.; Wang, X.; Hemingway, J.; Collins, F.H. Identification of a point mutation in the voltage-gated sodium channel gene of Kenyan Anopheles gambiae associated with resistance to DDT and pyrethroids. Insect Mol. Biol. 2000, 9, 491–497. [Google Scholar] [CrossRef]
- Etang, J.; Chandre, F.; Guillet, P.; Manga, L. Reduced bio-efficacy of permethrin EC impregnated bednets against an Anopheles gambiae strain with oxidase-based pyrethroid tolerance. Malar. J. 2004, 3, 46. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.G.; Kim, D.; Pham, A.; Paul, C.J. A meta-regression analysis of the effectiveness of mosquito nets for malaria control: The value of long-lasting insecticide nets. Int. J. Environ. Res. Public Health 2018, 15, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eskenazi, B.; Chevrier, J.; Rosas, L.G.; Anderson, H.A.; Bornman, M.S.; Bouwman, H.; Chen, A.; Cohn, B.A.; de Jager, C.; Henshel, D.S.; et al. The Pine River Statement: Human health consequences of DDT use. Environ. Health Perspect. 2009, 117, 1359–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouwman, H.; van den Berg, H.; Kylin, H. DDT and malaria prevention: Addressing the paradox. Environ. Health Perspect. 2011, 119, 744–747. [Google Scholar] [CrossRef] [Green Version]
- Hosie, A.M.; Aronstein, K.; Sattelle, D.B.; ffrench-Constant, R.H. Molecular biology of insect neuronal GABA receptors. Trends Neurosci. 1997, 20, 578–583. [Google Scholar] [CrossRef]
- Lawrence, L.J.; Casida, J.E. Stereospecifc action of pyrethroid insecticides on the gamma-aminobutyric acid receptor-ionophore complex. Science 1983, 221, 1399–1401. [Google Scholar] [CrossRef] [PubMed]
- Bloomquist, J.R. Toxicology, mode of action and target site-mediated resistance to insecticides acting on chloride channels. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1993, 106, 301–314. [Google Scholar] [CrossRef]
- Buckingham, S.D.; Ihara, M.; Sattelle, D.B.; Matsuda, K. Mechanisms of Action, Resistance and Toxicity of Insecticides Targeting GABA Receptors. Curr. Med. Chem. 2017, 24, 2935–2945. [Google Scholar] [CrossRef]
- Mengle, D.C.; Casida, J.E. Inhibition and recovery of brain cholinesterase activity in house flies poisoned with organophosphate and carbamate compounds. J. Econ. Entomol. 1958, 51, 750–757. [Google Scholar] [CrossRef]
- Fukuto, T.R. Mechanism of action of organophosphorus and carbamate insecticides. Environ. Health Perspect. 1990, 87, 245–254. [Google Scholar] [CrossRef]
- Fournier, D.; Mutero, A. Modification of acetylcholinesterase as a mechanism of resistance to insecticides. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1994, 108, 19–31. [Google Scholar] [CrossRef]
- Smith, G. Kern MAD has circumstantial evidence on DDT resistance. Mosquito Buzz 1949, 3, 2. [Google Scholar]
- Gjullin, C.M.; Peters, R.F. Recent studies of mosquito resistance to insecticides in California. Mosq. News 1952, 12, 1–7. [Google Scholar]
- Akogbeto, M.; Yakoubou, S. Resistance of malaria vectors to pyrethroids used for impregnated mosquito nets in Benin, West Africa. Bull. Soc. Path. Exot. 1999, 92, 123–130. [Google Scholar]
- Ranson, H.; N’Guessan, R.; Lines, J.; Moiroux, N.; Nkuni, Z.; Corbel, V. Pyrethroid resistance in African anopheline mosquitoes: What are the implications for malaria control? Trends Parasitol. 2011, 27, 91–98. [Google Scholar] [CrossRef]
- Deletre, E.; Martin, T.; Duménil, C.; Chandre, F. Insecticide resistance modifies mosquito response to DEET and natural repellents. Parasites Vectors 2019, 12, 89. [Google Scholar] [CrossRef] [Green Version]
- WHO. Test Procedures for Insecticide Resistance Monitoring in Malaria Vector Mosquitoes, 2nd ed.; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Rozendaal, J.A. House-spraying with residual insecticides. In Vector Control—Methods for Use by Individuals and Communities; World Health Organization: Geneva, Switzerland, 1997; pp. 357–384. [Google Scholar]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annu. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Plan for Insecticide Resistance Management in Malaria Vectors; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Liu, N. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef]
- Andreazza, F.; Oliveira, E.E.; Martins, G.F. Implications of Sublethal Insecticide Exposure and the Development of Resistance on Mosquito Physiology, Behavior, and Pathogen Transmission. Insects 2021, 12, 917. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Torres, D.; Chandre, F.; Williamson, M.S.; Darriet, F.; Berge, J.B.; Devonshire, A.L.; Guillet, P.; Pasteur, N.; Pauron, D. Molecular characterization of pyrethroid knockdown resistance (kdr) in the major malaria vector Anopheles gambiae s.s. Insect Mol. Biol. 1998, 7, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Dong, K. Insect sodium channels and insecticide resistance. Invert. Neurosci. 2007, 7, 17–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, E.E.; Du, Y.; Nomura, Y.; Dong, K. A residue in the transmembrane segment 6 of domain I in insect and mammalian sodium channels regulate differential sensitivities to pyrethroid insecticides. Neurotoxicology 2013, 38, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Rinkevich, F.D.; Du, Y.; Dong, K. Diversity and convergence of sodium channel mutations involved in resistance to pyrethroids. Pestic. Biochem. Physiol. 2013, 106, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Dong, K.; Du, Y.; Rinkevich, F.; Nomura, Y.; Xu, P.; Wang, L.; Silver, K.; Zhorov, B.S. Molecular biology of insect sodium channels and pyrethroid resistance. Insect Biochem. Mol. Biol. 2014, 50, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Silver, K.S.; Du, Y.; Nomura, Y.; Oliveira, E.E.; Salgado, V.L.; Zhorov, B.S.; Dong, K. Voltage-gated sodium channels as insecticide targets. Adv. Insect Phys. 2014, 46, 389–433. [Google Scholar]
- Pauron, D.; Barhanin, J.; Amichot, M.; Pralavorio, M.; Berge, J.B.; Lazdunski, M. Pyrethroid receptor in the insect Na+ channel: Alteration of its properties in pyrethroid-resistant flies. Biochem J. 1989, 28, 1673–1677. [Google Scholar] [CrossRef]
- Soderlund, D.M.; Knipple, D.C. The molecular biology of knockdown resistance to pyrethroid insecticides. Insect Biochem. Mol. Biol. 2003, 33, 563–577. [Google Scholar] [CrossRef]
- Chandre, F.; Darriet, F.; Duchon, S.; Finot, L.; Manguin, S.; Carnevale, P.; Guillet, P. Modifications of pyrethroid effects associated with kdr mutation in Anopheles gambiae. Med. Vet. Entomol. 2000, 14, 81–88. [Google Scholar] [CrossRef]
- Gan, S.J.; Leong, Y.Q.; bin Barhanuddin, M.F.H.; Wong, S.T.; Wong, S.F.; Mak, J.W.; Ahmad, R.B. Dengue fever and insecticide resistance in Aedes mosquitoes in Southeast Asia: A review. Parasites Vectors 2021, 14, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Weill, M.; Fort, P.; Berthomieu, A.; Dubois, M.P.; Pasteur, N.; Raymond, M. A novel acetyl-cholinesterase gene in mosquitoes codes for the insecticide target and is non-homologous to the ace gene in Drosophila. Proc. Biol. Sci. 2002, 269, 2007–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weill, M.; Lutfalla, G.; Mogensen, K.; Chandre, F.; Berthomieu, A.; Berticat, C.; Pasteur, N.; Philips, A.; Fort, P.; Raymond, M. Comparative genomics: Insecticide resistance in mosquito vectors. Nature 2003, 423, 136–137. [Google Scholar] [CrossRef] [PubMed]
- Alout, H.; Djogbénou, L.; Berticat, C.; Chandre, F.; Weill, M. Comparison of Anopheles gambiae and Culex pipiens acetylcholinesterase 1 biochemical properties. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 150, 271–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djogbénou, L.; Akogbeto, M.; Chandre, F. Presence of insensitive acetylcholinesterase in wild populations of Culex pipiens quinquefasciatus from Benin. Acta Trop. 2008, 107, 272–274. [Google Scholar] [CrossRef] [PubMed]
- Cole, L.M.; Roush, T.T.; Casida, J.E. Drosophila GABA-gated chloride channel: Modified [3H]EBOB binding site associated with Ala→Ser or Gly mutants of Rdl subunit. Life Sci. 1995, 56, 757–765. [Google Scholar] [CrossRef]
- Du, W.; Awolola, T.S.; Howell, P.; Koekemoer, L.L.; Brooke, B.D. Independent mutations in the Rdl locus confer dieldrin resistance to Anopheles gambiae and An. arabiensis. Insect Mol. Biol. 2005, 14, 179–183. [Google Scholar] [CrossRef]
- Wondji, C.S.; Dabire, R.K.; Tukur, Z.; Irving, H.; Djouaka, R.; Morgan, J.C. Identification and distribution of a GABA receptor mutation conferring dieldrin resistance in the malaria vector Anopheles funestus in Africa. Insect Biochem. Mol. Biol. 2011, 41, 484–491. [Google Scholar] [CrossRef] [Green Version]
- Taylor-Wells, J.; Brooke, B.D.; Bermudez, I.; Jones, A.K. The neonicotinoid imidacloprid, and the pyrethroid deltamethrin, are antagonists of the insect Rdl GABA receptor. J. Neurochem. 2015, 135, 705–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcombe, S.; Thammavong, P.; Luangamath, P.; Chonephetsarath, S.; Phommavanh, N.; Lakeomany, K.; Nilaxay, S.; Rahmani, Z.; Saverton, P.J.; Abdullateef, O.H. Malaria and Dengue Mosquito Vectors from Lao PDR Show a Lack of the rdl Mutant Allele Responsible for Cyclodiene Insecticide Resistance. J. Med. Entomol. 2020, 57, 815–823. [Google Scholar] [CrossRef]
- Amelia-Yap, Z.H.; Tan, T.K.; Prakash, B.K.; Chen, C.D.; Sofian-Azirun, M.; Low, V.L. First report of Rdl mutant alleles in Culex quinquefasciatus (Diptera: Culicidae) in Malaysia. J. Vector Borne Dis. 2020, 57, 359–361. [Google Scholar] [PubMed]
- Kasai, S.; Weerasinghe, I.S.; Shono, T. P450 Monooxygenases are an important mechanism of permethrin resistance in Culex quinquefasciatus say larvae. Arch. Insect Biochem. Physiol. 1998, 37, 47–56. [Google Scholar] [CrossRef]
- Brogdon, W.G.; McAllister, J.C.; Corwin, A.M.; Cordon-Rosales, C. Independent selection of multiple mechanisms for pyrethroid resistance in Guatemalan Anopheles albimanus (Diptera:Culicidae). J. Econ. Entomol. 1999, 92, 298–302. [Google Scholar] [CrossRef]
- Vulule, J.M.; Beach, R.F.; Atieli, F.K.; McAllister, J.C.; Brogdon, W.G.; Roberts, J.M.; Mwangi, R.W.; Hawley, W.A. Elevated oxidase and esterase levels associated with permethrin tolerance in Anopheles gambiae from Kenyan villages using permethrin-impregnated nets. Med. Vet. Entomol. 1999, 13, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Wondji, C.S.; Irving, H.; Morgan, J.; Lobo, N.F.; Collins, F.H.; Hunt, R.H.; Coetzee, M.; Hemingway, J.; Ranson, H. Two duplicated P450 genes are associated with pyrethroid resistance in Anopheles funestus, a major malaria vector. Genome Res. 2009, 19, 452–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wondji, C.S.; Coleman, M.; Kleinschmidt, I.; Mzilahowa, T.; Irving, H.; Ndula, M. Impact of pyrethroid resistance on operational malaria control in Malawi. Proc. Natl. Acad. Sci. USA 2012, 109, 19063–19070. [Google Scholar] [CrossRef] [Green Version]
- David, J.-P.; Ismail, H.M.; Chandor-Proust, A.; Paine, M.J.I. Role of cytochrome P450s in insecticide resistance: Impact on the control of mosquito-borne diseases and use of insecticides on Earth. Phil. Trans. R. Soc. B 2013, 368, 20120429. [Google Scholar] [CrossRef]
- Vontas, J.; Katsavou, E.; Mavridis, K. Cytochrome P450-based metabolic insecticide resistance in Anopheles and Aedes mosquito vectors: Muddying the waters. Pestic. Biochem. Physiol. 2020, 170, 104666. [Google Scholar] [CrossRef]
- Yang, T.; Li, T.; Feng, X.; Li, M.; Liu, S.; Liu, N. Multiple cytochrome P450 genes: Conferring high levels of permethrin resistance in mosquitoes, Culex quinquefasciatus. Sci. Rep. 2021, 11, 9041. [Google Scholar] [CrossRef] [PubMed]
- Lycett, G.J.; McLaughlin, L.A.; Ranson, H.; Hemingway, J.; Kafatos, F.C.; Loukeris, T.G.; Paine, M.J.I. Anopheles gambiae P450 reductase is highly expressed in oenocytes and in vivo knockdown increases permethrin susceptibility. Insect Mol. Biol. 2006, 15, 321–327. [Google Scholar] [CrossRef]
- Li, T.; Cao, C.; Yang, T.; Zhang, L.; He, L.; Xi, Z.; Bian, G.; Liu, N. A G-protein-coupled receptor regulation pathway in cytochrome P450-mediated permethrin-resistance in mosquitoes, Culex quinquefasciatus. Sci. Rep. 2016, 5, 17772. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, N. Regulation of P450-mediated permethrin resistance in Culex quinquefasciatus by the GPCR/Gas/AC/cAMP/PKA signaling cascade. Biochem. Biophys. Rep. 2017, 12, 12–19. [Google Scholar]
- Hemingway, J.; Hawkes, N.J.; McCarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Grant, D.F.; Matsumura, F. Glutathione S-transferase 1 and 2 in susceptible and insecticide resistant Aedes aegypti. Pestic. Biochem. Physiol. 1989, 33, 132–143. [Google Scholar] [CrossRef]
- Prapanthadara, L.; Hemingway, J.; Ketterman, A.J. Partial purification and characterization of glutathione S-transferase involved in DDT resistance from the mosquito Anopheles gambiae. Pest. Biochem. Physiol. 1993, 47, 119–133. [Google Scholar] [CrossRef]
- Ortelli, F.; Rossiter, L.C.; Vontas, J.; Ranson, H.; Hemingway, J. Heterologous expression of four glutathione transferase genes genetically linked to a major insecticide-resistance locus from the malaria vector Anopheles gambiae. Biochem. J. 2003, 373, 957–963. [Google Scholar] [CrossRef]
- Ding, Y.; Ortelli, F.; Rossiter, L.C.; Hemingway, J.; Ranson, H. The Anopheles gambiae glutathione transferase supergene family: Annotation, phylogeny and expression profiles. BMC Genom. 2003, 4, 35. [Google Scholar] [CrossRef] [Green Version]
- Lumjuan, N.; MaCarroll, L.; Prapanthadara, L.A.; Hemingway, J.; Ranson, H. Elevated activity of an Epsilon class glutathione S-transferase confers DDT resistance in the dengue vector, Aedes aegypti. Insect Biochem. Mol. Biol. 2005, 35, 861–871. [Google Scholar] [CrossRef]
- Lumjuan, N.; Rajatileka, S.; Changsom, D.; Wicheer, J.; Leelapat, P.; Prapanthadara, L.; Somboon, P.; Lycett, G.; Ranson, H. The role of the Aedes aegypti Epsilon glutathione transferases in conferring resistance to DDT and pyrethroid insecticides. Insect Biochem. Mol. Biol. 2011, 41, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Marcombe, S.; Poupardin, R.; Darriet, F.; Reynaud, S.; Bonnet, J.; Strode, C.; Brengues, C.; Yebakima, A.; Ranson, H.; Corbel, V.; et al. Exploring the molecular basis of insecticide resistance in the dengue vector Aedes aegypti: A case study in Martinique Island (French West Indies). BMC Genom. 2009, 10, 494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunaratne, S.H.; Hemingway, J.; Jayawardena, K.G.; Dassanayaka, V.; Vaughan, A. Kinetic and molecular differences in the amplified and non-amplified esterases from insecticide-resistant and susceptible Culex quinquefasciatus mosquitoes. J. Biol. Chem. 1995, 270, 31124–31128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ketterman, A.J.; Jayawardena, K.; Hemingway, J. Purification and characterization of a carboxylesterase involved in insecticide resistance from the mosquito Culex quinquefasciatus. Biochem. J. 1992, 287, 355–360. [Google Scholar] [CrossRef]
- Vaughan, A.; Hemingway, J. Mosquito carboxylesterase Est alpha 2(1) (A2). Cloning and sequence of the full-length cDNA for a major insecticide resistance gene worldwide in the mosquito Culex quinquefasciatus. J. Biol. Chem. 1995, 270, 17044–17049. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, A.; Hawkes, N.; Hemingway, J. Co-amplification explains linkage disequilibrium of two mosquito esterase genes in insecticide-resistant Culex quinquefasciatus. Biochem. J. 1997, 325, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Bisset, J.; Rodriguez, M.; Soca, A.; Pasteur, N.; Raymond, M. Cross-resistance to pyrethroid and organophosphorus insecticides in the southern house mosquito (Diptera:Culicidae) from Cuba. J. Med. Entomol. 1997, 34, 244–246. [Google Scholar] [CrossRef]
- Liu, H.; Cupp, E.W.; Micher, K.M.; Guo, A.; Liu, N. Insecticide resistance and cross-resistance in Alabama and Florida strains of Culex quinquefasciatus. J. Med. Entomol. 2004, 41, 408–413. [Google Scholar] [CrossRef]
- Yunta, C.; Hemmings, K.; Stevenson, B.; Koekemoer, L.L.; Matambo, T.; Pignatelli, P.; Voice, M.; Nász, S.; Paine, M.J.I. Cross-resistance profiles of malaria mosquito P450s associated with pyrethroid resistance against WHO insecticides. Pestic. Biochem. Physiol. 2019, 161, 61–67. [Google Scholar] [CrossRef]
- Guillemaud, T.; Lenormand, T.; Bourguet, D.; Chevillon, C.; Pasteur, N.; Raymond, M. Evolution of Resistance in Culex pipiens: Allele Replacement and Changing Environment. Evolution 1998, 52, 443–453. [Google Scholar] [CrossRef]
- Schechtman, H.; Souza, M.O. Costly inheritance and the persistence of insecticide resistance in Aedes aegypti populations. PLoS ONE 2015, 10, e0123961. [Google Scholar] [CrossRef] [PubMed]
- Laxmishree, C.; Nandita, S. Botanical pesticides—A major alternative to chemical pesticides: A review. Int. J. Life Sci. 2017, 5, 722–729. [Google Scholar]
- Thakore, Y. The biopesticide market for global agricultural use. Ind. Biotechnol. 2006, 2, 194–208. [Google Scholar] [CrossRef]
- Prabha, S.; Yadav, A.; Kumar, A.; Yadav, A.; Yadav, H.K.; Kumar, S.; Kumar, R. Biopesticides—An alternative and eco-friendly source for the control of pests in agricultural crops. Plant Arch. 2016, 16, 902–906. [Google Scholar]
- Ohia, C.M.D.; Ana, G.R.E.E. Bio-insecticides: The one-health response to mosquito-borne diseases of public health importance. J. Biol. Agric. Healthc. 2015, 5, 22–26. [Google Scholar]
- Dara, S.K. Insect Resistance to Biopesticides. Available online: https://ucanr.edu/blogs/blogcore/postdetail.cfm?postnum=25819 (accessed on 23 November 2021).
- Tyagi, B.K. Advances in Vector Mosquito Control Technologies, with Particular Reference to Herbal Products. In Herbal Insecticides, Repellents and Biomedicines: Effectiveness and Commercialization; Veer, V., Gopalakrishnan, R., Eds.; Springer: New Delhi, India, 2016; pp. 1–11. [Google Scholar]
- Shaalan, E.A.S.; Canyonb, D.; Younesc, M.W.F.; Abdel-Wahaba, H.; Mansoura, A.H. A review of botanical phytochemicals with mosquitocidal potential. Environ. Int. 2005, 3, 1149–1166. [Google Scholar] [CrossRef] [PubMed]
- Souto, A.L.; Sylvestre, M.; Tölke, E.D.; Tavares, J.F.; Barbosa-Filho, J.M.; Cebrián-Torrejón, G. Plant-Derived Pesticides as an Alternative to Pest Management and Sustainable Agricultural Production: Prospects, Applications and Challenges. Molecules 2021, 26, 4835. [Google Scholar] [CrossRef]
- Sharma, P.; Mohan, L.; Srivastava, C.N. Phytoextract-induced developmental deformities in malaria vector. Bioresour. Technol. 2006, 97, 1599–1604. [Google Scholar] [CrossRef] [PubMed]
- Al-Mekhlafi, F.A. Larvicidal, ovicidal activities and histopathological alterations induced by Carum copticum (Apiaceae) extract against Culex pipiens (Diptera: Culicidae). Saudi J. Biol. Sci. 2018, 25, 52–56. [Google Scholar] [CrossRef]
- Senthil-Nathan, S. A review of resistance mechanisms of synthetic insecticides and botanicals, phytochemicals, and essential oils as alternative larvicidal agents against mosquitoes. Front. Physiol. 2020, 10, 1591. [Google Scholar] [CrossRef]
- Syed, Z.; Leal, W.S. Mosquitoes smell and avoid the insect repellent DEET. Proc. Natl. Acad. Sci. USA 2008, 105, 13598–13603. [Google Scholar] [CrossRef] [Green Version]
- Dickens, J.C.; Bohbot, J.D. Mini review: Mode of action of mosquito repellents. Pestic. Biochem. Physiol. 2013, 106, 149–155. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G. Ethnobotanical knowledge on botanical repellents employed in the African region against mosquito vectors—A review. Exp. Parasitol. 2016, 167, 103–108. [Google Scholar] [CrossRef]
- Maia, M.F.; Moore, S.J. Plant-based insect repellents: A review of their efficacy, development and testing. Malar. J. 2011, 10, S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagegowda, D.A. Plant volatile terpenoid metabolism: Biosynthetic genes, transcriptional regulation and subcellular compartmentation. FEBS Lett. 2010, 584, 2965–2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isman, M.B.; JunHyung, T. Inhibition of acetylcholinesterase by essential oils and monoterpenoids: A relevant mode of action for insecticidal essential oils? Biopestic. Int. 2017, 13, 71–78. [Google Scholar]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular targets for components of essential oils in the insect nervous system—A review. Molecules 2018, 23, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavela, R. Essential oils for the development of eco-friendly mosquito larvicides: A review. Ind. Crops Prod. 2015, 76, 174–187. [Google Scholar] [CrossRef]
- Houghton, P.J.; Ren, Y.; Howes, M.J. Acetylcholinesterase inhibitors from plants and fungi. Nat. Prod. Rep. 2006, 23, 181–199. [Google Scholar] [CrossRef]
- Abdelgaleil, S.A.; Mohamed, M.I.; Badawy, M.E.; El-arami, S.A. Fumigant and contact toxicities of monoterpenes to Sitophilus oryzae (L.) and Tribolium castaneum (Herbst) and their inhibitory effects on acetylcholinesterase activity. J. Chem. Ecol. 2009, 35, 518–535. [Google Scholar] [CrossRef]
- Hideyuki, T.; Mitsuo, M. Inhibition of acetylcholinesterase activity by essential oil from Bergamot. Koryo Terupen oyobi Seiyu Kagaku ni Kansuru Toronkai Koen Yoshishu 2001, 45, 435–436. [Google Scholar]
- Enan, E. Insecticidal activity of essential oils: Octopaminergic sites of action. Comp. Biochem. Physiol. C Toxicol. 2001, 130, 325–337. [Google Scholar] [CrossRef]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef] [PubMed]
- Li, A.S.; Iijima, A.; Huang, J.; Li, Q.X.; Chen, Y. Putative Mode of Action of the Monoterpenoids Linalool, Methyl Eugenol, Estragole, and Citronellal on Ligand-Gated Ion Channels. Engineering 2020, 6, 541–545. [Google Scholar] [CrossRef]
- Bloomquist, J.R. Chloride channels as tools for developing selective insecticides. Arch. Insect Biochem. Physiol. 2003, 54, 145–156. [Google Scholar] [CrossRef]
- Bekele, D. Review on insecticidal and repellent activity of plant products for malaria mosquito control. Biomed. Res. Rev. 2018, 2, 1–7. [Google Scholar] [CrossRef]
- Jaenson, T.G.; Pålsson, K.; Borg-Karlson, A.K. Evaluation of extracts and oils of mosquito (Diptera: Culicidae) repellent plants from Sweden and Guinea-Bissau. J. Med. Entomol. 2006, 43, 113–119. [Google Scholar] [CrossRef]
- Gillij, Y.G.; Gleiser, R.M.; Zygadlo, J.A. Mosquito repellent activity of essential oils of aromatic plants growing in Argentina. Bioresour. Technol. 2008, 99, 2507–2515. [Google Scholar] [CrossRef]
- Pandey, S.K.; Upadhyay, S.; Tripathi, A.K. Insecticidal and repellent activities of thymol from the essential oil of Trachyspermum ammi (Linn) Sprague seeds against Anopheles stephensi. Parasitol. Res. 2009, 105, 507–512. [Google Scholar] [CrossRef]
- Park, B.S.; Choi, W.S.; Kim, J.H.; Kim, K.H.; Lee, S.E. Monoterpenes from thyme (Thymus vulgaris) as potential mosquito repellents. J. Am. Mosq. Control Assoc. 2005, 21, 80–83. [Google Scholar] [CrossRef]
- Pavela, R.; Vrchotová, N.; Tříska, J. Mosquitocidal activities of thyme oils (Thymus vulgaris L.) against Culex quinquefasciatus (Diptera: Culicidae). Parasitol. Res. 2009, 105, 1365–1370. [Google Scholar] [CrossRef] [PubMed]
- Pitarokili, D.; Michaelakis, A.; Koliopoulos, G.; Koliopoulos, G.; Giatropoulos, A.; Tzakouet, O. Chemical composition, larvicidal evaluation, and adult repellency of endemic Greek Thymus essential oils against the mosquito vector of West Nile virus. Parasitol. Res. 2011, 109, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Michaelakis, A.; Theotokatos, S.A.; Koliopoulos, G.; Chorianopoulos, N.G. Essential oils of Satureja species: Insecticidal effect on Culex pipiens larvae (Diptera: Culicidae). Molecules 2007, 12, 2567–2578. [Google Scholar] [CrossRef] [PubMed]
- Evergetis, E.; Bellini, R.; Balatsos, G.; Michaelakis, A.; Carrieri, M.; Veronesi, R.; Papachristos, D.P.; Puggioli, A.; Kapsaski-Kanelli, V.-N.; Haroutounian, S.A. From Bio-Prospecting to Field Assessment: The Case of Carvacrol Rich Essential Oil as a Potent Mosquito Larvicidal and Repellent Agent. Front. Ecol. Evol. 2018, 6, 204. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.S.; Liu, J.Y.; Tsai, K.H.; Chen, W.J.; Chang, S.T. Chemical composition and mosquito larvicidal activity of essential oils form leaves of different Cinnamonum osmophloem provenances. J. Agric. Food Chem. 2004, 52, 4395–4400. [Google Scholar] [CrossRef] [PubMed]
- Tabari, M.A.; Youssefi, M.R.; Esfandiari, A.; Benelli, G. Toxicity of β-citronellol, geraniol and linalool from Pelargonium roseum essential oil against the West Nile and filariasis vector Culex pipiens (Diptera: Culicidae). Res. Vet. Sci. 2017, 114, 36–40. [Google Scholar] [CrossRef]
- Andrade-Ochoa, S.; Sánchez-Aldana, D.; Chacón-Vargas, K.F.; Rivera-Chavira, B.E.; Sánchez-Torres, L.E.; Camacho, A.D.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. Oviposition Deterrent and Larvicidal and Pupaecidal Activity of Seven Essential Oils and their Major Components against Culex quinquefasciatus Say (Diptera: Culicidae): Synergism–antagonism Effects. Insects 2018, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Govindarajan, M.; Benelli, G. Artemisia absinthium-borne compounds as novel larvicides: Effectiveness against six mosquito vectors and acute toxicity on non-target aquatic organisms. Parasitol. Res. 2016, 115, 4649–4661. [Google Scholar] [CrossRef] [Green Version]
- Vera, S.S.; Zambrano, D.F.; Méndez-Sanchez, S.C.; Rodríguez-Sanabria, F.; Stashenko, E.E.; Luna, J.E.D. Essential oils with insecticidal activity against larvae of Aedes aegypti (Diptera: Culicidae). Parasitol. Res. 2014, 113, 2647–2654. [Google Scholar] [CrossRef]
- Warikoo, R.; Wahab, N.; Kumar, S. Oviposition-altering and ovicidal potentials of five essential oils against female adults of the dengue vector, Aedes aegypti L. Parasitol. Res. 2011, 109, 1125–1131. [Google Scholar] [CrossRef]
- Manh, H.D.; Hue, D.T.; Hieu, N.T.T.; Tuyen, D.T.T.; Tuyet, O.T. The Mosquito larvicidal activity of essential oils from Cymbopogon and Eucalyptus Species in Vietnam. Insects 2020, 11, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnankiné, O.; Bassolé, I.H.N. Essential oils as an alternative to pyrethroids’ resistance against Anopheles species complex giles (Diptera: Culicidae). Molecules 2017, 22, 1321. [Google Scholar] [CrossRef] [Green Version]
- Bassolé, I.H.N.; Guelbeogo, W.M.; Nébié, R.; Costantini, C.; Sagnon, N.; Kabore, Z.I.; Traoré, S.A. Ovicidal and larvicidal activity against Aedes aegypti and Anopheles gambiae complex mosquitoes of essential oils extracted from three spontaneous plants of Burkina Faso. Parassitologia 2003, 45, 23–26. [Google Scholar] [PubMed]
- Cantrell, C.L.; Jones, A.M.; Ali, A. Isolation and Identification of Mosquito (Aedes aegypti) Biting-Deterrent Compounds from the Native American Ethnobotanical Remedy Plant Hierochloë odorata (Sweetgrass). J. Agric. Food Chem. 2016, 64, 8352–8358. [Google Scholar] [CrossRef]
- Abbassy, M.A.; Abdelgaleil, S.A.; Rabie, R.Y. Insecticidal and synergistic effects of Majorana hortensis essential oil and some of its major constituents. Entomol. Exp. Appl. 2009, 131, 225–232. [Google Scholar] [CrossRef]
- Osanloo, M.; Sedaghat, M.M.; Esmaeili, F.; Amani, A. Larvicidal activity of essential oil of Syzygium aromaticum (Clove) in comparison with its major constituent, eugenol, against Anopheles stephensi. J. Arthropod Borne Dis. 2018, 12, 361–369. [Google Scholar]
- Dhinakaran, S.R.; Mathew, N.; Munusamy, S. Synergistic terpene combinations as larvicides against the Dengue vector Aedes aegypti Linn. Drug Dev. Res. 2019, 80, 791–799. [Google Scholar] [CrossRef]
- Odalo, J.O.; Omolo, M.O.; Malebo, H.; Angira, J.; Njeru, P.M.; Ndiege, I.O.; Hassanali, A. Repellency of essential oils of some plants from the Kenyan coast against Anopheles gambiae. Acta Trop. 2005, 95, 210–218. [Google Scholar] [CrossRef]
- Schmutterer, H. Properties and potential of natural pesticides from the neem tree, Azadirachta indica. Annu. Rev. Entomol. 1990, 35, 271–297. [Google Scholar] [CrossRef]
- Campos, E.V.; de Oliveira, J.L.; Pascoli, M.; de Lima, R.; Fraceto, L.F. Neem oil and crop protection: From now to the future. Front. Plant Sci. 2016, 7, 1494. [Google Scholar] [CrossRef] [Green Version]
- Govindchari, T.R. Chemistry and biological investigation on Azadirachta indica (the neem tree). Curr. Sci. 1992, 63, 117–122. [Google Scholar]
- Mordue, A.J.; Blackwell, A. Azadirachtin: An update. J. Insect Physiol. 1993, 11, 903–924. [Google Scholar] [CrossRef]
- Sidhu, O.P.; Kumar, V.; Behl, H.M. Variability in neem (Azadirachta indica) with respect to azadirachtin content. J. Agric. Food Chem. 2003, 51, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.D. Azadirachtin, a scientific gold mine. Bioorg. Med. Chem. 2009, 17, 4096–4105. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Murugan, K.; Panneerselvam, C.; Madhiyazhagan, P.; Conti, B.; Nicoletti, M. Old ingredients for a new recipe? Neem cake, a low-cost botanical by-product in the fight against mosquito-borne diseases. Parasitol. Res. 2015, 114, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, N. Neem: Present status and future prospects. Plant Arch. 2008, 8, 17–22. [Google Scholar]
- Beckage, N.E. Insect growth regulators. In Biological and Biotechnological Control of Insect Pests; Rechcigl, J.E., Rechcigl, N.A., Eds.; CRC Press LLC: Boca Raton, FL, USA, 2000; pp. 123–137. [Google Scholar]
- Mulla, M.S.; Su, M. Activity and biological effects of neem products against arthropods of medical and veterinary importance. J. Am. Mosq. Control Assoc. 1999, 15, 133–152. [Google Scholar] [PubMed]
- Nisbet, A.J. Azadirachtin from the neem tree Azadirachta indica: Its action against insects. An. Soc. Entomol. Bras. 2000, 29, 615–632. [Google Scholar]
- Schmutterer, H. The Neem Tree, Azadirachta indica (A. Juss) and Other Meliaceous Plants: Source of Unique Natural Products for Integrated Pest Management, Medicine, Industry and Other Purposes; VCH: Weinheim, Germany, 1995; p. 696. [Google Scholar]
- Lucantoni, L.; Giusti, F.; Cristofaro, M.; Pasqualini, L.; Esposito, F.; Lupetti, P.; Habluetzel, A. Effects of a neem extract on blood feeding, oviposition and oocyte ultrastructure in Anopheles stephensi Liston (Diptera: Culicidae). Tissue Cell 2006, 38, 361–371. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides, deterrents and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [Green Version]
- Dua, V.K.; Pandey, A.C.; Raghavendra, K.; Gupta, A.; Sharma, T.; Dash, A.P. Larvicidal activity of neem oil (Azadirachta indica) formulation against mosquitoes. Malar. J. 2009, 8, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayinde, A.A.; Morakinyo, O.M.; Sridhar, M.K.C. Repellency and larvicidal activities of Azadirachta indica seed oil on Anopheles gambiae in Nigeria. Heliyon 2020, 6, e03920. [Google Scholar] [CrossRef] [PubMed]
- Maheswaran, R.; Ignacimuthu, S. A novel herbal formulation against dengue vector mosquitoes Aedes aegypti and Aedes albopictus. Parasitol. Res. 2012, 110, 1801–1813. [Google Scholar] [CrossRef] [PubMed]
- Senthil-Nathan, S.; Kalaivani, K.; Muruganb, K. Effects of neem limonoids on the malaria vector Anopheles stephensi Liston (Diptera: Culicidae). Acta Trop. 2005, 96, 47–55. [Google Scholar] [CrossRef]
- Vatandoost, H.; Vaziri, V.M. Larvicidal activity of a neem tree extract (Neemarin) against mosquito larvae in the Islamic Republic of Iran. East. Mediterr. Health J. 2004, 10, 573–581. [Google Scholar] [CrossRef]
- Grdiša, M.; Gršić, K. Botanical insecticides in plant protection. Agric. Conspec. Sci. 2013, 78, 85–93. [Google Scholar]
- Pavela, R. History, presence and perspective of using plant extracts as commercial botanical insecticides and farm products for protection against insects—A review. Plant Prot. Sci. 2016, 52, 229–241. [Google Scholar]
- Osimitz, T.G.; Sommers, N.; Kingston, R. Human exposure to insecticide products containing pyrethrins and piperonyl butoxide (2001–2003). Food Chem. Toxicol. 2009, 47, 1406–1415. [Google Scholar] [CrossRef]
- Duchon, S.; Bonnet, J.; Marcombe, S.; Zaim, M.; Corbel, V. Pyrethrum: A mixture of natural pyrethrins has potential for malaria vector control. J. Med. Entomol. 2009, 46, 516–522. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Wang, Q.; Xu, P.; Andreazza, F.; Valbon, W.R.; Bandason, E.; Dong, K. A dual-target molecular mechanism of pyrethrum repellency against mosquitoes. Nat. Commun. 2021, 12, 2553. [Google Scholar] [CrossRef]
- Gutiérrez-Grijalva, E.P.; López-Martínez, L.X.; Contreras-Angulo, L.A.; Elizalde-Romero, C.A.; Heredia, J.B. Plant Alkaloids: Structures and Bioactive Properties. In Plant-Derived Bioactives; Swamy, M.K., Ed.; Springer: Singapore, 2020; pp. 85–117. [Google Scholar]
- Ahmed, N.; Alam, M.; Saeed, M.; Ullah, H.; Iqbal, T.; Al-Mutairi, K.A.; Shahjeer, K.; Ullah, R.; Ahmed, S.; Ahmed, N.A.A.H.; et al. Botanical insecticides are a non-toxic alternative to conventional pesticides in the control of insects and pests. In Global Decline of Insects; El-Shafie, H., Ed.; IntechOpen: London, UK, 2021. [Google Scholar]
- Secoy, D.M.; Smith, A.E. Use of plants in control of agricultural and domestic pests. Econ. Bot. 1983, 37, 28–57. [Google Scholar] [CrossRef]
- Chowański, S.; Adamski, Z.; Marciniak, P.; Rosiński, G.; Büyükgüzel, E.; Büyükgüzel, K.; Bufo, S.A. A review of bioinsecticidal activity of Solanaceae alkaloids. Toxins 2016, 8, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balachandran, C.; Anbalagan, S.; Kandeepan, C.; Nagendran, N.A.; Jayakumar, M.; Abd Allah, E.F.; Alqarawi, A.A.; Hashem, A.; Baskar, K. Molecular docking studies of natural alkaloids as acetylcholinesterase (ache1) inhibitors in Aedes aegypti. J. Asia Pac. Entomol. 2021, 24, 645–652. [Google Scholar] [CrossRef]
- Elimam, A.M.; Elmalik, K.H.; Ali, F.S. Larvicidal, adult emergence inhibition and oviposition deterrent effects of foliage extract from Ricinus communis L. against Anopheles arabiensis and Culex quinquefasciatus in Sudan. Trop. Biomed. 2009, 26, 130–139. [Google Scholar]
- Wachira, S.W.; Omar, S.; Jacob, J.W.; Wahome, M.; Alborn, H.T.; Spring, D.R.; Masiga, D.; Torto, B. Toxicity of six plant extracts and two pyridone alkaloids from Ricinus communis against the malaria vector Anopheles gambiae. Parasites Vectors 2014, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.I.; Ahn, Y.J. Larvicidal activity of lignans and alkaloid identified in Zanthoxylum piperitum bark toward insecticide-susceptible and wild Culex pipiens pallens and Aedes aegypti. Parasites Vectors 2017, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.L.; Liu, Q.Z.; Du, S.S.; Deng, Z.W. Mosquito larvicidal activity of alkaloids and limonoids derived from Evodia rutaecarpa unripe fruits against Aedes albopictus (Diptera: Culicidae). Parasitol. Res. 2012, 111, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Velu, K.; Elumalai, D.; Hemalatha, P.; Babu, M.; Janaki, A.; Kaleena, P.K. Phytochemical screening and larvicidal activity of peel extracts of Arachis hypogaea against chikungunya and malarial vectors. Int. J. Mosq. Res. 2015, 2, 1–8. [Google Scholar]
- Jufri, M.; Irmayanti, E.; Gozan, M. Formulation of tobacco-based mosquito repellent to avoid dengue fever. Int. J. Pharmtech. Res. 2016, 9, 140–145. [Google Scholar]
- Perumalsamy, H.; Jang, M.J.; Kim, J.R.; Kadarkarai, M.; Ahn, Y.J. Larvicidal activity and possible mode of action of four flavonoids and two fatty acids identified in Millettia pinnata seed toward three mosquito species. Parasites Vectors 2015, 8, 237. [Google Scholar] [CrossRef] [Green Version]
- Ningrum, D.S.; Wijayanti, S.P.M.; Kuswanto, K. Mosquito larvacidal activity of Zingiber Montanum rhizome extract against Aedes aegypti larvae. Balaba 2019, 15, 33–40. [Google Scholar] [CrossRef]
- Yenesew, A.; Twinomuhwezi, H.; Kabaru, J.M.; Akala, H.M.; Kiremire, B.T.; Heydenreich, M.; Peter, M.G.; Eyase, F.L.; Waters, N.C.; Walsh, D.S. Antiplasmodial and larvicidal flavonoids from Derris trifoliata. Bull. Chem. Soc. Ethiop. 2009, 23, 409–414. [Google Scholar] [CrossRef]
- Zubairi, S.I.; Sarmidi, M.R.; Aziz, R.A. A preliminary study on mosquito larvicidal efficacy of rotenone extracted from Malaysia Derris sp. J. Teknol. 2015, 76, 275–279. [Google Scholar] [CrossRef] [Green Version]
- WHO. Guidelines for Laboratory and Field Testing of Mosquito Larvicides; World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- WHO. Guidelines for Testing Mosquito Adulticides for Indoor Residual Spraying and Treatment of Mosquito Nets; World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- WHO. Guidelines for Laboratory and Field-Testing of Long-Lasting Insecticidal Nets; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- WHO. Techniques to Detect Insecticide Resistance Mechanisms (Field and Laboratory Manual); World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- Tehri, K.; Singh, N. The role of botanicals as green pesticides in integrated mosquito management—A review. Int. J. Mosq. Res. 2015, 2, 18–23. [Google Scholar]
- Namias, A.; Jobe, N.B.; Paaijmans, K.P.; Huijben, S. The need for practical insecticide-resistance guidelines to effectively inform mosquito-borne disease control programs. eLife 2021, 10, e65655. [Google Scholar] [CrossRef]
- BioWorks. Available online: https://bioworksinc.com/wp-content/uploads/products/shared/resistance-to-biopesticides.pdf (accessed on 20 January 2022).
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in a high-stakes world. Annu. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Aïzoun, N.; Aïkpon, R.; Azondekon, R.; Gnanguenon, V.; Osse, R.; Padonou, G.G.; AkogbÃeto, M. Centre for Disease Control and Prevention (CDC) bottle bioassay: A real complementary method to World Health Organization (WHO) susceptibility test for the determination of insecticide susceptibility in malaria vectors. J. Parasitol. Vector Biol. 2014, 6, 42–47. [Google Scholar]
- Centers for Disease Control. Available online: https://www.cdc.gov/malaria/resources/pdf/fsp/ir_manual/ir_cdc_bioassay_en.pdf (accessed on 20 January 2022).
- Zhu, K.Y. RNA interference: A powerful tool in entomological research and a novel approach for insect pest management. Insect Sci. 2013, 20, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Airs, P.M.; Bartholomay, L.C. RNA Interference for Mosquito and Mosquito-Borne Disease Control. Insects 2017, 8, 4. [Google Scholar] [CrossRef]
- Zhu, F.; Cui, Y.; Walsh, D.B.; Lavine, L.C. Application of RNAi toward insecticide resistance management. In Short Views on Insect Biochemistry and Molecular Biology, 1st ed.; Chandrasekar, R., Tyagi, B.K., Gui, Z.Z., Reeck, G.R., Eds.; International Book Mission, Academic Publisher: Manhattan, NY, USA, 2014; Chapter 27; Volume 2, pp. 595–619. [Google Scholar]
- Munawar, K.; Alahmed, A.M.; Khalil, S.M.S. Delivery methods for RNAi in mosquito larvae. J. Insect Sci. 2020, 20, 1–8. [Google Scholar] [CrossRef]
- Singh, A.D.; Wong, S.; Ryan, C.P.; Whyard, S. Oral delivery of double-stranded RNA in larvae of the yellow fever mosquito, Aedes aegypti: Implications for pest mosquito control. J. Insect Sci. 2013, 13, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blitzer, E.J.; Vyazunova, I.; Lan, Q. Functional analysis of AeSCP-2 using gene expression knockdown in the yellow fever mosquito, Aedes aegypti. Insect Mol. Biol. 2005, 14, 301–307. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, J.; Zhu, K.Y. Chitosan/double-stranded RNA nanoparticle-mediated RNA interference to silence chitin synthase genes through larval feeding in the African malaria mosquito (Anopheles gambiae). Insect Mol. Biol. 2010, 19, 683–693. [Google Scholar] [CrossRef]
- Mafra-Neto, A.; de Lame, F.M.; Fettig, C.J.; Munson, A.S.; Perring, T.M.; Stelinski, L.L.; Stoltman, L.; Mafra, L.E.J.; Borges, R.; Vargas, R.I. Manipulation of Insect Behavior with Specialized Pheromone and Lure Application Technology (SPLAT®). In Natural Products for Pest Management; Beck, J., Coats, J., Duke, S., Koivunen, M., Eds.; ACS Publications: Washington, DC, USA, 2013; Chapter 4; pp. 31–58. [Google Scholar]
- Schorkopf, D.L.P.; Spanoudis, C.G.; Mboera, L.E.; Mafra-Neto, A.; Ignell, R.; Dekker, T. Combining attractants and larvicides in biodegradable matrices for sustainable mosquito vector control. PLoS Negl. Trop. Dis. 2016, 10, e0005043. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B.; Miresmailli, S.; Machial, C. Commercial opportunities for pesticides based on plant essential oils in agriculture, industry and consumer products. Phytochem. Rev. 2011, 10, 197–204. [Google Scholar] [CrossRef]
- Specos, M.M.; García, J.J.; Tornesello, J.; Marino, P.; Vecchia, M.D.; Tesoriero, M.D.; Hermida, L.G. Microencapsulated citronella oil for mosquito repellent finishing of cotton textiles. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Grancaric, A.M.; Laird, K.; Botteri, L.; Shen, J.; Laatikainen, K. Microencapsulation for improved mosquitoes’ repellent efficacy of cotton fabrics. IOP Conf. Ser. Mater. Sci. Eng. 2020, 827, 012056. [Google Scholar] [CrossRef]
- Specos, M.M.; Garcia, J.J.; Gutierrez, A.C.; Hermida, L.G. Application of microencapsulated biopesticides to improve repellent finishing of cotton fabrics. J. Text. Inst. 2017, 108, 1454–1460. [Google Scholar] [CrossRef]
- Pardini, F.; Iregui, Á.; Faccia, P.; Amalvy, J.; González, A.; Irusta, L. Development and characterization of electrosprayed microcaspules of poly ε-caprolactone with citronella oil for mosquito-repellent application. Int. J. Polym. Anal. Charact. 2021, 26, 497–516. [Google Scholar] [CrossRef]
- Sakulkua, U.; Nuchuchuaa, O.; Uawongyartb, N.; Puttipipatkhachornc, S.; Soottitantawatd, A.; Ruktanonchaia, U. Characterization and mosquito repellent activity of citronella oil nanoemulsion. Int. J. Pharm. Nanotechnol. 2009, 372, 105–111. [Google Scholar] [CrossRef]
- Pant, M.; Dubey, S.; Raza, S.K.; Patanjali, P.K. Encapsulation of neem and karanja oil mixture for synergistic as well as larvicidal activity for mosquito control. J. Sci. Ind. Res. 2012, 71, 348–352. [Google Scholar]
- Schleier, J.J., III; Peterson, R.K.D. Pyrethrins and pyrethroid insecticides. In Green Trends in Insect Control; Lopez, O., Fernandez-Bolanos, J., Eds.; Royal Society of Chemistry: Cambridge, UK, 2011; Chapter 3; pp. 94–131. [Google Scholar]
- Veronesi, R.; Gentile, G.; Carrieri, M.; Maccagnani, B.; Stermieri, L.; Bellini, R. Seasonal pattern of daily activity of Aedes caspius, Aedes detritus, Culex modestus, and Culex pipiens in the Po Delta of northern Italy and significance for vector-borne disease risk assessment. J. Vector Ecol. 2012, 37, 49–61. [Google Scholar] [CrossRef]
- Silva, L.L.S.; Silva, S.C.C.; de Oliveira, A.P.S.; Nascimento, J.D.S.; Silva, E.O.; Coelho, L.C.B.B.; Neto, P.J.R.; Navarro, D.M.D.A.F.; Napoleão, T.H.; Paiva, P.M.G. Effects of a solid formulation containing lectin-rich fraction of Moringa oleifera seeds on egg hatching and development of Aedes aegypti larvae. Acta Trop. 2021, 214, 105789. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Şengül Demirak, M.Ş.; Canpolat, E. Plant-Based Bioinsecticides for Mosquito Control: Impact on Insecticide Resistance and Disease Transmission. Insects 2022, 13, 162. https://doi.org/10.3390/insects13020162
Şengül Demirak MŞ, Canpolat E. Plant-Based Bioinsecticides for Mosquito Control: Impact on Insecticide Resistance and Disease Transmission. Insects. 2022; 13(2):162. https://doi.org/10.3390/insects13020162
Chicago/Turabian StyleŞengül Demirak, Meryem Ş., and Emel Canpolat. 2022. "Plant-Based Bioinsecticides for Mosquito Control: Impact on Insecticide Resistance and Disease Transmission" Insects 13, no. 2: 162. https://doi.org/10.3390/insects13020162