
A New Species of Ascodipteron (Diptera: Hippoboscidae) from China Based on Morphology and DNA Barcodes †


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Collection and Preparation
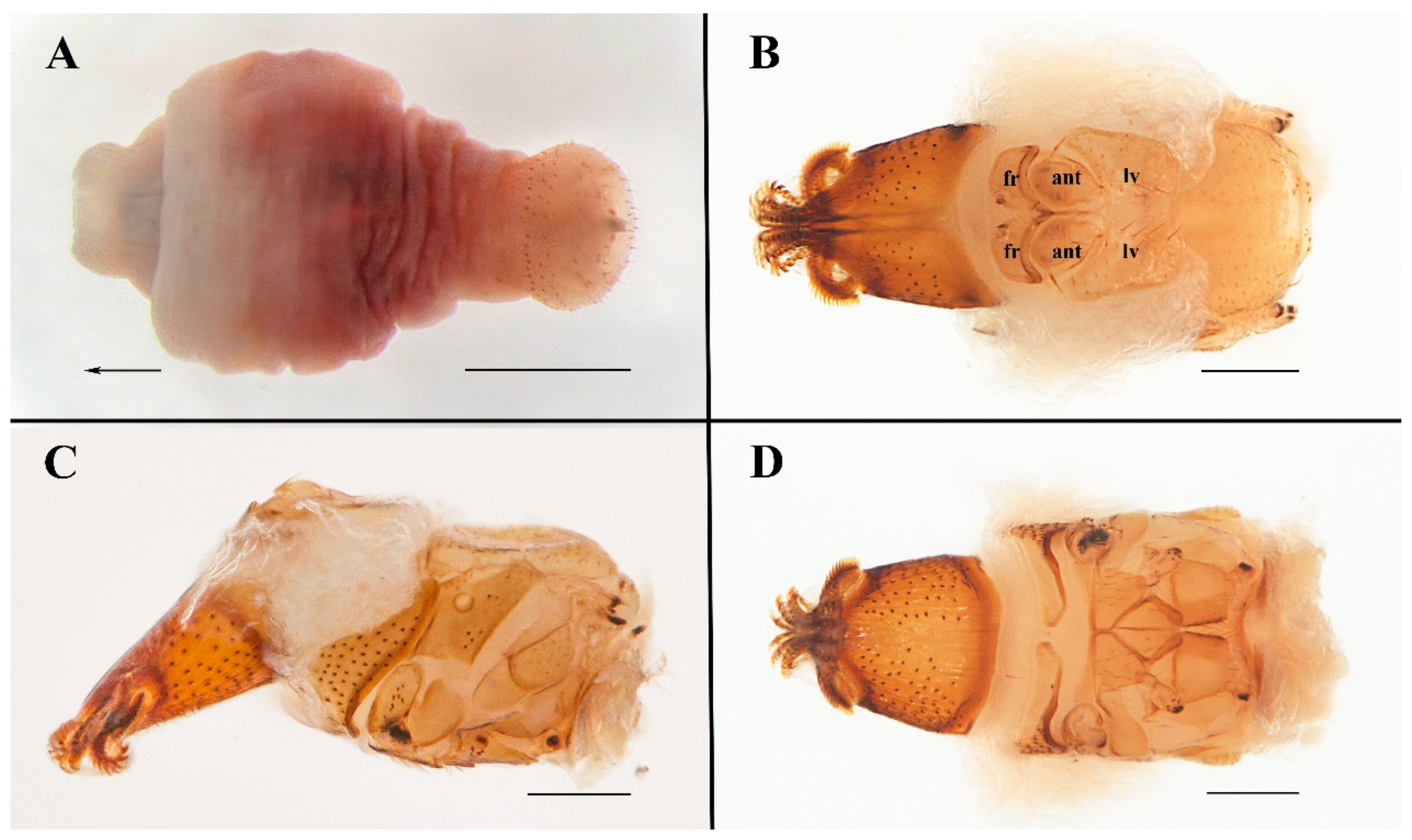
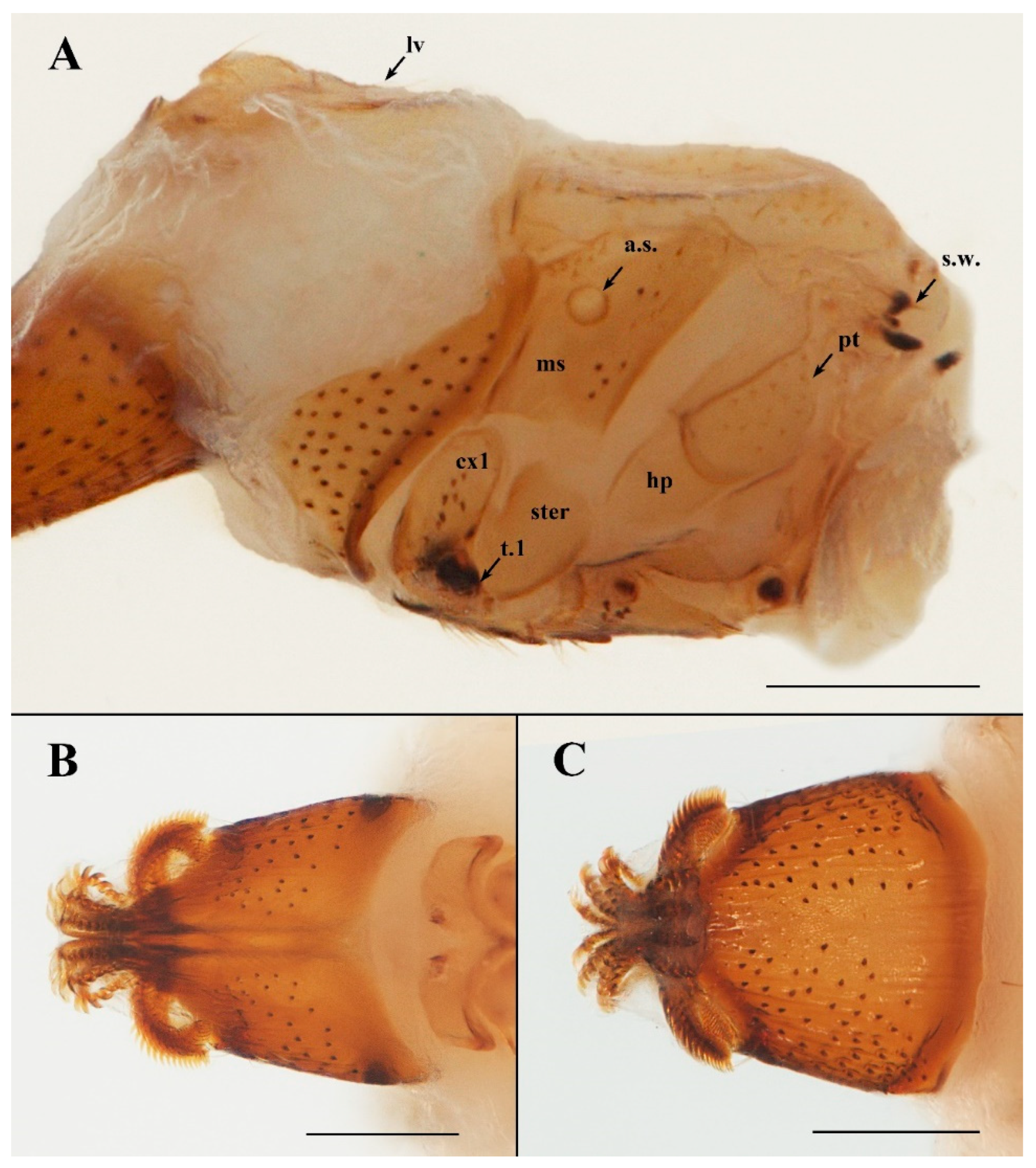
2.2. Specimen Imaging, Measurements, and Terminology
2.3. DNA Extraction, Amplification, Sequencing and Sequence Editing
2.4. DNA Sequence Analysis
3. Results
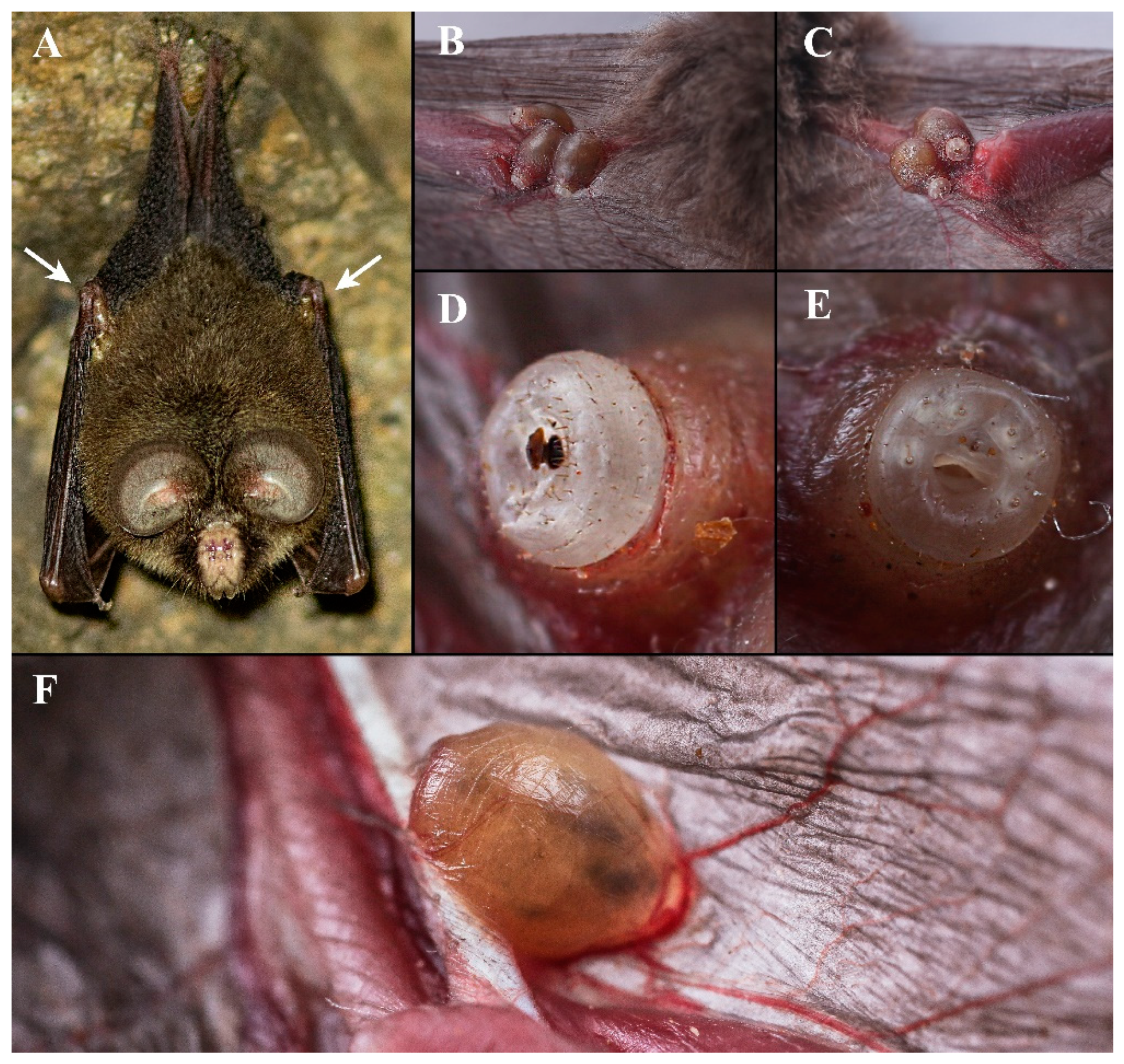
3.1. Taxonomy
- [Couplets 1–12 as in Hastriter (2007) [3]]
- 13 Spiniform setae on labial theca large and pigmented. MSS comprised two or more setae. R1 may or may not be present ...........................................................................................14
- – Spiniform setae smaller and not pigmented; MSS comprised one seta, or none .........17
- 14 R1–R5 present and complete .................................................................................................15
- – R1–R5 absent or incomplete .................................................................................................16
- 15 Labial theca dorsally with 25–30 lightly pigmented peg-like spiniform setae. Lateral vertex without fold or reinforcement in lateral portion of sclerite. [Hipposideros spp., usually on wing, SE China to Solomon Islands.].......A. phyllorhinae Adensamer, 1896
- – Labial theca dorsally with ca. 50+ normally pigmented peg-like spiniform setae. Lateral vertex with longitudinal fold or reinforcement in the lateral portion of the sclerite. [Hipposideros armiger, at base of ear or on lower jaw area, SE China.]....................................................................................A. sanmingense Sun, Ding, Yan, Pape & Zhang, 2021
- 16 First abdominal annular row (R1) present; fourth abdominal annular row (R4) absent. [Coelops frithii, in elbow pit where upper arm meets forearm, SE China.]..................................................................................................................Ascodipteron guoliangi sp. nov.
- – First abdominal annular row (R1) absent; fourth abdominal annular row (R4) present. [Rhinolophus spp., Africa.]....................................................................A. brevior Maa, 1965
- 17 MSS comprised of one seta. R1 and R2 absent (M. schreibersi, Africa).............................................................................................................................................A. theodori Maa, 1965
- – MSS lacking setae. R1–R3 present, with short, triangular spines (M. schreibersi, Africa)..........................................................................................................................................A. minor Theodor, 1973
3.2. Biology
3.3. Molecular Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ant | antenna |
| a.s. | anterior thoracic spiracle |
| cerc | cercus |
| cx1–3 | coxa 1–3 |
| fr | frons |
| g | gena |
| hp | hypopleuron |
| lv | lateral vertex |
| ms | mesopleuron |
| pt | pteropleuron |
| s.w. | stump of wing |
| ster | sternopleuron |
| sp5–7 | spiracles 5–7 |
| t.1–3 | trochanter 1–3 |
| vo | vaginal orifice |
| DSS | dorsal spiracular setae |
| MSS | medial spiracular setae |
| VSS | ventral spiracular setae |
| R1–5 | abdominal setae arranged roughly into annular rows, R1 the proximal and R5 the distal row |
References
- Dick, C.W.; Patterson, B.D. Bat flies-obligate ectoparasites of bats. In Micromammals and Macroparasites; Morand, S., Krasnov, B.R., Poulin, R., Eds.; Springer: Tokyo, Japan, 2006; Chapter 11; pp. 179–194. [Google Scholar]
- Pape, T.; Blagoderov, V.; Mostovski, M.B. Order DIPTERA Linnaeus, 1758. In Animal Biodiversity: An Outline of Higher-Level Classification and Survey of Taxonomic Richness; Zhang, Z.Q., Ed.; Magnolia press: Waco, TX, USA, 2011; Volume 3148, pp. 222–229. [Google Scholar]
- Hastriter, M.W. A review of Ascodipterinae (Diptera: Streblidae) of the Oriental and Australasian regions with a description of three new species of Ascodipteron Adensamer and a key to the subfamily. Zootaxa 2007, 1636, 1–32. [Google Scholar] [CrossRef]
- Sun, H.; Ding, L.; Yan, L.; Pape, T.; Zhang, D. Ascodipteron sanmingensis sp. nov., a new bat fly (Hippoboscidae: Streblid grade) from Fujian, China. Biodivers. Data J. 2021, 9, e64558. [Google Scholar] [CrossRef] [PubMed]
- Hastriter, M.W.; Bush, S.E. Maabella gen. nov. (Streblidae: Ascodipterinae) from Guangxi Province, China and Vietnam with notes on preservation of Ascodipterinae. Zootaxa 2006, 1176, 27–40. [Google Scholar] [CrossRef]
- Muir, F. Two new species of Ascodipteron. Bull. Mus. Comp. Zool. 1912, 54, 351–366. [Google Scholar]
- Jobling, B. On the African Streblidae (Diptera Acalypterae) including the morphology of the genus Ascodipteron Adens. and a description of a new species. Parasitology 1939, 31, 147–165. [Google Scholar] [CrossRef]
- Tang, Z.; Wang, Z.; Zheng, C.; Fang, J. Biodiversity in China’s mountains. Front. Ecol. Environ. 2006, 4, 347–352. [Google Scholar] [CrossRef]
- Cumming, J.; Wood, D.M. Adult morphology and terminology. In Manual of Afrotropical Diptera; Kirk-Spriggs, A.H., Sinclair, B.J., Eds.; Suricata 4, SANBI Publications: Pretoria, South Africa, 2017; Volume 1, Chapter 3; pp. 89–133. [Google Scholar]
- Dittmar, K.; Whiting, M.F. Genetic and phylogeographic structure of populations of Pulex simulans (Siphonaptera) in Peru inferred from two genes (CytB and CoII). Parasitol. Res. 2003, 91, 55–59. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Zhang, D.; Yan, L.; Zhang, M.; Chu, H.; Cao, J.; Li, K.; Hu, D.; Pape, T. Phylogenetic inference of calyptrates, with the first mitogenomes for Gasterophilinae (Diptera: Oestridae) and Paramacronychiinae (Diptera: Sarcophagidae). Int. J. Biol. Sci. 2016, 12, 489–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Pape, T.; Elgar, M.A.; Gao, Y.; Zhang, D. Evolutionary history of stomach bot flies in the light of mitogenomics. Syst. Entomol. 2019, 44, 797–809. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Zhang, M.; Pape, T.; Gu, C.W.; Wu, W. Sarcophaga (Hoa) flexuosa Ho (Diptera: Sarcophagidae): Association of sexes using morphological and molecular approaches, and a redefinition of Hoa Rohdendorf. Zootaxa 2013, 3670, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Vogt, K.; Feeroz, M.M.; Hasan, M.K. A re-discovery of Coelops frithii (Chiroptera, Hipposideridae) from its type locality after one and a half century. Mammalia 2022, 86, 271–275. [Google Scholar] [CrossRef]
- ter Hofstede, H.M.; Fenton, M.B.; Whitaker, J.O., Jr. Host and host-site specificity of bat flies (Diptera: Streblidae and Nycteribiidae) on Neotropical bats (Chiroptera). Can. J. Zool. 2004, 82, 616–626. [Google Scholar] [CrossRef]
- Maa, T.C. Revision of the Australian batflies. Pac. Insects Monogr. 1971, 28, 1–118. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | GenBank Accession Numbers | Host | Locality | ||
|---|---|---|---|---|---|
| Cytb | COI | ||||
| 1 | Ascodipteron guoliangi sp. nov. 1 | - | OP900074 | Coelops frithii | Chenda Town, Sanyuan District, Sanming, Fujian, China (in abandoned bomb shelter) |
| 2 | Ascodipteron guoliangi sp. nov. 2 | OP903228 | OP900075 | Coelops frithii | Chenda Town, Sanyuan District, Sanming, Fujian, China (in abandoned bomb shelter) |
| 3 | Ascodipteron sp2 1 | - | OP900076 | Hipposideros pratti | Jingzhou, Huaihua, Hunan, China (in cave) |
| 4 | Ascodipteron sp2 2 | - | OP900077 | Hipposideros pratti | Jingzhou, Huaihua, Hunan, China (in cave) |
| 5 | Ascodipteron sp2 3 | OP903229 | OP900078 | Hipposideros pratti | Jingzhou, Huaihua, Hunan, China (in cave) |
| 6 | Ascodipteron sp2 4 | OP903230 | OP900079 | Hipposideros pratti | Jingzhou, Huaihua, Hunan, China (in cave) |
| 7 | Ascodipteron sanmingense1 Sun, Ding, Yan, Pape and Zhang, 2021 | OP903231 | OP900080 | Hipposideros armiger | Sanming, Fujian, China (in abandoned bomb shelter) |
| 8 | Ascodipteron sanmingense2 | - | OP900081 | Hipposideros armiger | Tianmushan, Hangzhou, Zhejiang, China (in cave) |
| 9 | Ascodipteron sanmingense3 | OP903232 | OP900082 | hatched from a puparium | Chenda Town, Sanyuan District, Sanming, Fujian, China (in abandoned bomb shelter) |
| 10 | Ascodipteron sanmingense4 | OP903233 | - | hatched from a puparium | Chenda Town, Sanyuan District, Sanming, Fujian, China (in abandoned bomb shelter) |
| 11 | Ascodipteron sanmingense5 | - | OP900083 | Hipposideros armiger | Qunying Second Village, Meilie District, Sanming, Fujian, China (in abandoned bomb shelter) |
| 12 | Ascodipteron sanmingense6 | - | OP900084 | Hipposideros armiger | Qunying Second Village, Meilie District, Sanming, Fujian, China (in abandoned bomb shelter) |
| 13 | Ascodipteron sanmingense * | MW822598.1 | - | Hipposideros armiger | Qunying Second Village, Meilie District, Sanming, Fujian, China (in abandoned bomb shelter) |
| 14 | Ascodipteron speiserianum1 Muir, 1912 | OP903234 | OP900085 | Miniopterus sp. | Xianren cave, Haikou, Hainan, China |
| 15 | Ascodipteron speiserianum2 | OP903235 | OP900086 | Miniopterus sp. | Xianren cave, Haikou, Hainan, China |
| 16 | Ascodipteron phyllorhinae * Adensamer, 1896 | DQ133149.1 | - | Hipposiderus bicolor | Tiger Cave, Penang, Malaysia |
| 17 | Ascodipteron n. sp. * | DQ133154.1 | - | Hipposiderus bicolor | Tiger Cave, Penang, Malaysia |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Ascodipteron guoliangi sp. nov. 2 | ||||||||||
| 2 | Ascodipteron sp2 3 | 0.0490 | |||||||||
| 3 | Ascodipteron sp2 4 | 0.0490 | 0.0000 | ||||||||
| 4 | Ascodipteron phyllorhinae * | 0.0631 | 0.0279 | 0.0279 | |||||||
| 5 | Ascodipteron sanmingense1 | 0.1011 | 0.0831 | 0.0831 | 0.0811 | ||||||
| 6 | Ascodipteron sanmingense3 | 0.0980 | 0.0801 | 0.0801 | 0.0811 | 0.0079 | |||||
| 7 | Ascodipteron sanmingense4 | 0.0980 | 0.0801 | 0.0801 | 0.0780 | 0.0026 | 0.0052 | ||||
| 8 | Ascodipteron sanmingense * | 0.0937 | 0.0751 | 0.0751 | 0.0780 | 0.0027 | 0.0027 | 0.0000 | |||
| 9 | Ascodipteron speiserianum1 | 0.1320 | 0.1164 | 0.1164 | 0.1093 | 0.1192 | 0.1224 | 0.1224 | 0.1223 | ||
| 10 | Ascodipteron speiserianum2 | 0.1320 | 0.1164 | 0.1164 | 0.1093 | 0.1192 | 0.1224 | 0.1224 | 0.1223 | 0.0000 | |
| 11 | Ascodipteron sp. * | 0.1229 | 0.1096 | 0.1096 | 0.1063 | 0.1227 | 0.1160 | 0.1193 | 0.1193 | 0.0914 | 0.0914 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Ascodipteron guoliangi sp. nov. 1 | ||||||||||||
| 2 | Ascodipteron guoliangi sp. nov. 2 | 0.0063 | |||||||||||
| 3 | Ascodipteron sp2 1 | 0.0504 | 0.0523 | ||||||||||
| 4 | Ascodipteron sp2 2 | 0.0494 | 0.0521 | 0.0113 | |||||||||
| 5 | Ascodipteron sp2 3 | 0.0510 | 0.0477 | 0.0084 | 0.0137 | ||||||||
| 6 | Ascodipteron sp2 4 | 0.0510 | 0.0461 | 0.0084 | 0.0120 | 0.0000 | |||||||
| 7 | Ascodipteron sanmingense1 | 0.0636 | 0.0655 | 0.0614 | 0.0655 | 0.0584 | 0.0584 | ||||||
| 8 | Ascodipteron sanmingense2 | 0.0732 | 0.0750 | 0.0629 | 0.0706 | 0.0628 | 0.0628 | 0.0145 | |||||
| 9 | Ascodipteron sanmingense3 | 0.0611 | 0.0611 | 0.0590 | 0.0565 | 0.0495 | 0.0495 | 0.0036 | 0.0107 | ||||
| 10 | Ascodipteron sanmingense5 | 0.0626 | 0.0645 | 0.0533 | 0.0593 | 0.0533 | 0.0533 | 0.0034 | 0.0120 | 0.0000 | |||
| 11 | Ascodipteron sanmingense6 | 0.0664 | 0.0683 | 0.0570 | 0.0635 | 0.0570 | 0.0570 | 0.0000 | 0.0120 | 0.0036 | 0.0034 | ||
| 12 | Ascodipteron speiserianum1 | 0.0849 | 0.0802 | 0.0721 | 0.0887 | 0.0778 | 0.0761 | 0.0876 | 0.0925 | 0.0726 | 0.0765 | 0.0808 | |
| 13 | Ascodipteron speiserianum2 | 0.0872 | 0.0845 | 0.0833 | 0.0851 | 0.0826 | 0.0826 | 0.0847 | 0.0905 | 0.0724 | 0.0754 | 0.0793 | 0.0034 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, H.; Ding, L.; Pape, T.; Zhang, D. A New Species of Ascodipteron (Diptera: Hippoboscidae) from China Based on Morphology and DNA Barcodes. Insects 2022, 13, 1148. https://doi.org/10.3390/insects13121148
Sun H, Ding L, Pape T, Zhang D. A New Species of Ascodipteron (Diptera: Hippoboscidae) from China Based on Morphology and DNA Barcodes. Insects. 2022; 13(12):1148. https://doi.org/10.3390/insects13121148
Chicago/Turabian StyleSun, Haoran, Liang Ding, Thomas Pape, and Dong Zhang. 2022. "A New Species of Ascodipteron (Diptera: Hippoboscidae) from China Based on Morphology and DNA Barcodes" Insects 13, no. 12: 1148. https://doi.org/10.3390/insects13121148