
Effect of Probiotics on Tenebrio molitor Larval Development and Resistance against the Fungal Pathogen Metarhizium brunneum


{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Insects
2.2. Origin and Culture of Probiotics
2.3. Preparation of Probiotic Diet Mix
2.4. Preparation of Metarhizium brunneum Inoculum
2.5. Bioassay Assessments
2.5.1. Assessment of Larval Body Weight and Survivability of Larvae
2.5.2. Pathogen Challenge with Metarhizium brunneum
2.6. Statistical Analysis
3. Results
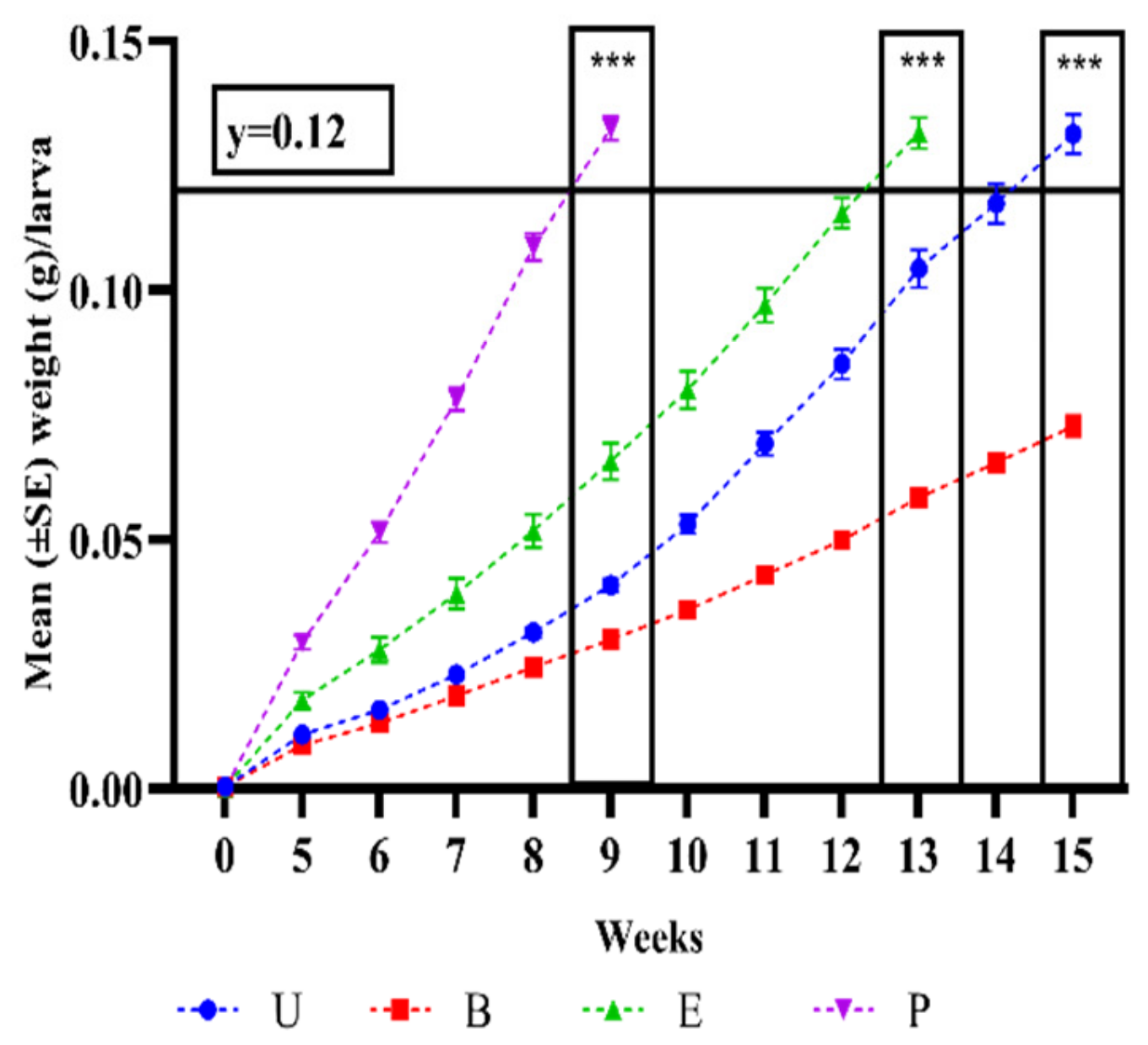
3.1. In Vivo Study of Probiotic-Based Diet Supplementation Effects
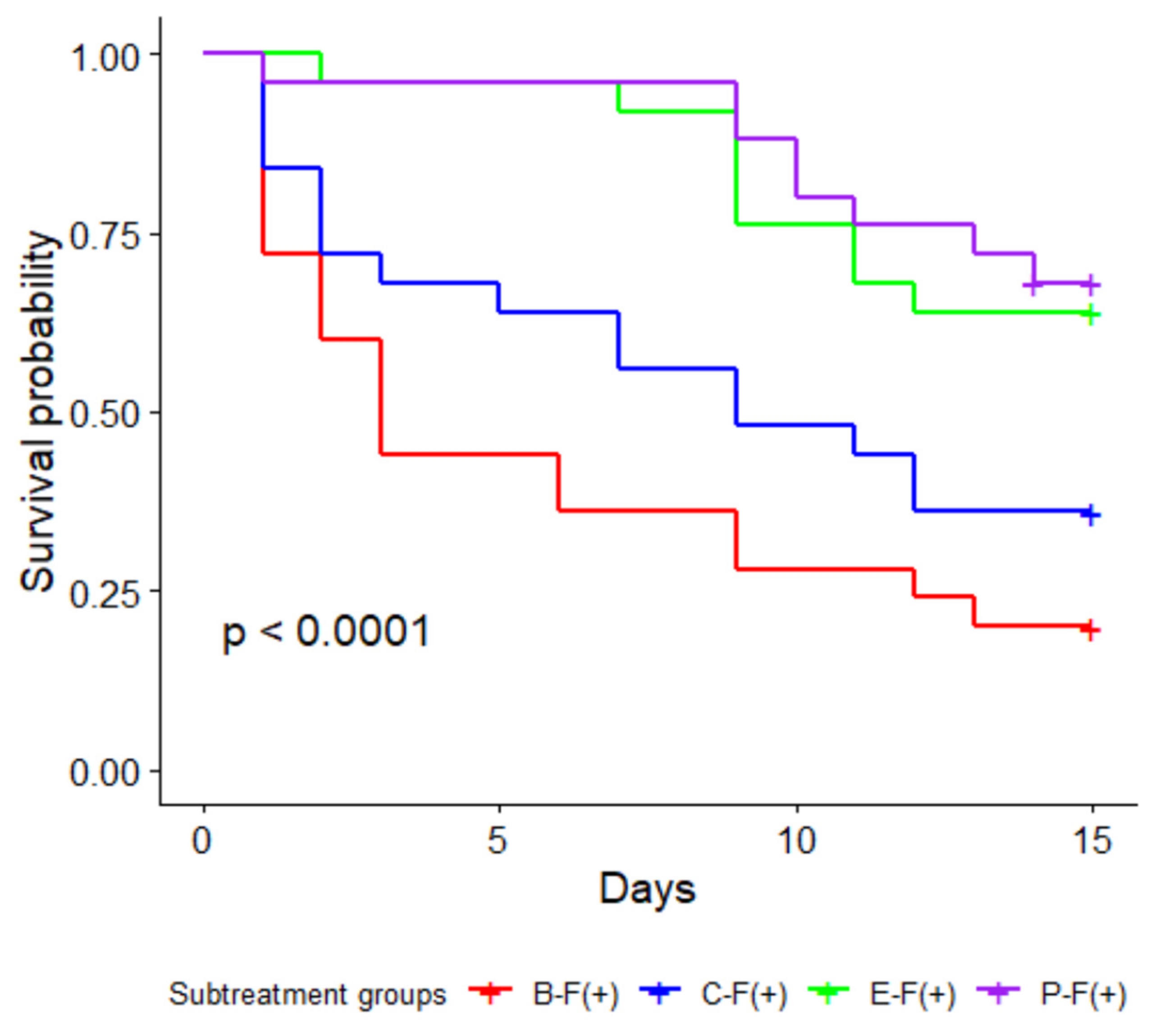
3.2. Pathogen Challenge with the Fungus Metarhizium brunneum
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vassileios, V. Food waste as a potential new source for edible insect mass production for food and feed: A review. Fermentation 2019, 5, 81. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, Y.; Isman, M.B. Insects as an alternative protein source. In Proteins in Food Processing, 2nd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2018; pp. 263–288. [Google Scholar] [CrossRef]
- Carus, M.; Dammer, L. The circular bioeconomy—Concepts, opportunities, and limitations. Ind. Biotechnol. 2018, 14, 83–91. [Google Scholar] [CrossRef]
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed. Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Pippinato, L.; Gasco, L.; Di Vita, G.; Mancuso, T. Current scenario in the European edible-insect industry: A preliminary study. J. Insects Food Feed. 2020, 6, 371–381. [Google Scholar] [CrossRef]
- Rizou, E.; Kalogiouri, N.; Bisba, M.; Papadimittriou, A.; Kyrila, G.; Lazou, A.; Stefanos, A.; Hatzikamari, M.; Mourtzinos, I. Amelioration of growth, nutritional value, and microbial load of Tenebrio molitor (Coleoptera: Tenebrionidae) through probiotic supplemented feed. Eur. Food Res. Technol. 2022, 248, 727–739. [Google Scholar] [CrossRef]
- Grau, T.; Vilcinskas, A.; Joop, G. Sustainable farming of the mealworm Tenebrio molitor for the production of food and feed. Z. Für Nat. 2017, 72, 337–349. [Google Scholar] [CrossRef]
- Lecocq, A.; Natsopoulou, M.E.; Berggreen, I.E.; Eilenberg, J.; Heckmann, L.H.L.; Nielsen, H.V.; Stensvold, C.R.; Jensen, A.B. Probiotic properties of an indigenous Pediococcus pentosaceus strain on Tenebrio molitor larval growth and survival. J. Insects Food Feed. 2021, 7, 975–986. [Google Scholar] [CrossRef]
- Eilenberg, J.; Vlak, J.M.; Nielsen-LeRoux, C.; Cappellozza, S.; Jensen, A.B. Diseases in insects produced for food and feed. J. Insects Food Feed. 2015, 1, 87–102. [Google Scholar] [CrossRef]
- Grau, T.; Vilcinskas, A.; Joop, G. Probiotic Enterococcus munditi isolate protects the model insect Tribolium castaneum against Bacillus thuringiensis. Front. Microbiol. 2017, 8, 1261. [Google Scholar] [CrossRef]
- Castrillo, L.A.; Griggs, M.H.; Ranger, C.M.; Reding, M.E.; Vandenberg, J.D. Virulence of commercial strains of Beauveria bassiana and Metarhizium brunneum (Ascomycota: Hypocreales) against adult Xylosandrus germanus (Coleoptera: Curculionidae) and impact on brood. Biol. Control 2011, 58, 121–126. [Google Scholar] [CrossRef]
- Maciel-Vergara, G.; Jensen, A.B.; Lecocq, A.; Eilenberg, J. Diseases in edible rearing systems. J. Insects Food Feed. 2021, 7, 621–638. [Google Scholar] [CrossRef]
- Savio, C.; Mugo-Kamiri, L.; Upfold, J.K. Bugs in bugs: The role of probiotics and prebiotics in maintenance of health in mass-reared insects. Insects 2022, 13, 376. [Google Scholar] [CrossRef]
- Leger, R.J.S.; Wang, J.B. Metarhizium: Jack of all trades, master of many. Open Biol. 2020, 10, 200307. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. Available online: http://www.nature.com/doifinder/10.1038/nrgastro.2014.66 (accessed on 10 February 2020). [CrossRef] [Green Version]
- Holzapfel, W.H.; Schillinger, U. Introduction to pre- and probiotics. Food Res. Int. 2002, 35, 109–116. [Google Scholar] [CrossRef]
- Kanak, E.K.; Yilmaz, S.Ö. Identification, antibacterial and antifungal effects, antibiotic resistance of some lactic acid bacteria. Food Sci. Technol. 2020, 41, 174–182. [Google Scholar] [CrossRef]
- Anee, I.J.; Alam, S.; Begum, R.A.; Shahjahan, R.M.; Khandaker, A.M. The role of probiotics on animal health and nutrition. J. Basic Appl. Zool. 2021, 82, 52. [Google Scholar] [CrossRef]
- R Studio, Version 4.0.3.; Posit Software: Boston, MA, USA. Available online: https://www.rstudio.com/products/rstudio/download/ (accessed on 5 December 2020).
- Hamden, H.; Guerfali, M.M.S.; Fadhl, S.; Saidi, M.; Chevrier, C. Fitness improvement of mass-reared sterile males of Ceratitis capitata (Vienna 8 strain) (Diptera: Tephritidae) after gut enrichment with probiotics. J. Econ. Entomol. 2013, 106, 641–647. [Google Scholar] [CrossRef]
- Augustinos, A.A.; Kyritsis, G.A.; Papadopoulos, N.T.; Abd-Alla, A.M.M.; Cáceres, C.; Bourtzis, K. Exploitation of the medfly gut microbiota for the enhancement of sterile insect technique: Use of Enterobacter sp. in larval diet-based probiotic applications. PLoS ONE 2015, 10, e0136459. [Google Scholar] [CrossRef] [Green Version]
- Audisio, M.C. Gram-Positive Bacteria with Probiotic Potential for the Apis mellifera L. Honeybee: The experience in the Northwest of Argentina. Probiotics Antimicrob. Proteins 2017, 9, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, Z.M.; Górka, P.; Schlagheck, A.; Jagusiak, W.; Micek, P.; Strzetelski, J. Performance of Holstein calves fed milk-replacer and starter mixture supplemented with probiotic feed additive. J. Anim. Feed Sci. 2009, 18, 399–411. [Google Scholar] [CrossRef]
- Gong, L.; He, H.; Li, D.; Cao, L.; Khan, T.A.; Li, Y.; Pan, L.; Yan, L.; Ding, X.; Sun, Y.; et al. A new isolate of Pediococcus pentosaceus (SL001) with antibacterial activity against fish pathogens and potency in facilitating the immunity and growth performance of grass carps. Front. Microbiol. 2019, 10, 1384. [Google Scholar] [CrossRef]
- Wang, W.; Cai, H.; Zhang, A.; Chen, Z.; Chang, W.; Liu, G.; Deng, X.; Bryden, W.L.; Zheng, A. Enterococcus faecium modulates the gut microbiota of broilers and enhances phosphorus absorption and utilization. Animals 2020, 20, 1232. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Ingale, S.L.; Kim, Y.W.; Kim, J.S.; Kim, K.H.; Lohakare, J.D.; Kim, E.K.; Kim, H.S.; Ryu, M.H.; Kwon, I.K.; et al. Effect of supplementation of Bacillus subtilis LS 1–2 to broiler diets on growth performance, nutrient retention, caecal microbiology and small intestinal morphology. Res. Vet. Sci. 2012, 93, 264–268. [Google Scholar] [CrossRef]
- Hu, Y.; Dun, Y.; Li, S.; Zhao, S.; Peng, N.; Liang, Y. Effects of Bacillus subtilis KN-42 on growth performance, diarrhea, and faecal bacterial flora of weaned piglets. Asian Australas. J. Anim. Sci. 2014, 27, 1131–1140. [Google Scholar] [CrossRef]
- Chai, P.C.; Song, X.L.; Chen, G.F.; Xu, H.; Huang, J. Dietary supplementation of probiotic Bacillus subtilis PC465 isolated from the gut of Fenneropenaeus chinensis improves the health status and resistance of Litopenaeus vannamei against white spot syndrome virus. Fish Shellfish. Immunol. 2016, 54, 602–611. [Google Scholar] [CrossRef]
- Sun, L.; Khalid, A.; Khalid, F.; Wang, Z. Effect of Bacillus Subtilis Protease on growth, production performance and feed efficiency of silkworm, Bombyx mori L. Res. Sq. 2020, 1. [Google Scholar] [CrossRef]
- Douglas, A.E. The microbial dimension in insect nutritional ecology. Funct. Ecol. 2009, 23, 38–47. [Google Scholar] [CrossRef]
- Dicks, L.M.T.; Botes, M. Probiotic lactic acid bacteria in the gastrointestinal tract: Health benefits, safety, and mode of action. Benef. Microbes 2010, 1, 11–29. [Google Scholar] [CrossRef]
- Hao, L.; Cheng, Y.; Su, W.; Wang, C.; Lu, Z.; Jin, M.; Wang, F.; Wang, Y. Pediococcus pentosaceus ZJUAF-4 relieves oxidative stress and restores the gut microbiota in diquat-induced intestinal injury. Appl. Microbiol. Biotechnol. 2021, 105, 1657–1668. [Google Scholar] [CrossRef]
- Huang, J.; Li, S.; Wang, Q.; Guan, X.; Qian, L.; Li, J.; Zheng, Y.; Lin, B. Pediococcus pentosaceus B49 from human colostrum ameliorates constipation in mice. Food Funct. 2020, 11, 5607–5620. [Google Scholar] [CrossRef]
- Samli, H.E.; Senkoylu, N.; Koc, F.; Kanter, M.; Agma, A. Effects of Enterococcus faecium and dried whey on broiler performance, gut histomorphology and intestinal microbiota. Arch. Anim. Nutr. 2007, 61, 42–49. [Google Scholar] [CrossRef]
- Mallo, J.J.; Rioperez, J.; Honrubia, P. The addition of Enterococcus faecium to diet improves piglet’s intestinal microbiota and performance. Livest. Sci. 2010, 133, 176–178. [Google Scholar] [CrossRef] [Green Version]
- Nghe, D.; Nguyen, T. Characterization of antimicrobial activities of Pediococcus pentosaceus Vtcc-B-601. J. Appl. Pharm. Sci. 2014, 4, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Cai, L.; Lv, L.; Li, L. Pediococcus pentosaceus, a future additive or probiotic candidate. Microb. Cell Fact. 2021, 20, 45. [Google Scholar] [CrossRef]
- Corr, S.C.; Hill, C.; Gahan, C.G.M. Understanding the mechanisms by which probiotics inhibit gastrointestinal pathogens. In Advances in Food and Nutrition Research, 1st ed.; Academic Press: Cambridge, MA, USA; Elsevier Ltd.: Amsterdam, The Netherlands, 2009; Volume 56, pp. 1–15. [Google Scholar] [CrossRef]
- Strompfová, V.; Lauková, A.; Ová, D.M. Effect of bacteriocin-like substance produced by Enterococcus faecium EF55 on the composition of avian gastrointestinal microflora. Acta Vet. Brno. 2003, 72, 559–564. Available online: https://actavet.vfu.cz/media/pdf/avb_2003072040559.pdf (accessed on 20 February 2020). [CrossRef] [Green Version]
- Papagianni, M.; Anastasiadou, S. Pediocins: The bacteriocins of Pediococci. Sources, production, properties, and applications. Microb. Cell Factories 2009, 8, 3. [Google Scholar] [CrossRef]
- Wynants, E.; Crauwels, S.; Lievens, B.; Luca, S.; Claes, J.; Borremans, A.; Bruyninckx, L.; Van Campenhout, L. Effect of post-harvest starvation and rinsing on the microbial numbers and the bacterial community composition of mealworm larvae (Tenebrio molitor). Innov. Food Sci. Emerg. Technol. 2017, 42, 8–15. [Google Scholar] [CrossRef]
- Osimani, A.; Milanović, V.; Cardinali, F.; Garofalo, C.; Clementi, F.; Pasquini, M.; Riolo, P.; Ruschioni, S.; Isidoro, N.; Loreto, N.; et al. The bacterial biota of laboratory-reared edible mealworms (Tenebrio molitor L.): From feed to frass. Int. J. Food Microbiol. 2018, 272, 49–60. [Google Scholar] [CrossRef]
- Leyva-Madrigal, K.Y.; Luna-González, A.; Escobedo-Bonilla, C.M.; Fierro-Coronado, J.A.; Maldonado-Mendoza, I.E. Screening for potential probiotic bacteria to reduce prevalence of WSSV and IHHNV in whiteleg shrimp (Litopenaeus vannamei) under experimental conditions. Aquaculture 2011, 322–323, 16–22. [Google Scholar] [CrossRef]
- Naidu, A.S.; Bidlack, W.R.; Clemens, R.A. Probiotic spectra of lactic acid bacteria (LAB). Crit. Rev. Food Sci. Nutr. 1999, 39, 13–126. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.X.; Wang, Y.B.; Li, W.F. Effect of probiotic on larvae shrimp (Penaeus vannamei) based on water quality, survival rate and digestive enzyme activities. Aquaculture 2009, 287, 349–353. [Google Scholar] [CrossRef]
- Zuo, Z.H.; Shang, B.J.; Shao, Y.C.; Li, W.Y.; Sun, J.S. Screening of intestinal probiotics and the effects of feeding probiotics on the growth, immune, digestive enzyme activity, and intestinal flora of Litopenaeus vannamei. Fish Shellfish. Immunol. 2019, 86, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, S. Molecular Characterization of Digestive Proteases of the Yellow Mealworm, Tenebrio molitor L. Ph.D. Thesis, Kansas State University, Manhattan, KS, USA, 2006. [Google Scholar]
- Rodjaroen, S.; Thongprajukaew, K.; Khongmuang, P.; Malawa, S.; Tuntikawinwong, K.; Saekhow, S. Ontogenic development of digestive enzymes in mealworm larvae (Tenebrio molitor L.) and their suitable harvesting time for use as fish feed. Insects 2020, 11, 393. [Google Scholar] [CrossRef]
- Won, S.; Hamidoghli, A.; Choi, W.; Bae, J.; Jang, W.J.; Lee, S.; Bai, S.C. Evaluation of potential probiotics Bacillus subtilis WB60, Pediococcus pentosaceus, and Lactococcus lactis on growth performance, immune response, gut histology, and immune-related genes in whiteleg shrimp, Litopenaeus vannamei. Microorganisms 2020, 8, 281. [Google Scholar] [CrossRef] [Green Version]
- Ringø, E.; Van Doan, H.; Lee, S.; Song, S.K. Lactic Acid Bacteria in shellfish: Possibilities and challenges. Rev. Fish. Sci. Aquac. 2020, 28, 139–169. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Sadegh, T.H.; Dawood, M.A.O.; Dadar, M.; Sheikhzadeh, N. The effects of dietary Pediococcus pentosaceus on growth performance, hemato-immunological parameters, and digestive enzyme activities of common carp (Cyprinus carpio). Aquaculture 2020, 516, 734656. [Google Scholar] [CrossRef]
- Ziaei-Nejad, S.; Rezaei, M.H.; Takami, G.A.; Lovett, D.L.; Mirvaghefi, A.R.; Shakouri, M. The effect of Bacillus spp. bacteria used as probiotics on digestive enzyme activity, survival, and growth in the Indian white shrimp Fenneropenaeus indicus L. Aquaculture 2006, 252, 516–524. [Google Scholar] [CrossRef]
- Zheng, X.; Duan, Y.; Dong, H.; Zhang, J. Effects of Dietary Lactobacillus plantarum on growth performance, digestive enzymes, and gut morphology of Litopenaeus vannamei. Probiotics Antimicrob. Proteins 2018, 10, 504–510. [Google Scholar] [CrossRef]
- Xing, C.F.; Hu, H.H.; Huang, J.B.; Fang, H.C.; Kai, Y.H.; Wu, Y.C.; Chi, S.C. Diet supplementation of Pediococcus pentosaceus in cobia (Rachycentron canadum) enhances growth rate, respiratory burst, and resistance against photobacteriosis. Fish Shellfish Immunol. 2013, 35, 1122–1128. [Google Scholar] [CrossRef]
- Juega, M.; Costantini, A.; Bonello, F.; Cravero, M.C.; Martinez-Rodriguez, A.J.; Carrascosa, A.V.; Garcia-Moruno, E. Effect of malolactic fermentation by Pediococcus damnosus on the composition and sensory profile of Albariño and Caiño white wines. J. Appl. Microbiol. 2014, 116, 586–595. [Google Scholar] [CrossRef]
- El-Haroun, E.R.; Goda, A.M.A.S.; Chowdhury, K.M.A. Effect of dietary probiotic Biogen® supplementation as a growth promoter on growth performance and feed utilization of Nile tilapia Oreochromis niloticus (L.). Aquac. Res. 2006, 37, 1473–1480. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, J.; Zhong, H.; Li, N.; Xu, H.; Zhu, Q.; Liu, Y. Effect of probiotics on the meat flavour and gut microbiota of chicken. Sci. Rep. 2017, 7, 6400. [Google Scholar] [CrossRef] [Green Version]
- Gheisar, M.; Hosseindoust, A.; Kim, I.H. Effects of dietary Enterococcus faecium on growth performance, carcass characteristics, faecal microbiota, and blood profile in broilers. Vet. Med. 2016, 61, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Bliūdžiūtė, S. Effects of Mannan-Oligosaccharides and Probiotics Pediococcus pentosaceus ir Lactobacillus sakei on Growth Performance, Microvilli Length of Gut and Morphological Blood Parameters of Carp (Cyprinus carpio) Farming in a Closed Recirculating System. Master’s Thesis, Lithuanian University of Health Sciences, Kaunas, Lithuania, 2018. Available online: https://hdl.handle.net/20.500.12512/103735 (accessed on 25 January 2020).
- Keebaugh, E.S.; Yamada, R.; Obadia, B.; Ludington, W.B.; Ja, W.W. Microbial quantity impacts drosophila nutrition, development, and lifespan. iScience 2018, 4, 247–259. [Google Scholar] [CrossRef]
- Kataria, J.; Li, N.; Wynn, J.L.; Neu, J. Probiotic microbes: Do they need to be alive to be beneficial? Nutr. Rev. 2009, 67, 546–550. [Google Scholar] [CrossRef]
- Cawoy, H.; Bettiol, W.; Fickers, P.; Ongena, M. Chapter 13. Bacillus-based biological control of plant diseases. In Pesticides in the Modern World—Pesticides Use and Management; Embrapa Meio Ambiente: Jaguariúna, Brazil, 2011; Volume 13, pp. 273–302. Available online: https://www.alice.cnptia.embrapa.br/bitstream/doc/920753/1/2011CL08.pdf (accessed on 17 March 2020).
- Liu, Q.; Zhang, B.; Shen, Y.; Yin, K. Effect of the protein elicitor AMEP412 from Bacillus subtilis artificially fed to adults of the whitefly, Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae). Egypt. J. Biol. Pest Control 2020, 30, 3. [Google Scholar] [CrossRef]
- Valenzuela-Soto, J.H.; Estrada-Hernández, M.G.; Ibarra-Laclette, E.; Délano-Frier, J.P. Inoculation of tomato plants (Solanum lycopersicum) with growth-promoting Bacillus subtilis retards whitefly Bemisia tabaci development. Planta 2010, 231, 397–410. [Google Scholar] [CrossRef]
- Ghribi, D.; Elleuch, M.; Abdelkefi-Mesrati, L.; Boukadi, H.; Ellouze-Chaabouni, S. Histopathological effects of Bacillus subtilis SPB1 biosurfactant in the midgut of Ephestia kuehniella (Lepidoptera: Pyralidae) and improvement of its insecticidal efficiency. J. Plant Dis. Prot. 2012, 119, 24–29. [Google Scholar] [CrossRef]
- Das, K.; Mukherjee, A.K. Assessment of mosquito larvicidal potency of cyclic lipopeptides produced by Bacillus subtilis strains. Acta Trop. 2006, 97, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, R.; Revathi, K.; Nisha, S.; Kirubakaran, S.A.; Sathish-Narayanan, S.; Senthil-Nathan, S. Physiological effect of chitinase purified from Bacillus subtilis against the tobacco cutworm Spodoptera litura Fab. Pestic. Biochem. Physiol. 2012, 104, 65–71. [Google Scholar] [CrossRef]
- Bulet, P.; Cociancich, S.; Dimarcq, J.L.; Lambert, J.; Reichhart, J.M.; Hoffmann, D.; Hetru, C.; Hoffmann, J.A. Insect immunity: Isolation from a coleopteran insect of a novel inducible antibacterial peptide and of new members of the insect defensin family. J. Biol. Chem. 1991, 266, 24520–24525. [Google Scholar] [CrossRef] [PubMed]
- Miyanoshita, A.; Hara, S.; Sugiyama, M.; Asaoka, A.; Taniai, K.; Yukuhiro, F.; Yamakawa, M. Isolation and characterization of a new member of the insect defensin family from a beetle, Allomyrina dichotoma. Biochem. Biophys. Res. Commun. 1996, 220, 526–531. [Google Scholar] [CrossRef]
- Zokaeifar, H.; Balcázar, J.L.; Saad, C.R.; Kamarudin, M.S.; Sijam, K.; Arshad, A.; Nejat, N. Effects of Bacillus subtilis on the growth performance, digestive enzymes, immune gene expression and disease resistance of white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2012, 33, 683–689. [Google Scholar] [CrossRef] [Green Version]
- Olmos, J.; Acosta, M.; Mendoza, G.; Pitones, V. Bacillus subtilis, an ideal probiotic bacterium to shrimp and fish aquaculture that increases feed digestibility, prevent microbial diseases, and avoid water pollution. Arch. Microbiol. 2020, 202, 427–435. [Google Scholar] [CrossRef]
- Hansen, C. Strains Matter: Selected Combinations Is the Key to a Successful Solution. pp. 1–4. Available online: http://www.chr.hansen.com/en/animal-health/cards/article-cards/strains-matter (accessed on 25 March 2021).
- Ilavenil, S.; Vijayakumar, M.; Kim, D.H.; Arasu, M.V.; Park, H.S.; Ravikumar, S.; Choi, K.C. Assessment of probiotic, antifungal and cholesterol lowering properties of Pediococcus pentosaceus KCC-23 isolated from Italian ryegrass. J. Sci. Food Agric. 2016, 96, 593–601. [Google Scholar] [CrossRef]
- Sadeghi, A.; Raeisi, M.; Ebrahimi, M.; Sadeghi, B. Antifungal activity of Pediococcus pentosaceus isolated from whole barley sourdough. J. Food Qual. Hazards Control 2016, 3, 30–36. Available online: https://jfqhc.ssu.ac.ir/article-1-228-en.pdf (accessed on 15 January 2020).
- González-santoyo, I.; Córdoba-aguilar, A. Phenoloxidase: A key component of the insect immune system. Entomol. Exp. Et Appl. 2012, 142, 1–16. [Google Scholar] [CrossRef]
- Zhang, L.; Chaomin, S. Fengycins, cyclic lipopeptides from marine Bacillus subtilis strains, kill the plant-pathogenic fungus Magnaporthe grisea by inducing reactive oxygen species production and chromatin condensation. Appl. Environ. Microbiol. 2018, 84, e00445-18. [Google Scholar] [CrossRef] [Green Version]
- Mardanova, A.M.; Hadieva, G.F.; Lutfullin, M.T.; Khilyas, I.V.; Minnullina, L.F.; Gilyazeva, A.G.; Bogomolnaya, L.M.; Sharipova, M.R. Bacillus subtilis strains with antifungal activity against the phytopathogenic fungi. Agric. Sci. 2017, 8, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Saini, P.; Shrivastave, J. Production of peptide antifungal antibiotic and biocontrol activity of Bacillus subtilis. Indian J. Exp. Biol. 2009, 47, 57–62. Available online: http://nopr.niscpr.res.in/bitstream/123456789/2895/1/IJEB%2047%281%29%2057-62.pdf (accessed on 5 February 2020).
- Liu, Y.; Chen, Z.; Ng, T.B.; Zhang, J.; Zhou, M.; Song, F.; Lu, F.; Liu, Y. Bacisubin, an antifungal protein with ribonuclease and hemagglutinating activities from Bacillus subtilis strain B-916. Peptides 2007, 28, 553–559. [Google Scholar] [CrossRef]
- De Roode, J.C.; Lefèvre, T. Behavioral immunity in insects. Insects 2012, 3, 789–820. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zhao, X.Y.; Tang, Q.B.; Chao-Liang Lei, C.L.; Huang, Q.Y. The mechanisms of social immunity against fungal. infections in eusocial insects. Toxins 2019, 11, 244. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dahal, S.; Jensen, A.B.; Lecocq, A. Effect of Probiotics on Tenebrio molitor Larval Development and Resistance against the Fungal Pathogen Metarhizium brunneum. Insects 2022, 13, 1114. https://doi.org/10.3390/insects13121114
Dahal S, Jensen AB, Lecocq A. Effect of Probiotics on Tenebrio molitor Larval Development and Resistance against the Fungal Pathogen Metarhizium brunneum. Insects. 2022; 13(12):1114. https://doi.org/10.3390/insects13121114
Chicago/Turabian StyleDahal, Sabina, Annette Bruun Jensen, and Antoine Lecocq. 2022. "Effect of Probiotics on Tenebrio molitor Larval Development and Resistance against the Fungal Pathogen Metarhizium brunneum" Insects 13, no. 12: 1114. https://doi.org/10.3390/insects13121114