



Floral Volatile Organic Compounds and a List of Pollinators of Fallopia baldschuanica (Polygonaceae)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
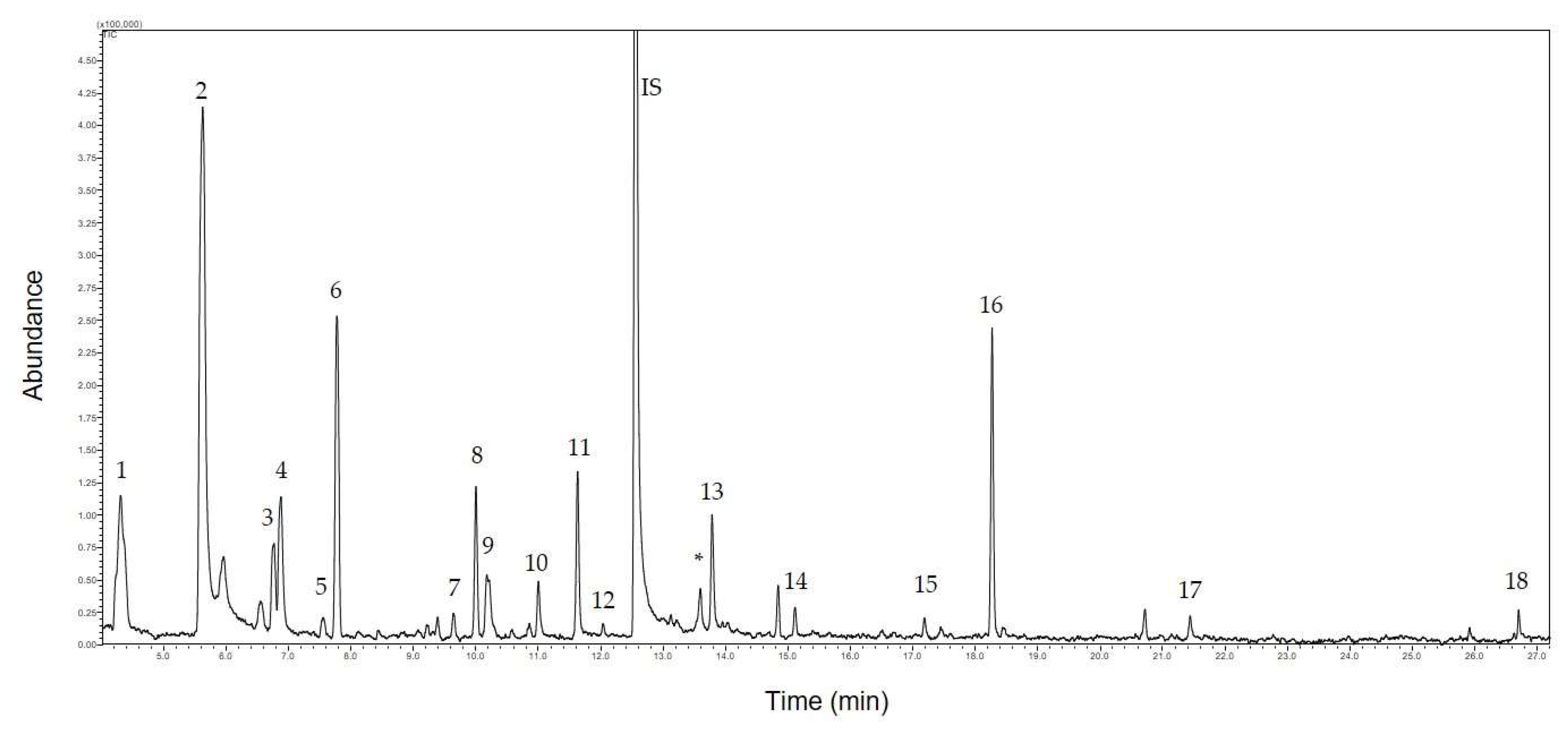
2.2. HS-GC/S Analysis of Volatiles Fractions
2.3. Field Observations of Insects Activity
3. Results
3.1. Identification of VOCs
3.2. True Pollinators and Visitors Insects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WFO. 2022: World Flora Online. Fallopia baldschuanica (Regel) Holub. Available online: http://www.worldfloraonline.org/taxon/wfo-0000685779 (accessed on 3 August 2022).
- Kubát, K.; Hrouda, L.; Chrtek, J.; Kaplan, Z.; Kirschner, J.; Stěpánek, J. Klíč ke květeně České republiky [Field Guide to the Flowers of the Czech Republic]; Academia: Praha, Czech Republic, 2002. [Google Scholar]
- Hassler, M. World Plants: Synonymic Checklists of the Vascular Plants of the World (version Nov 2018). In Species 2000 and ITIS Catalogue of Life; (2019) Annual Checklist; Roskov, Y., Ower, G., Orrell, T., Nicolson, D., Bailly, N., Kirk, P.M., Bourgoin, T., DeWalt, R.E., Decock, W., van Nieukerken, E., et al., Eds.; Species 2000: Naturalis, Leiden, 2020; Available online: www.catalogueoflife.org/annual-checklist/2019 (accessed on 2 October 2022).
- Stace, C.A.; Thompson, H. New Flora of the British Isles; University Press: Cambridge, UK, 2010. [Google Scholar]
- Gutte, P.; Hardtke, H.J.; Schmidt, P.A. (Eds.) Die Flora Sachsens und der angrenzenden Gebiete: Ein pflanzenkundlicher Exkursionsführer; Quelle & Meyer: Wiebelsheim, Germany, 2013. [Google Scholar]
- World Checklist of Selected Plant Families (WCSP). Available online: http://wcsp.science.kew.org/prepareChecklist.do;jsessionid=BD13BC5465862F5D74079A6ACC47EE0B.kppapp06-wcsp?checklist=selected_families%40%40135140520202307081 (accessed on 15 July 2020).
- Li, A.J.; Bao, B.; Grabovskaya-Borodina, A.E.; Hong, S.P.; McNeill, J.; Mosyakin, S.L.; Ohba, H.; Park, C.W. Polygonaceae. In Flora of China 5; Wu, Z.Y., Raven, P.H., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MI, USA, 2003; pp. 277–350. [Google Scholar]
- Rutkowski, L. Klucz do oznaczania roślin naczyniowych Polski niżowej; Wyd. Naukowe PWN: Warszawa, Poland, 2006. [Google Scholar]
- USDA; ARS. National Genetic Resources Program; Germplasm Resources Information Network-(GRIN): Beltsville, MD, USA, 2009; [Online Database]. [Google Scholar]
- Arnold, K. Russian Vine–Fallopia baldschuanica. 2020. Available online: https://www.graftingardeners.co.uk/russian-vine-fallopia-baldschuanica/ (accessed on 20 July 2020).
- Olaru, O.T.; Anghel, A.I.; Istudor, V.; Ancuceanu, R.V.; Dinu, M. Contributions to the pharmacognostical and phytobiological study of Fallopia aubertii (L. Henry) Holub. (Polygonaceae). Farmacia 2013, 61, 991–999. [Google Scholar]
- Wang, S.C.; Tseng, T.Y.; Huang, C.M.; Tsai, T.H. Gardenia herbal active constituents: Applicable separation procedures. J. Chromatogr. B 2004, 812, 193–202. [Google Scholar] [CrossRef]
- Wang, J.; Ma, G.; Hu, J. Phytochemical and chemotaxonomic study on Fallopia aubertii (L. Henry) Holub. Biochem Syst Ecol 2019, 85, 50–53. [Google Scholar] [CrossRef]
- Stace, C.A. New Flora of the British Isles, 2nd ed.; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Hollingsworth, M.L.; Bailey, J.P. Hybridisation and clonal diversity in some introduced Fallopia species (Polygonaceae). Watsonia 2000, 23, 111–122. [Google Scholar]
- Verloove, F. Fallopia baldschuanica. In Manual of the Alien Plants of Belgium; Botanic Garden Meise: Meise, Belgium, 2020; Available online: http://alienplantsbelgium.be/content/fallopia-baldschuanica (accessed on 22 July 2020).
- Stace, C.A.; Crawley, M.J. Alien Plants; Harper Collins: London, UK, 2015. [Google Scholar]
- Booy, O.; Wade, M.; Roy, H. Field Guide to Invasive Plants and Animals in Britain; Bloomsbury Publishing: London, UK, 2015. [Google Scholar]
- Tiébré, M.S.; Vanderhoeven, S.; Saad, L.; Mahy, G. Hybridization and sexual reproduction in the invasive alien Fallopia (Polygonaceae) complex in Belgium. Ann. Bot. 2007, 99, 193–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saad, L.; Tiébré, M.S.; Hardy, O.J.; Mahy, G.; Vanderhoeven, S. Patterns of hybridization and hybrid survival in the invasive alien Fallopia complex (Polygonaceae). Plant Ecol. Evol. 2011, 144, 12–18. [Google Scholar] [CrossRef]
- Johnson, L.R.; Breger, B.; Drummond, F. Novel plant–insect interactions in an urban environment: Enemies, protectors, and pollinators of invasive knotweeds. Ecosphere 2019, 10, e02885. [Google Scholar] [CrossRef]
- Suni, S.; Hall, E.; Bahu, E.; Hayes, H. Urbanization increases floral specialization of pollinators. Ecol. Evol. 2022, 12, e8619. [Google Scholar] [CrossRef]
- El-Sayed, A.M. The Pherobase: Database of Pheromones and Semiochemicals. 2019. Available online: https://www.pherobase.com (accessed on 2 October 2022).
- Berger, W.H.; Parker, F.L. Diversity of Planktonic Foraminifera in Deep Sea Sediments. Science 1970, 168, 1345–1347. [Google Scholar] [CrossRef]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Margalef, R. Information Theory in Ecology. Gen. Syst. 1958, 3, 36–71. [Google Scholar]
- Trojan, P. General Ecology (Ekologia ogólna); PWN Publishing: Warsaw, Poland, 1980. [Google Scholar]
- Jaccard, P. The Distribution of the Flora of the Alpine Zone. New Phytol. 1912, 11, 37–50. [Google Scholar] [CrossRef]
- Huelin, F.E.; Murray, K.E. α-Farnesene in the Natural Coating of Apples. Nature 1966, 210, 1260–1261. [Google Scholar] [CrossRef]
- Nabeta, K.; Ara, Y.; Aoki, Y.; Miyake, M. Biosynthesis of monoterpenes and sesquiterpenes in Larix leptolepis callus from deuterated mevalonates. J. Nat. Prod. 1990, 53, 1241–1248. [Google Scholar] [CrossRef]
- Mostafavi, R.; Henning, J.A.; Gardea-Torresday, J.; Ray, I.M. Variation in aphid alarm pheromone content among glandular and eglandular-haired Medicago accessions. J. Chem. Ecol. 1996, 22, 1629–1638. [Google Scholar] [CrossRef]
- Raal, A.; Kaur, H.; Orav, A.; Arak, E.; Kailas, T.; Müürisepp, M. Content and composition of essential oils in some Asteraceae species. Proc. Est. Acad. Sci. 2011, 60, 55–63. [Google Scholar] [CrossRef]
- Zaks, A.; Davidovich-Rikanati, R.; Bar, E.; Inbar, M.; Lewinsohn, E. Biosynthesis of linalyl acetate and other terpenes in lemon mint (Mentha aquatica var. citrata, Lamiaceae) glandular trichomes. Isr. J. Plant Sci. 2008, 56, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Hillig, K.W. A chemotaxonomic analysis of terpenoid variation in Cannabis. Biochem. Syst. Ecol. 2004, 32, 875–891. [Google Scholar] [CrossRef]
- Avé, D.A.; Gregory, P.; Tingey, W.M. Aphid repellent sesquiterpenes in glandular trichomes of Solanum berthaultii and S. tuberosum. Entomol. Exp. Appl. 1987, 44, 131–138. [Google Scholar] [CrossRef]
- Reinhold, C. Der Einfluss von (E)-β-Farnesen auf das Wirtswahlverhalten von Myzus persicae. 2011. Available online: http://www.clib-jena.mpg.de/theses/ice/ICE11019.pdf (accessed on 13 November 2020).
- Unsicker, S.B.; Kunert, G.; Gershenzon, J. Protective perfumes: The role of vegetative volatiles in plant defense against herbivores. Curr. Opin Plant Biol. 2009, 12, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Novotny, M.; Harvey, S.; Jemiolo, B. Chemistry of male-dominance in the house mouse Mus domesticus. Experientia 1990, 46, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, T.E.; Eggert, M.S.; House, S.J.; Weddell, M.E.; Schulte, B.A.; Rasmussen, L.E.L. Insect pheromones and precursors in female African elephant urine. J. Chem. Ecol. 2006, 32, 1849–1853. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.; Duffield, R.M.; Wheeler, J.W.; Laberge, W.E. Chemistry of the Dufours gland secretions of North American andrenid bees (Hymnoptera, Andrenidae). J. Chem. Ecol. 1981, 7, 453–463. [Google Scholar] [CrossRef]
- Jackson, B.D.; Morgan, E.D.; Billen, J.P.J. Contents of the Pygidial gland of the ant Myrmecia nigriceps. Naturwissenschaften 1990, 77, 187–188. [Google Scholar] [CrossRef]
- Kielty, J.P.; Allen Williams, L.J.; Underwood, N.; Eastwood, E.A. Behavioral responses of three species of ground beetle (Coleoptera, Carabidae) to olfactory cues associated with prey and habitat. J. Insect Behav. 1996, 9, 237–250. [Google Scholar] [CrossRef]
- Šobotník, J.; Hanus, R.; Kalinová, B.; Piskorski, R.; Cvačka, J.; Bourguignon, T.; Roisin, Y. (E,E)-α-Farnesene, an Alarm Pheromone of the Termite Prorhinotermes canalifrons. J. Chem. Ecol. 2008, 34, 478–486. [Google Scholar] [CrossRef]
- Hern, A.; Dorn, S. Sexual dimorphism in the olfactory orientation of adult Cydia pomonella in response to alpha-farnesene. Entomol. Exp. Appl. 1999, 92, 63–72. [Google Scholar] [CrossRef]
- Gerlach, G.; Schill, R. Composition of orchid scents attracting euglossine bees. Bot. Acta 1991, 104, 379–384. [Google Scholar] [CrossRef]
- Gerlach, G.; Schill, R. Die Gattung Coryanthes Hook. (Orchidaceae)—Eine monographische Bearbeitung unter besonderer Berücksichtigung der Blütenduftstoffe. Trop. und subtrop. Pflanzenwelt 1993, 83, 1–205. [Google Scholar]
- Francke, W.; Schulz, S. Pheromones. In Comprehensive Natural Products Chemistry; Barton, S.D., Nakanishi, K., Eds.; Elsevier: Amsterdam, The Netherlands, 1999; Volume 8, pp. 197–261. [Google Scholar] [CrossRef]
- Gibson, R.W.; Pickett, J.A. Wild potato repels aphids by release of aphid alarm pheromone. Nature 1983, 302, 608–609. [Google Scholar] [CrossRef]
- Leroy, P.D.; Verheggen, F.J.; Capella, Q.; Francis, F.; Haubruge, E. An introduction device for the aphidophagous hoverfly Episyrphus balteatus (De Geer) (Diptera: Syrphidae). Biol. Control. 2010, 54, 181–188. [Google Scholar] [CrossRef]
- Krzyżanowski, R. Wpływ lotnych związków na zachowanie mszyc związane z żerowaniem. Kosmos 2017, 66, 413–420. [Google Scholar]
- Giatropoulos, A.; Kimbaris, A.; Michaelakis, A.; Papachristos, D.P.; Polissiou, M.G.; Emmanouel, N. Chemical composition and assessment of larvicidal and repellent capacity of 14 Lamiaceae essential oils against Aedes albopictus. Parasitol. Res. 2018, 117, 1953–1964. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, A.B.; Barszcz, E.S. Hexanal as an attractant for the carrot rust fly Psila rosae (Diptera: Psilidae) for monitoring adults in Ontario. Proc. Entomol. Soc. Ontario. 1997, 128, 85–91. [Google Scholar]
- Mohammed, K.; Agarwal, M.; Li, B.; Newman, J.; Liu, T.; Ren, Y. Evaluation of d-Limonene and β-Ocimene as Attractants of Aphytis melinus (Hymenoptera: Aphelinidae), a Parasitoid of Aonidiella aurantii (Hemiptera: Diaspididae) on Citrus spp. Insects 2020, 11, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farré-Armengol, G.; Filella, I.; Llusià, J.; Peñuelas, J. β-Ocimene, a Key Floral and Foliar Volatile Involved in Multiple Interactions between Plants and Other Organisms. Molecules 2017, 22, 1148. [Google Scholar] [CrossRef] [Green Version]
- Pecetti, L.; Tava, A.; Felicioli, A.; Pinzauti, M.; Piano, E. Effect of three volatile compounds from Lucerne flowers on their attractiveness towards pollinators. Bull. Insectol. 2002, 55, 21–27. [Google Scholar]
- Granero, A.M.; Guerra Sanz, J.M.; Egea Gonzalez, F.J.; Martinez Vidal, J.L.; Dornhaus, A.; Ghani, J.; Serrano, A.R.; Chittka, L. Chemical compounds of the foraging recruitment pheromone in bumblebees. Naturwissenschaften 2005, 92, 371–374. [Google Scholar] [CrossRef] [Green Version]
- Rostelie, T.; Borg-Karlson, A.K.; Mustaparta, H. Selective receptor neurone responses to E-β-ocimene,β-myrcene, E,E-α-farnesene and homo-farnesene in the moth Heliothis virescens, identified by gas chromatography linked to electrophysiology. J. Comp. Physiol. 2000, 186, 833–847. [Google Scholar]
- Kariyat, R.R.; Mauck, K.E.; Balogh, C.M.; Stephenson, A.G.; Mescher, M.C.; De Moraes, C.M. Inbreeding in horsenettle (Solanum carolinense) alters night-time volatile emissions that guide oviposition by Manduca sexta moths. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, T.; Yue, Y.; Shi, M.; Chen, M.; Yang, X.; Wang, L. Exploration of Floral Volatile Organic Compounds in Six Typical Lycoris taxa by GC-MS. Plants 2019, 8, 422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, D.G. Further field evaluation of synthetic herbivore-induced plant volatiles as attractants for beneficial insects. J. Chem. Ecol. 2005, 31, 481–495. [Google Scholar] [CrossRef] [PubMed]
- De Greet, P.; Poland, T.M. Attraction of Hylastes opacus (Coleoptera: Scolytidae) to nonanal. Can. Entomol. 2003, 135, 309–311. [Google Scholar] [CrossRef]
- Metcalf, R.L.; Kogan, M. Plant volatiles as insect attractants. Crit. Rev. Plant Sci. 1987, 5, 251–301. [Google Scholar] [CrossRef]
- Bruce, T.J.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef]
- McCormick, A.C.; Unsicker, S.B.; Gershenzon, J. The specificity of herbivore-induced plant volatiles in attracting herbivore enemies. Trends Plant Sci. 2012, 17, 303–310. [Google Scholar] [CrossRef]
- Kovács-Hostyánszki, A.; Szigeti, V.; Miholcsa, Z.; Sándor, D.; Soltész, Z.; Török, E.; Fenesi, A. Threats and benefits of invasive alien plant species on pollinators, Basic Appl. Ecol. 2022. [Google Scholar] [CrossRef]
- Wenzel, A.; Grass, I.; Belavadi, V.V.; Tscharntke, T. How urbanization is driving pollinator diversity and pollination–A systematic review. Biol. Conserv. 2020, 241, 108321. [Google Scholar] [CrossRef]
- Scheper, J.; Reemer, M.; van Kats, R.; Ozinga, W.A.; van der Linden, G.T.; Schaminée, J.H.; Siepel, H.; Kleijn, D. Museum specimens reveal loss of pollen host plants as key factor driving wild bee decline in The Netherlands. Proc. Natl. Acad. Sci. USA 2014, 111, 17552–17557. [Google Scholar] [CrossRef] [Green Version]
- Cadenasso, M.L.; Pickett, S.T.A.; Grove, J.M. Dimensions of ecosystem complexity: Heterogeneity, connectivity, and history. Ecol. Complex. 2006, 3, 1–13. [Google Scholar] [CrossRef]
- Balogh, L. Japanese, giant and bohemian knotweed. In The Most Important Invasive Plants in Hungary; Botta-Dukát, Z., Balogh, L., Eds.; Institute of Ecology and Botany, Hungarian Academy of Sciences: Vácrátót, Hungary, 2008; pp. 13–33. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Chemical Name | RT | µg/2 g Sample | SD | NIST Search [%] | Kovats Index | Structure | |
|---|---|---|---|---|---|---|---|
| 1 | hexanal | 4.31 | 0.66 | 0.08 | 90 | 824 |  |
| Odor characteristic: green, fruity, acorn, tallowy, fishy, grassy, herbal, leafy | |||||||
| 2 | hex-3-en-1-ol | 5.67 | 7.44 | 0.56 | 95 | 857 |  |
| Odor characteristic: fresh, green, grassy, leafy | |||||||
| 3 | nonane | 6.75 | 1.02 | 0.12 | 94 | 900 |  |
| Odor characteristic: fusel-like | |||||||
| 4 | heptanal | 6.87 | 1.66 | 0.19 | 94 | 904 |  |
| Odor characteristic: citrus, green, fatty, dry fish, pesticide, solvent, smoky, rancid, fruity | |||||||
| 5 | 3-thujene (5-isopropyl-2-methylbicyclo[3.1.0]hex-2-ene) | 7.55 | 0.24 | 0.04 | 92 | 928 |  |
| Odor characteristic: woody, herbal, green | |||||||
| 6 | α-pinene (2,6,6-trimethylbicyclo[3.1.1]hept-2-ene) | 7.75 | 2.48 | 0.19 | 94 | 935 |  |
| Odor characteristic: terpeny, fruity, sweet, green, woody, pine, citrus, lime, camphor | |||||||
| 7 | β -myrcene (7-methyl-3-methylene-1,6-octadiene) | 9.65 | 0.16 | 0.03 | 92 | 991 |  |
| Odor characteristic: metallic, musty, geranium, sweet, fruity, ethereal, soapy, lemon, spicy, woody | |||||||
| 8 | decane | 10 | 1.1 | 0.11 | 95 | 1000 |  |
| Odor characteristic: fusel-like, fruity, sweet | |||||||
| 9 | hex-3-ene-1-ol acetate | 10.21 | 1.34 | 0.18 | 90 | 1008 |  |
| Odor characteristic: green, freshly cut grass, slightly fruity | |||||||
| 10 | limonene (1-methyl-4-(1-methylethenyl)-cyclohexene) | 11.01 | 2.12 | 0.33 | 93 | 1036 |  |
| Odor characteristic: licorice, green, citrus, ethereal, fruity | |||||||
| 11 | β-ocimene ((E)-3,7-dimethyl-1,3,6-octatriene) | 11.63 | 4.02 | 0.76 | 96 | 1056 |  |
| Odor characteristic: herbal, mild, citrus, floral, woody, sweet, orange, lemon | |||||||
| 12 | γ-terpinene (1-isopropyl-4-methyl-cyclohexa-1,4-diene) | 12.05 | 0.06 | 0.01 | 90 | 1069 |  |
| Odor characteristic: citrus, terpeny, herbal, fruity, sweet | |||||||
| 13 | nonanal | 13.78 | 0.72 | 0.1 | 97 | 1118 |  |
| Odor characteristic: gravy, green, tallowy, fruity, gas, chlorine, floral (rose, orris), waxy, sweet, melon, soapy, fatty, lavender, citrus fruit | |||||||
| 14 | hex-3-ene-1-ol butanoate | 15.12 | 0.52 | 0.1 | 97 | 1153 |  |
| Odor characteristic: sweet green, freshly cut grass, slightly fruity | |||||||
| 15 | dodecane | 17.18 | 0.16 | 0.03 | 93 | 1200 |  |
| Odor characteristic: fusel-like | |||||||
| 16 | butanoic acid, 2-methyl, 3-hexenyl ester (3-hexen-1-yl 2-methyl butyrate) | 18.19 | 16 | 2.05 | 96 | 1233 |  |
| Odor characteristic: fresh green apple sweet fruity pear | |||||||
| 17 | 2-butenoic acid, 2-methyl, (3Z)-3-hexen-1-yl | 21.45 | 1.56 | 0.23 | 91 | 1325 |  |
| Odor characteristic: leafy, green, vegetable | |||||||
| 18 | β-farnesene ((E)-7,11-dimethyl-3-methylene-1,6,10-dodecatriene) | 26.59 | 0.14 | 0.02 | 90 | 1458 |  |
| Odor characteristic: woody, green | |||||||
| Site | 1 | 2 | 3 | 4 | 5 | Total | C | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | d | n | d | n | d | N | d | N | d | |||
| Episyrphus balteatus | 150.80 | 0.13 * | 29.78 | 0.15 | 7.00 | 0.04 | 202.40 | 0.15 | 87.89 | 0.14 | 219.62 | 170.83 |
| Sarcophaga spp. | 129.00 | 0.11 | 19.85 | 0.17 | 49.25 | 0.34 | 180.70 | 0.13 | 145.89 | 0.25 | 207.02 | 191.67 |
| Musca domestica | 149.70 | 0.13 | 26.47 | 0.32 | 37.75 | 0.28 | 164.10 | 0.12 | 121.44 | 0.19 | 195.06 | 200.00 |
| Chrysotoxum bicinctum | 97.40 | 0.11 | 0.00 | 0.00 | 0.00 | 0.00 | 110.00 | 0.08 | 52.75 | 0.07 | 172.14 | 116.67 |
| Apis sp. | 116.20 | 0.09 | 4.38 | 0.03 | 9.00 | 0.07 | 138.10 | 0.10 | 6.22 | 0.01 | 127.14 | 170.83 |
| Syrphus ribesii | 93.10 | 0.07 | 4.33 | 0.04 | 4.00 | 0.00 | 118.80 | 0.09 | 27.86 | 0.03 | 124.05 | 154.17 |
| Lucilia caesar | 95.33 | 0.07 | 13.93 | 0.22 | 23.67 | 0.15 | 116.00 | 0.08 | 59.56 | 0.10 | 122.61 | 187.50 |
| Lucilia sericata | 95.11 | 0.06 | 16.00 | 0.03 | 0.00 | 0.00 | 53.00 | 0.04 | 24.50 | 0.03 | 109.03 | 116.67 |
| Eupeodes corollae | 97.75 | 0.05 | 2.00 | 0.03 | 66.00 | 0.08 | 56.60 | 0.04 | 13.00 | 0.01 | 101.79 | 116.67 |
| Eristalis tenax | 76.10 | 0.05 | 6.33 | 0.01 | 0.00 | 0.00 | 58.30 | 0.04 | 13.80 | 0.01 | 98.76 | 116.67 |
| Eristalis pertinax | 72.90 | 0.04 | 5.00 | 0.01 | 0.00 | 0.00 | 38.78 | 0.02 | 17.75 | 0.01 | 89.15 | 104.17 |
| Eristalis intricaria | 53.63 | 0.03 | 0.00 | 0.00 | 0.00 | 0.00 | 25.11 | 0.02 | 84.67 | 0.03 | 86.57 | 83.33 |
| Sphaerophoria scripta | 31.83 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 33.33 | 0.02 | 68.13 | 0.09 | 86.33 | 95.83 |
| Stomoxys calcitrans | 43.67 | 0.01 | 1.50 | 0.01 | 0.00 | 0.00 | 28.80 | 0.02 | 20.25 | 0.01 | 55.13 | 91.67 |
| Cerceris rybyensis | 36.80 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 9.00 | 0.00 | 1.00 | 0.00 | 45.11 | 33.33 |
| Cantharis pellucida | 39.33 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 11.22 | 0.01 | 40.00 | 0.01 | 37.00 | 54.17 |
| Bombus sp. | 24.20 | 0.02 | 2.25 | 0.01 | 8.50 | 0.03 | 28.20 | 0.02 | 6.56 | 0.01 | 33.83 | 145.83 |
| Bombus terrestris | 8.50 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 4.80 | 0.00 | 14.00 | 0.00 | 12.17 | 45.83 |
| Vespula vulgaris | 5.00 | 0.00 | 0.00 | 0.00 | 7.00 | 0.01 | 3.70 | 0.00 | 1.86 | 0.00 | 7.29 | 112.50 |
| Gasteruption assectator | 5.75 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.00 | 0.00 | 0.00 | 0.00 | 6.60 | 37.50 |
| Aglais io | 3.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 3.00 | 20.83 |
| Gaurotes virginea | 2.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.00 | 4.17 |
| Polistes dominula | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 1.00 | 0.00 | 1.50 | 12.50 |
| Gasteruption spp. | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 1.33 | 8.33 |
| Community indices | ||||||||||||
| Site | 1 | 2 | 3 | 4 | 5 | Chi-Square | p | |||||
| Total | 1288.50 | a | 86.07 | b | 142.75 | b | 1362.10 | a | 639.22 | Ab | 33.02 | 0.0001 |
| Species number | 16.50 | a ** | 5.80 | b | 6.00 | b | 18.40 | a | 12.67 | Ab | 37.88 | 0.0001 |
| Berger-Parker dominance (D) | 2.36 | b | 2.49 | b | 2.43 | b | 6.33 | a | 3.95 | B | 33.09 | 0.0001 |
| Margalef (S) | 1.50 | ab | 1.25 | b | 1.01 | b | 2.41 | a | 1.81 | B | 29.57 | 0.0001 |
| 1.23 | b | 1.40 | b | 1.41 | b | 2.43 | a | 1.97 | Ab | 33.63 | 0.0001 | |
| Pielou (J) | 0.44 | b | 0.82 | a | 0.80 | a | 0.84 | a | 0.78 | A | 17.20 | 0.0018 |
| Site | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| 1 | x | 0.52 | 0.50 | 0.92 | 0.86 |
| 2 | x | 0.62 | 0.52 | 0.88 | |
| 3 | x | 0.41 | 0.50 | ||
| 4 | x | 0.92 | |||
| 5 | x |
| Site | 1 | 2 | 3 | 4 | 5 | Total | C | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | d | n | d | n | d | n | d | n | d | |||
| Musca domestica | 170.10 | 0.17 | 233.40 | 0.22 | 212.58 | 0.17 | 119.33 | 0.07 | 115.09 | 0.15 | 171.65 | 150.72 |
| Episyrphus balteatus | 114.92 | 0.17 | 187.64 | 0.19 | 178.50 | 0.15 | 180.33 | 0.16 | 153.25 | 0.23 | 162.51 | 159.46 |
| Apis spp. | 94.75 | 0.13 | 179.18 | 0.20 | 193.42 | 0.14 | 140.75 | 0.11 | 35.58 | 0.04 | 127.88 | 200.00 |
| Sarcophaga spp. | 81.38 | 0.05 | 88.78 | 0.06 | 102.29 | 0.02 | 137.67 | 0.12 | 128.17 | 0.20 | 111.58 | 135.21 |
| Syrphus ribesii | 30.10 | 0.03 | 89.20 | 0.07 | 152.10 | 0.06 | 171.45 | 0.11 | 63.89 | 0.05 | 103.50 | 126.58 |
| Lucilia caesar | 62.08 | 0.08 | 137.56 | 0.11 | 100.55 | 0.07 | 98.73 | 0.09 | 58.80 | 0.06 | 89.87 | 158.21 |
| Sphaerophoria scripta | 5.20 | 0.00 | 8.67 | 0.00 | 122.88 | 0.04 | 167.33 | 0.08 | 62.09 | 0.06 | 83.33 | 100.00 |
| Lucilia sericata | 54.33 | 0.09 | 84.90 | 0.08 | 69.10 | 0.04 | 126.60 | 0.07 | 52.57 | 0.03 | 78.08 | 144.12 |
| Eupeodes corollae | 40.83 | 0.02 | 3.88 | 0.00 | 150.75 | 0.06 | 5.00 | 0.00 | 44.40 | 0.02 | 61.04 | 76.71 |
| Chrysotoxum bicinctum | 95.70 | 0.11 | 4.60 | 0.00 | 58.42 | 0.07 | 70.00 | 0.07 | 8.67 | 0.00 | 60.64 | 116.67 |
| Eristalis tenax | 32.50 | 0.03 | 8.90 | 0.01 | 82.27 | 0.04 | 72.71 | 0.03 | 55.92 | 0.06 | 49.31 | 136.84 |
| Eristalis pertinax | 13.63 | 0.01 | 2.83 | 0.00 | 90.00 | 0.04 | 38.86 | 0.01 | 48.78 | 0.03 | 43.43 | 103.90 |
| Eristalis intricaria | 29.33 | 0.02 | 19.71 | 0.02 | 63.64 | 0.03 | 50.00 | 0.01 | 19.11 | 0.02 | 36.51 | 104.00 |
| Bombus sp. | 18.64 | 0.02 | 11.09 | 0.01 | 50.67 | 0.03 | 60.25 | 0.05 | 20.42 | 0.02 | 32.81 | 193.33 |
| Stomoxys calcitrans | 47.25 | 0.02 | 4.00 | 0.00 | 25.88 | 0.01 | 76.00 | 0.00 | 54.33 | 0.01 | 31.56 | 77.14 |
| Polistes dominula | 5.43 | 0.00 | 8.86 | 0.01 | 23.56 | 0.01 | 4.00 | 0.00 | 12.50 | 0.00 | 13.07 | 84.85 |
| Vespula vulgaris | 7.17 | 0.01 | 10.27 | 0.01 | 9.80 | 0.00 | 4.55 | 0.00 | 4.40 | 0.00 | 7.24 | 166.15 |
| Bombus terrestris | 5.00 | 0.00 | 4.20 | 0.00 | 8.33 | 0.00 | 11.13 | 0.00 | 6.00 | 0.00 | 6.88 | 108.20 |
| Cantharis pellucida | 4.00 | 0.00 | 2.40 | 0.00 | 12.86 | 0.00 | 0.00 | 0.00 | 5.00 | 0.00 | 6.50 | 55.00 |
| Cerceris rybyensis | 4.50 | 0.00 | 7.38 | 0.01 | 5.00 | 0.00 | 4.00 | 0.00 | 2.00 | 0.00 | 5.46 | 77.42 |
| Gaurotes virginea | 3.00 | 0.00 | 1.25 | 0.00 | 5.17 | 0.00 | 14.00 | 0.00 | 0.00 | 0.00 | 3.89 | 46.91 |
| Gasteruption assectator | 2.00 | 0.00 | 1.33 | 0.00 | 0.00 | 0.00 | 2.00 | 0.00 | 1.00 | 0.00 | 1.57 | 22.22 |
| Aglais io | 1.50 | 0.00 | 1.50 | 0.00 | 1.00 | 0.00 | 2.00 | 0.00 | 0.00 | 0.00 | 1.40 | 24.39 |
| Gasteruption spp. | 1.25 | 0.00 | 1.25 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.18 | 34.38 |
| Community indices | ||||||||||||
| Site | 1 | 2 | 3 | 4 | 5 | Chi-Square | p | |||||
| Total | 784.58 | b | 995.64 | b | 1481.25 | a | 1255.25 | a | 777.50 | B | 33.47 | 0.0001 |
| Species number | 17.25 | a | 16.73 | a | 17.00 | a | 12.75 | b | 12.92 | B | 37.88 | 0.0001 |
| Berger-Parker dominance (d) | 2.69 | b | 3.88 | ab | 5.26 | a | 5.58 | a | 4.42 | ab | 33.09 | 0.0001 |
| Margalef (S) | 1.66 | b | 2.28 | a | 2.26 | a | 1.70 | b | 1.83 | B | 29.57 | 0.0001 |
| 1.37 | b | 1.90 | ab | 2.33 | a | 2.21 | a | 2.03 | ab | 33.63 | 0.0001 | |
| Pielou (J) | 1.37 | b | 1.90 | ab | 2.33 | a | 2.21 | a | 2.03 | ab | 33.63 | 0.0001 |
| Total | 0.48 | b | 0.69 | ab | 0.83 | a | 0.87 | a | 0.80 | ab | 17.20 | 0.0017 |
| 1 | 2 | 3 | 4 | 5 | |
|---|---|---|---|---|---|
| 1 | x | 1 | 0.96 | 0.92 | 0.86 |
| 2 | x | 0.96 | 0.92 | 0.92 | |
| 3 | x | 0.96 | 0.83 | ||
| 4 | x | 0.87 | |||
| 5 | x |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakubska-Busse, A.; Dziadas, M.; Gruss, I.; Kobyłka, M.J. Floral Volatile Organic Compounds and a List of Pollinators of Fallopia baldschuanica (Polygonaceae). Insects 2022, 13, 904. https://doi.org/10.3390/insects13100904
Jakubska-Busse A, Dziadas M, Gruss I, Kobyłka MJ. Floral Volatile Organic Compounds and a List of Pollinators of Fallopia baldschuanica (Polygonaceae). Insects. 2022; 13(10):904. https://doi.org/10.3390/insects13100904
Chicago/Turabian StyleJakubska-Busse, Anna, Mariusz Dziadas, Iwona Gruss, and Michał J. Kobyłka. 2022. "Floral Volatile Organic Compounds and a List of Pollinators of Fallopia baldschuanica (Polygonaceae)" Insects 13, no. 10: 904. https://doi.org/10.3390/insects13100904