
Effect of Irradiation on Reproduction of Female Spodoptera litura (Fabr.) (Lepidoptera: Noctuidae) in Relation to the Inherited Sterility Technique
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Culture and Maintenance of Spodoptera litura
2.2. Irradiation Treatment
2.3. Behavioral Study
2.4. GC-MS Analysis
2.5. Sampling, RNA Isolation and cDNA Synthesis
2.6. Measurement of Gene Expression through qPCR
2.7. Statistical Analyses
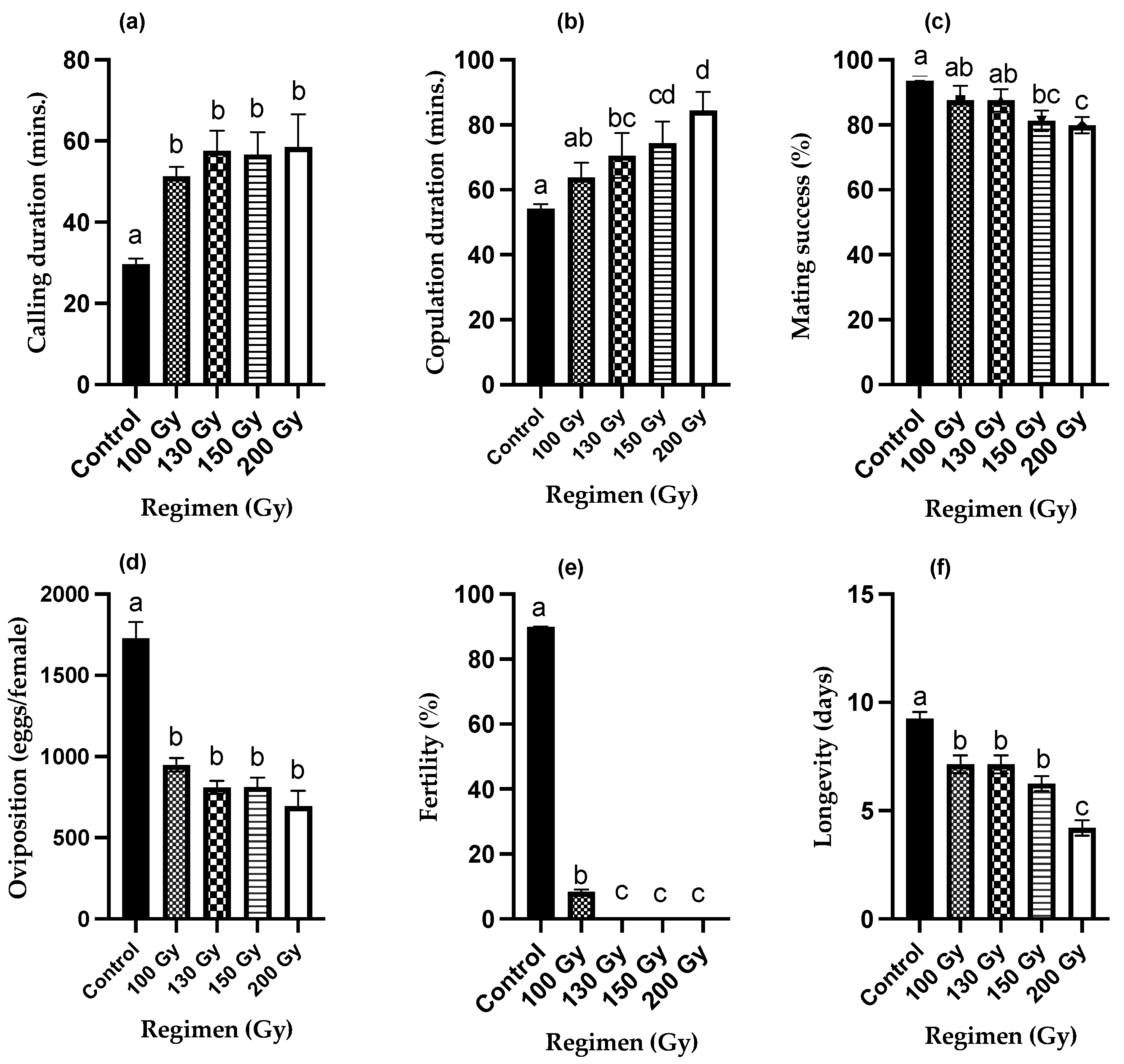
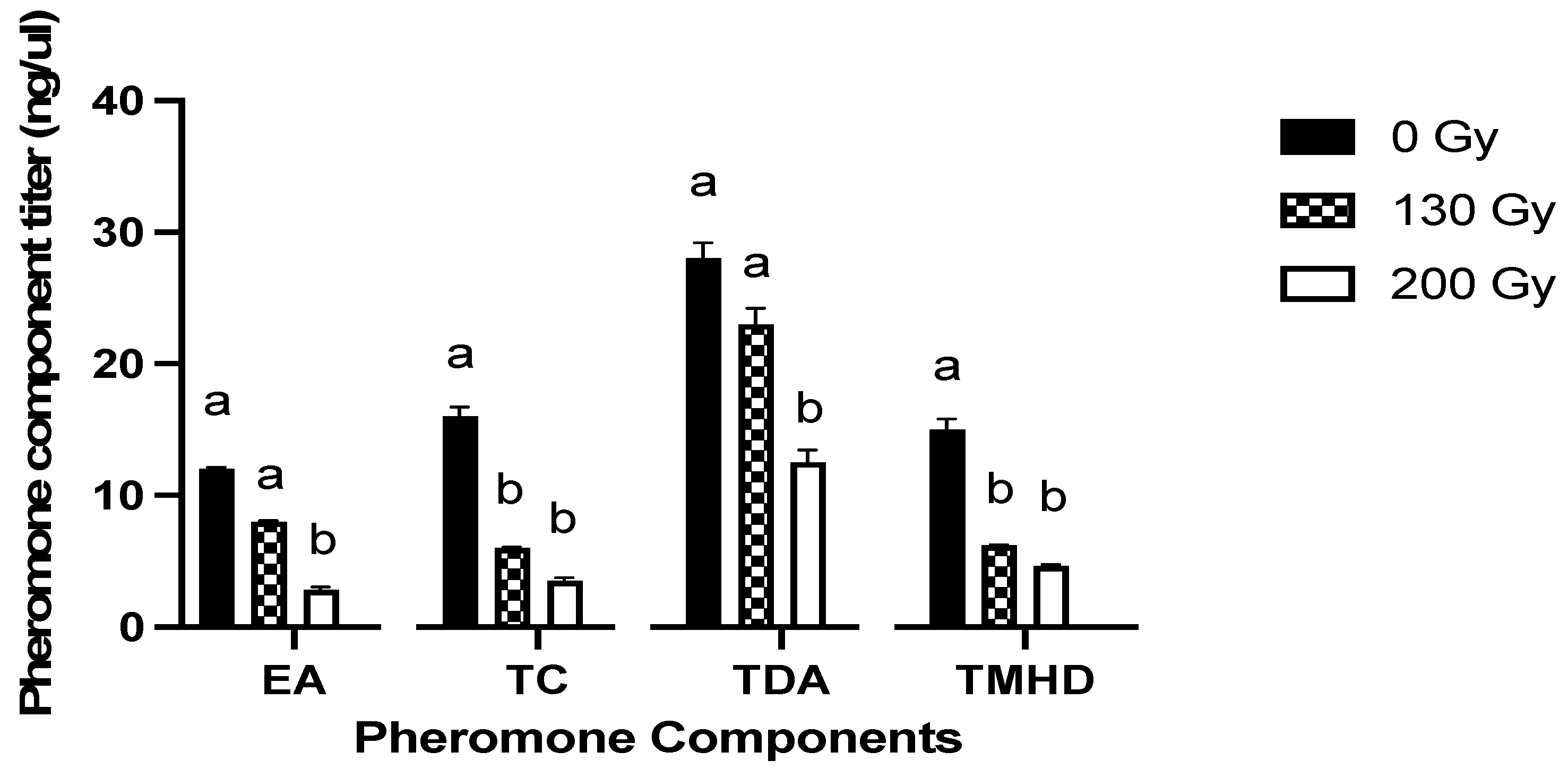
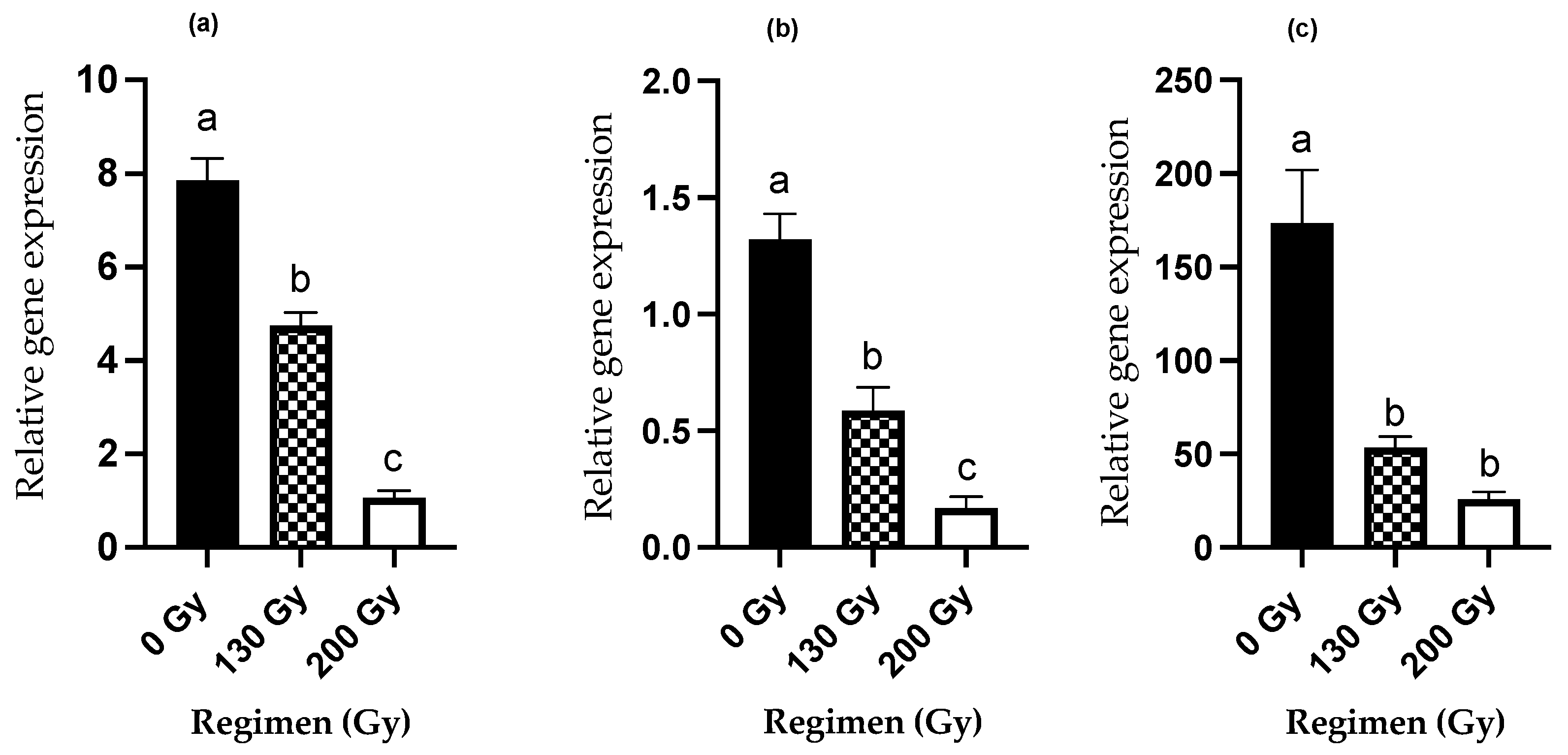
3. Results
3.1. Radiation Effects on Mating Behavior and Reproduction
3.2. GC-MS Analysis of Pheromone Profiles
3.3. Irradiation Effects on Reproductive Genes
3.3.1. PBAN Expression Levels
3.3.2. PBAN-R Expression Levels
3.3.3. Vg Expression Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bernays, E.A. Evolution of feeding behaviour in insect herbivores. Bioscience 1998, 48, 35–44. [Google Scholar] [CrossRef]
- Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.A.; Jaques Miret, J.A.; Justesen, A.F.; Magnusson, C.S.; Milonas, P.; Navas-Cortes, J.A.; et al. EFSA Panel on Plant Health (PLH) Pest categorisation of Spodoptera litura. EFSA J. 2019, 17, e05765. [Google Scholar] [PubMed] [Green Version]
- Armes, N.J.; Wightman, J.A.; Jadhav, D.R.; Ranga Rao, G.V. Status of insecticide resistance in Spodoptera litura in Andhra Pradesh, India. Pestic. Sci. 1997, 50, 240–248. [Google Scholar] [CrossRef]
- Shi, L.; Shi, Y.; Zhang, Y.; Liao, X. A systemic study of indoxacarb resistance in Spodoptera litura revealed complex expression profiles and regulatory mechanism. Sci. Rep. 2019, 9, 14997. [Google Scholar] [CrossRef] [Green Version]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Taylor & Francis: Abingdon, UK, 2021; p. 1216. [Google Scholar]
- Marec, F.; Vreysen, M.J. Advances and challenges of using the sterile insect technique for the management of pest Lepidoptera. Insects 2019, 10, 371. [Google Scholar] [CrossRef] [Green Version]
- LaChance, L.E. Genetic Methods for the Control of Lepidopteran Species: Status and Potential; United States Department of Agriculture, Agricultural Research Service: Washington, DC, USA, 1985; Volume ARS-28, p. 44.
- LaChance, L.E. Dominant lethal mutations in insects with holokinetic chromosomes. 2. Irradiation of sperm of cabbage looper. Ann. Entomol. Soc. Am. 1974, 67, 35–39. [Google Scholar] [CrossRef]
- Marec, F.; Tothová, A.; Sahara, K.; Traut, W. Meiotic pairing of sex chromosome fragments and its relation to atypical transmission of a sex-linked marker in Ephestia kuehniella (Insecta: Lepidoptera). Heredity 2001, 87, 659–671. [Google Scholar] [CrossRef] [PubMed]
- LaChance, L.E.; Graham, C.K. Insect radiosensitivity: Dose curves and dose-fractionation studies of dominant lethal mutations in the mature sperm of 4 insect species. Mutat. Res. 1984, 127, 49–59. [Google Scholar] [CrossRef]
- Carpenter, J.E.; Bloem, S.; Marec, F. Inherited sterility in insects. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 115–146. [Google Scholar]
- Vreysen, M.J.; Hendrichs, J.; Enkerlin, W.R. The sterile insect technique as a component of sustainable area-wide integrated pest management of selected horticultural insect pests. J. Fruit Ornam. Plant Res. 2006, 14, 107. [Google Scholar]
- Vreysen, M.J.B.; Klassen, W.; Carpenter, J.E. Overview of technological advances toward greater efficiency and efficacy in sterile insect-inherited sterility programs against moth pests. Fla. Entomol. 2016, 99, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Traut, W. A study of recombination, formation of chiasmata and synaptonemal complexes in female and male meiosis of Ephestia kuehniella (Lepidoptera). Genetica 1977, 47, 135–142. [Google Scholar] [CrossRef]
- Marec, F.; Kollárová, I.; Pavelka, J. Radiation-induced inherited sterility combined with a genetic sexing system in Ephestia kuehniella (Lepidoptera: Pyralidae). Ann. Entomol. Soc. Am. 1999, 92, 250–259. [Google Scholar] [CrossRef]
- Seth, R.K.; Sehgal, S.S. Partial sterilizing radiation dose effect on the F1 progeny of Spodoptera litura (Fabr.). In Management of Insect Pests: Nuclear and Related Molecular and Genatic Technique, Proceedings of the Symposium Held in Vienna, Vienna, Austria, 19–23 October 1992; IAEA: Vienna, Austria, 1993; pp. 19–23. [Google Scholar]
- Seth, R.K.; Sharma, V.P. Inherited sterility by substerilizing radiation in Spodoptera litura (Lepidoptera: Noctuidae): Bioefficacy and potential for pest suppression. Fla. Entomol. 2001, 84, 183–193. [Google Scholar] [CrossRef]
- Lu, Q.; Huang, L.Y.; Liu, F.T.; Wang, X.F.; Chen, P.; Xu, J.; Deng, J.Y.; Ye, H. Sex pheromone titer in the glands of Spodoptera litura females: Circadian rhythm and the effects of age and mating. Physiol. Entomol. 2017, 42, 156–162. [Google Scholar] [CrossRef]
- Lu, Q.; Huang, L.Y.; Chen, P.; Yu, J.F.; Xu, J.; Deng, J.Y.; Ye, H. Identification and RNA interference of the pheromone biosynthesis activating neuropeptide (PBAN) in the common cutworm moth Spodoptera litura (Lepidoptera: Noctuidae). J. Econ. Entomol. 2015, 108, 1344–1353. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Luo, L.; Jiang, X.; Zhang, L.; Niu, C. Expression of pheromone biosynthesis activating neuropeptide and its receptor (PBANR) mRNA in adult female Spodoptera exigua (Lepidoptera: Noctuidae). Arch. Insect Biochem. Physiol. 2010, 75, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Zhou, J.; Tang, W.; Lu, K.; Zhou, Q.; Zhang, G. Molecular characterization and expression pattern of Spodoptera litura (Lepidoptera: Noctuidae) vitellogenin, and its response to lead stress. J. Insect Physiol. 2009, 55, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Rio, D.C.; Ares, M.; Hannon, G.J.; Nilsen, T.W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb. Protoc. 2010, 6, pdb-prot5439. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Löfstedt, C.; Herrebout, W.M.; Du, J.W. Evolution of the ermine moth pheromone tetradecyl acetate. Nature 1986, 323, 621–623. [Google Scholar] [CrossRef]
- Booth, Y.K.; Kitching, W.; De Voss, J.J. Biosynthesis of insect spiroacetals. Nat. Prod. Rep. 2009, 26, 490–525. [Google Scholar] [CrossRef] [PubMed]
- Jurenka, R.A. Biosynthetic pathway for producing the sex pheromone component (Z, E)-9, 12-tetradecadienyl acetate in moths involves a Δ 12 desaturase. Cell. Mol. Life Sci. 1997, 53, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Cocchiararo-Bastias, L.M.; Mijailovsky, S.J.; Calderon-Fernández, G.M.; Lorenzo Figueiras, A.N.; Juárez, M.P. Epicuticle lipids mediate mate recognition in Triatoma infestans. J. Chem. Ecol. 2011, 37, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Aviles, A.; Cordeiro, A.; Maria, A.; Bozzolan, F.; Boulogne, I.; Dacher, M.; Goutte, A.; Alliot, F.; Maïbèche, M.; Massot, M.; et al. Effects of DEHP on the ecdysteroid pathway, sexual behavior and offspring of the moth Spodoptera littoralis. Horm. Behav. 2020, 125, 104808. [Google Scholar] [CrossRef]
- Böröczky, K.; Wada-Katsumata, A.; Batchelor, D.; Zhukovskaya, M.; Schal, C. Insects groom their antennae to enhance olfactory acuity. Proc. Natl. Acad. Sci. USA 2013, 110, 3615–3620. [Google Scholar] [CrossRef] [Green Version]
- Hoyt, C.P.; Osborne, G.O.; Mulcock, A.P. Production of an insect sex attractant by symbiotic bacteria. Nature 1971, 230, 472–473. [Google Scholar] [CrossRef] [PubMed]
- Böröczky, K.; Crook, D.J.; Jones, T.H.; Kenny, J.C.; Zylstra, K.E.; Mastro, V.C.; Tumlinson, J.H. Monoalkenes as contact sex pheromone components of the woodwasp Sirex noctilio. J. Chem. Ecol. 2009, 35, 1202–1211. [Google Scholar] [CrossRef]
- Caravantes-Villatoro, L.A.; Cruz-Esteban, S.; Rojas, J.C. Cuticular hydrocarbons of Anastrepha obliqua (Diptera: Tephritidae) as influenced by extraction method, natal host, and age. Fla. Entomol. 2021, 104, 289–296. [Google Scholar] [CrossRef]
- Attygalle, A.B.; Cai-Hong, W.U.; Schwarz, J.; Vostrowsky, O.; Hasenfuss, I.; Bestmann, H.J. Sex pheromone of female Myelois cribrella Hübner (Lepidoptera: Pyralidae). J. Chem. Ecol. 1988, 14, 485–494. [Google Scholar] [CrossRef]
- Martinez, T.; Fabrias, G.; Camps, F. Sex pheromone biosynthetic pathway in Spodoptera littoralis and its activation by a neurohormone. J. Biol. Chem. 1990, 265, 1381–1387. [Google Scholar] [CrossRef]
- Wang, H.L.; Zhao, C.H.; Wang, C.Z. Comparative study of sex pheromone composition and biosynthesis in Helicoverpa armigera, H. assulta and their hybrid. Insect Biochem. Mol. Biol. 2005, 35, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Mast, J.D.; De Moraes, C.M.; Alborn, H.T.; Lavis, L.D.; Stern, D.L. Evolved differences in larval social behavior mediated by novel pheromones. Elife 2014, 3, e04205. [Google Scholar] [CrossRef] [PubMed]
- Park, G.W.; Jung, R.; Woon, K. Sex Pheromones as a Tool to Overcome Parnassius bremeri Bremer Shortfall in Conservation Biology. (Lepidoptera: Papilionidae). Proceedings of the Korean Society of Applied Entomology Conference 2016 Regular General Assembly and International Symposium of the Korean Society of Applied Entomology. 2016, p. 93. Available online: https://db.koreascholar.com/article?code=312969 (accessed on 6 September 2022).
- Rouault, J.; Capy, P.; Jallon, J.M. Variations of male cuticular hydrocarbons with geoclimatic variables: An adaptative mechanism in Drosophila melanogaster? Genetica 2000, 110, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Liénard, M.A.; Zhao, C.H.; Wang, C.Z.; Löfstedt, C. Neofunctionalization in an ancestral insect desaturase lineage led to rare Δ6 pheromone signals in the Chinese tussah silkworm. Insect Biochem. Mol. Biol. 2010, 40, 742–751. [Google Scholar] [CrossRef]
- Ananthakrishnan, T.N. Allelochemical synergism and insect behavioural diversity. Curr. Sci. 1997, 72, 628–630. [Google Scholar]
- Charlton, R.E.; Roelofs, W.L. Biosynthesis of a volatile, methly-branched hydrocarbon sex pheromone from leucine by arctiid moths (Holomelina spp.). Arch. Insect Biochem. Physiol. 1991, 18, 81–97. [Google Scholar] [CrossRef]
- Linley, J.R.; Carlson, D.A. A contact mating pheromone in the biting midge, Culicoides melleus. J. Insect Physiol. 1978, 24, 423–427. [Google Scholar] [CrossRef]
- Eom, I.Y.; Risticevic, S.; Pawliszyn, J. Simultaneous sampling and analysis of indoor air infested with Cimex lectularius L. (Hemiptera: Cimicidae) by solid phase microextraction, thin film microextraction and needle trap device. Anal. Chim. Acta 2012, 716, 2–10. [Google Scholar] [CrossRef]
- Seenivasagan, T.; Sharma, K.R.; Sekhar, K.; Ganesan, K.; Prakash, S.; Vijayaraghavan, R. Electroantennogram, flight orientation, and oviposition responses of Aedes aegypti to the oviposition pheromone n-heneicosane. Parasitol. Res. 2009, 104, 827–833. [Google Scholar] [CrossRef]
- Grant, G.G.; Frech, D.; MacDonald, L.; Slessor, K.N.; King, G.G.S. Copulation releaser pheromone in body scales of female whitemarked tussock moth, Orgyia leucostigma (Lepidoptera: Lymantriidae): Identification and behavioral role. J. Chem. Ecol. 1987, 13, 345–356. [Google Scholar] [CrossRef]
- Kittayapong, P.; Kaeothaisong, N.O.; Ninphanomchai, S.; Limohpasmanee, W. Combined sterile insect technique and incompatible insect technique: Sex separation and quality of sterile Aedes aegypti male mosquitoes released in a pilot population suppression trial in Thailand. Parasites Vectors 2018, 11, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vera, M.T.; Cladera, J.L.; Calcagno, G.; Vilardi, J.C.; Mcinnis, D.W. Remating of wild Ceratitis capitata (Diptera: Tephritidae) females in field cages. Ann. Entomol. Soc. Am. 2003, 96, 563–570. [Google Scholar] [CrossRef]
- Mudavanhu, P.; Addison, P.; Carpenter, J.E.; Conlong, D.E. Mating compatibility and competitiveness between wild and laboratory strains of Eldana saccharina (Lepidoptera: Pyralidae) after radiation treatment. Fla. Entomol. 2016, 99 (Suppl. 1), 54–65. [Google Scholar] [CrossRef] [Green Version]
- Walton, A.J.; Conlong, D.E. General biology of Eldana saccharina (Lepidoptera: Pyralidae): A target for the sterile insect technique. Fla. Entomol. 2016, 99 (Suppl. 1), 30–35. [Google Scholar] [CrossRef] [Green Version]
- Chakroun, S.; Rempoulakis, P.; Lebdi-Grissa, K.; Vreysen, M.J. Gamma irradiation of the carob or date moth Ectomyelois ceratoniae: Dose–response effects on egg hatch, fecundity, and survival. Entomol. Exp. Appl. 2017, 164, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Bloem, S.; Bloem, K.A.; Carpenter, J.E.; Calkins, C.O. Inherited sterility in codling moth (Lepidoptera: Tortricidae): Effect of substerilizing doses of radiation on insect fecundity, fertility, and control. Ann. Entomol. Soc. Am. 1999, 92, 222–229. [Google Scholar] [CrossRef]
- Kouloussis, N.A.; Gerofotis, C.D.; Ioannou, C.S.; Iliadis, I.V.; Papadopoulos, N.T.; Koveos, D.S. Towards improving sterile insect technique: Exposure to orange oil compounds increases sexual signalling and longevity in Ceratitis capitata males of the Vienna 8 GSS. PLoS ONE 2017, 12, e0188092. [Google Scholar] [CrossRef] [Green Version]
- Abdu, R.M.; Abdel-Kader, M.M.; Hussein, M.A.; Abdel-Rahman, H.A. Biological effects of gamma radiation on stored product insects. 4-radiation effects on sex pheromone production and perception by the rust-red flour beetle. Tribolium castaneum (herbst). Qatar Univ. Sci. Bull. 1985, 5, 286–289. [Google Scholar]
- Sato, Y.; Ikeda, M.; Yamashita, O. Neurosecretory cells expressing the gene for common precursor for diapause hormone and pheromone biosynthesis-activating neuropeptide in the suboesophageal ganglion of the silkworm, Bombyx mori. Gen. Comp. Endocrinol. 1994, 96, 27–36. [Google Scholar] [CrossRef]
- Sengupta, M.; Angmo, N.; Vimal, N.; Seth, R.K. Effect of ionizing radiation on pheromone biosynthesis activating neuropeptide (PBAN) gene expression and its photosensitive rhythm in female Spodoptera litura (F.). Indian J. Entomol. 2022, 85, 1–6. [Google Scholar] [CrossRef]
- Rafaeli, A. Pheromone biosynthesis activating neuropeptide (PBAN): Regulatory role and mode of action. Gen. Comp. Endocrinol. 2009, 162, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Cheek, A.O.; Brouwer, T.H.; Carroll, S.; Manning, S.; McLachlan, J.A.; Brouwer, M. Experimental evaluation of vitellogenin as a predictive biomarker for reproductive disruption. Environ. Health Perspect. 2001, 109, 681–690. [Google Scholar] [CrossRef]
- Husain, M.; Rasool, K.G.; Tufail, M.; Sutanto, K.D.; Alwaneen, W.S.; Aldawood, A.S. Ultra Violet (UV-B) radiation intrudes Cadra cautella reproductive biology by influencing vitellogenin expression. J. King Saud Univ. Sci. 2022, 34, 102290. [Google Scholar] [CrossRef]
- Hasaballah, A.I. Impact of paternal transmission of gamma radiation on reproduction, oogenesis, and spermatogenesis of the housefly, Musca domestica L. (Diptera: Muscidae). Int. J. Radiat. Biol. 2021, 97, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Seth, R.K.; Khan, Z.; Rao, D.K.; Zarin, M. Flight activity and mating behavior of irradiated Spodoptera litura (Lepidoptera: Noctuidae) males and their F1 progeny for use of inherited sterility in pest management approaches. Fla. Entomol. 2016, 99 (Suppl. 1), 119–130. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| S.No. | Gene | Primer Sequences | Accession No. |
|---|---|---|---|
| 1 | EF1 α | F 5′-GACAAACGTACCCATCGAGAAG-3′ R 5′-GATACCAGCCTCGAACTCAC-3′ | XM_022965580.1 |
| 2 | PBAN | F 5′-CTCGGCAGGACGATGAATTT-3′ R 5′-CTGTTGGTACTCCTGACCATTC-3′ | KP_006328.1 |
| 3 | PBAN-R | F 5′-GTATTCTTCGTGGTGCCTATGT-3′ R 5′-CGAGAGCTTCTTCACTGGATG-3′ | KM_023791.1 |
| 4 | Vg | F 5′-GTTGTCTGCCGGTCGAATAA-3′ R 5′-GACTTTCCTGAGTCTGTGTGAG-3′ | EU_095334.1 |
| Functional Category | Pheromone Components | Reference |
|---|---|---|
| Sex attractants | 1-tetradecyl acetate | [24] |
| 11-oxohexadecanoic acid | [25] | |
| (Z,E)-9,12-Tetradecadienyl acetate | [26] | |
| 1-docosanol acetate | [27] | |
| Bis(2-ethylexyl) phthalate | [28] | |
| Tetracontane | [29] | |
| (E,E,E,E)-squalene | [30] | |
| Hexatriacontane | [31] | |
| 2-methyl octacosane | [32] | |
| Tricosyl acetate | [33] | |
| Sex pheromone precursor | n-Hexadecanoic acid | [34] |
| 9,12-octadecadienoic acid | [35] | |
| 9-octadecanoic-(Z)-methyl ester | [36] | |
| n-Eicosane | [37] | |
| 2-methyl octasane | [38] | |
| (9,12)-hexadecadieonoic acetate | [39] | |
| (Z)-7-hexadecanal | [40] | |
| 1-heptadecane carboxylic acid | [41] | |
| Orientation pheromone | n-docosane | [42] |
| Ovipositor releasor | 1,2-benzenedicarboxylic acid | [43] |
| Heneicosane | [44] | |
| Copulation releasor/terminator | 2,6,10,15-tetramethyl heptadecane | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sengupta, M.; Vimal, N.; Angmo, N.; Seth, R.K. Effect of Irradiation on Reproduction of Female Spodoptera litura (Fabr.) (Lepidoptera: Noctuidae) in Relation to the Inherited Sterility Technique. Insects 2022, 13, 898. https://doi.org/10.3390/insects13100898
Sengupta M, Vimal N, Angmo N, Seth RK. Effect of Irradiation on Reproduction of Female Spodoptera litura (Fabr.) (Lepidoptera: Noctuidae) in Relation to the Inherited Sterility Technique. Insects. 2022; 13(10):898. https://doi.org/10.3390/insects13100898
Chicago/Turabian StyleSengupta, Madhumita, Neha Vimal, Nilza Angmo, and Rakesh Kumar Seth. 2022. "Effect of Irradiation on Reproduction of Female Spodoptera litura (Fabr.) (Lepidoptera: Noctuidae) in Relation to the Inherited Sterility Technique" Insects 13, no. 10: 898. https://doi.org/10.3390/insects13100898