
Myzus persicae Management through Combined Use of Beneficial Insects and Thiacloprid in Pepper Seedlings

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Insecticides
2.2. Acute Toxicity Determination
2.3. Risk Assessment Procedures
2.4. Greenhouse Efficacy Trial
3. Results
3.1. Acute Toxicity of Neonicotinoids to Insects
3.2. Risk Assessment of Pesticides to Beneficial Insects in Field
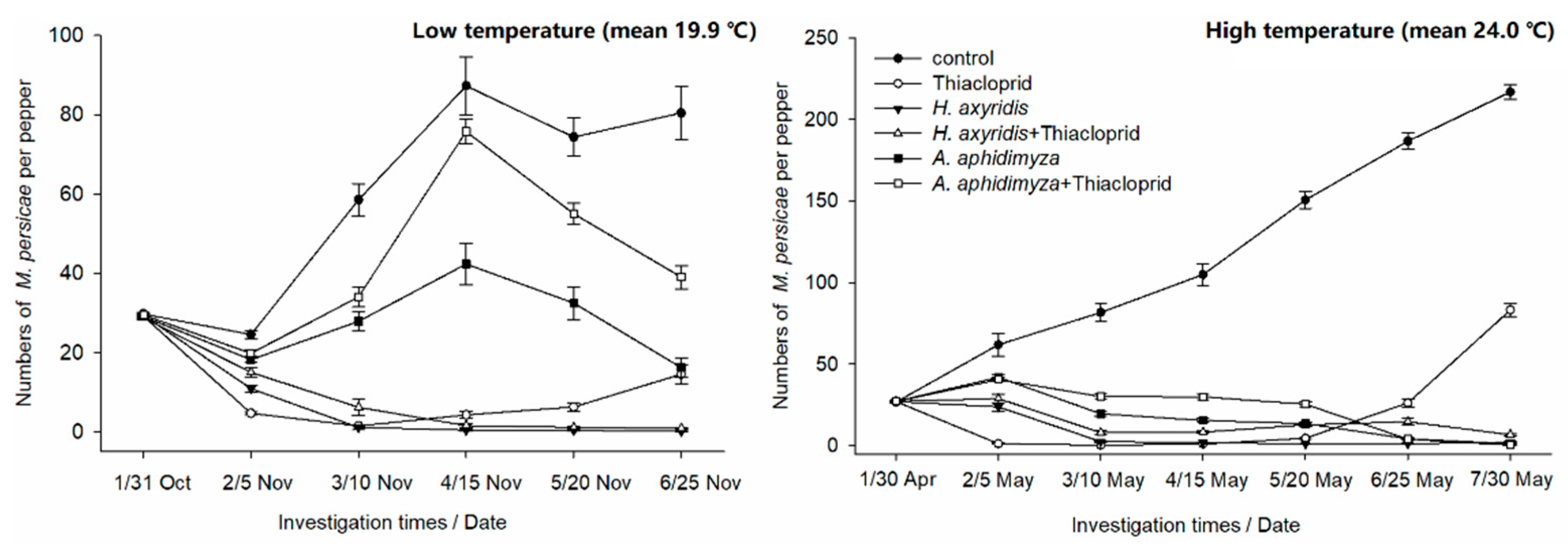
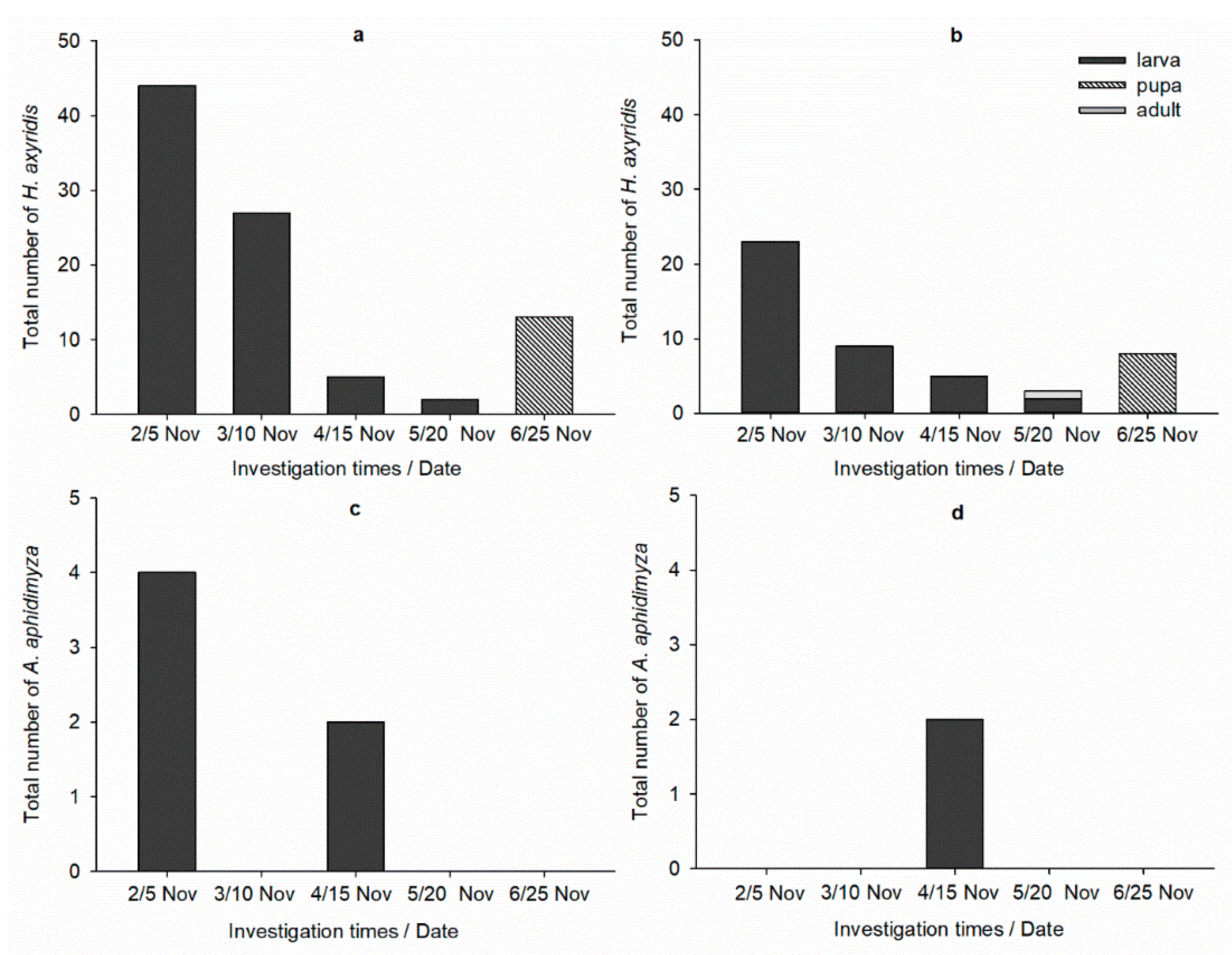
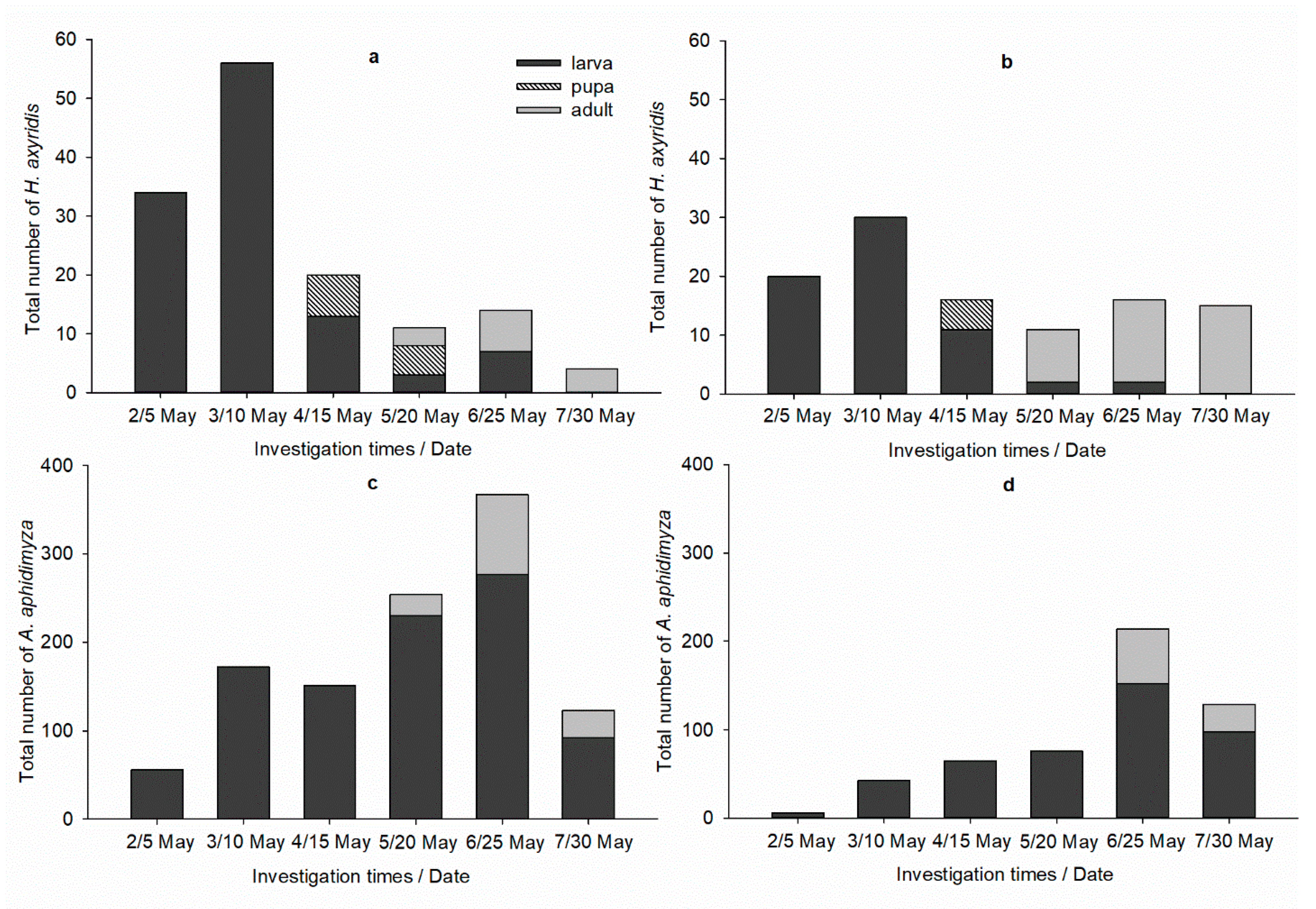
3.3. Greenhouse Efficacy Trial
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Information and Identification Guide; John Wiley & Sons: Chichester, UK, 2000; p. 466. [Google Scholar]
- Bass, C.; Puinean, A.M.; Zimmer, C.T.; Denholm, I.; Field, L.M.; Foster, S.P.; Gutbrod, O.; Nauen, R.; Slater, R.; Williamson, M.S. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 2014, 51, 41–51. [Google Scholar] [CrossRef] [Green Version]
- van Emden, H.F. Integrated pest management and introduction to IPM case studies. In Aphids as Crop Pests; van Emden, H.F., Harrington, R., Eds.; CABI: Wallingford, UK, 2007; Chapter 20; pp. 537–548. [Google Scholar]
- Matsuda, K.; Ihara, M.; Sattelle, D.B. Neonicotinoid insecticides: Molecular targets, resistance, and toxicity. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 241–255. [Google Scholar] [CrossRef]
- Fray, L.M.; Leather, S.R.; Powell, G.; Slater, R.; Mcindoe, E.; Lind, R.J. Behavioural avoidance and enhanced dispersal in neonicotinoid-resistant Myzus persicae (Sulzer). Pest Manag. Sci. 2014, 70, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Crall, J.D.; Switzer, C.M.; Oppenheimer, R.L.; Ford Versypt, A.N.; Dey, B.; Brown, A.; Eyster, M.; Guérin, C.; Pierce, N.E.; Combes, S.A.; et al. Neonicotinoid exposure disrupts bumblebee nest behavior, social networks, and thermoregulation. Science 2018, 362, 683–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sappington, J.D. Imidacloprid alters ant sociobehavioral traits at environmentally relevant concentrations. Ecotoxicology 2018, 27, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, C.A.; Foppen, R.P.B.; Van Turnhout, C.A.M.; De Kroon, H.; Jongejans, E. Declines in insectivorous birds are associated with high neonicotinoid concentrations. Nature 2014, 511, 341–343. [Google Scholar] [CrossRef]
- Goulson, D. Call to restrict neonicotinoids. Science 2018, 360, 973. [Google Scholar] [PubMed]
- Koch, R.L. The multicolored Asian lady beetle, Harmonia axyridis: A review of its biology, uses in biological control, and non-target impacts. J. Insect Sci. 2003, 3, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Riddick, E.W. Spotlight on the positive effects of the ladybird Harmonia axyridis on agriculture. BioControl 2017, 62, 319–330. [Google Scholar] [CrossRef]
- Roy, H.E.; Brown, P.M.J. Ten years of invasion: Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae) in Britain. Ecol. Entomol. 2015, 40, 336–348. [Google Scholar] [CrossRef] [Green Version]
- Boulanger, F.X.; Jandricic, S.; Bolckmans, K.; Wäckers, F.L.; Pekas, A. Optimizing aphid biocontrol with the predator Aphidoletes aphidimyza, based on biology and ecology. Pest Manag. Sci. 2019, 75, 1479–1493. [Google Scholar] [CrossRef] [PubMed]
- Van Schelt, J.; Mulder, S. Improved methods of testing and release of Aphidoletes aphidimyza (Diptera: Cecidomyiidae) for aphid control in glasshouses. Eur. J. Entomol. 2000, 97, 511–516. [Google Scholar] [CrossRef] [Green Version]
- Harris, K.M. Aphidophagous Cecidomyiidae (Diptera): Taxonomy, biology and assessments of field populations. Bull. Entomol. Res. 1973, 63, 305–325. [Google Scholar] [CrossRef]
- Lin, Q.C.; Zhai, Y.F.; Chen, H.; Yin, Y.Y.; Sun, M.; Yu, Y.; Zheng, L. Predatory capacity of Aphidoletes aphidimyza (Rondani). Chin. J. Biol. Control 2017, 33, 171–175. [Google Scholar]
- Jandricic, S.E.; Wraight, S.P.; Gillespie, D.R.; Sanderson, J.P. Biological control outcomes using the generalist aphid predator Aphidoletes aphidimyza under multi-prey conditions. Insects 2016, 7, 75. [Google Scholar] [CrossRef]
- Siviter, H.; Brown, M.J.F.; Leadbeater, E. Sulfoxaflor exposure reduces bumblebee reproductive success. Nature 2018, 561, 109–112. [Google Scholar] [CrossRef]
- Thompson, H.M.; Wilkins, S.; Harkin, S.; Milner, S.; Walters, K.F.A. Neonicotinoids and bumblebees (Bombus terrestris): Effects on nectar consumption in individual workers. Pest Manag. Sci. 2015, 71, 946–950. [Google Scholar] [CrossRef]
- Srigiriraju, L.; Semtner, P.J.; Bloomquist, J.R. Monitoring for imidacloprid resistance in the tobacco-adapted form of the green peach aphid, Myzus persicae (Sulzer) (Hemiptera: Aphididae), in the eastern United States. Pest Manag. Sci. 2010, 66, 676–685. [Google Scholar] [CrossRef]
- Lin, Q.C.; Chen, H.; Babendreier, D.; Zhang, J.P.; Zhang, F.; Dai, X.Y.; Sun, Z.W.; Shi, Z.P.; Dong, X.L.; Wu, G.A.; et al. Improved control of Frankliniella occidentalis on greenhouse pepper through the integration of Orius sauteri and neonicotinoid insecticides. J. Pest Sci. 2021, 94, 101–109. [Google Scholar] [CrossRef]
- Yuan, S.K.; Xu, H.; Qu, W.G.; Shan, Z.J.; Bu, Y.Q.; Yan, Q.P.; Wang, H.L. Environmental Safety Assessment Test Criteria for Chemical Pesticides. Part 10: Acute Toxicity Test for Bees (GB/T31270.10-2014); Standards Press of China: Beijing, China, 2014. [Google Scholar]
- Candolfi, M.P.; Barrett, K.L.; Campbell, P.J.; Forster, R.; Grandy, N.; Huet, M.C.; Lewis, G.; Oomen, P.A.; Schmuck, R.; Vogt, H. Guidance document on regulatory testing and risk assessment procedures for plant protection products with nontarget arthropods. In ESCORT2 Workshop; SETAC-Europe: Wageningen, The Netherlands, 2001. [Google Scholar]
- Lin, R.H.; Yu, C.H.; Jiang, H.; Yuan, S.K.; Ma, X.D.; Li, W.J.; Qu, M.M.; Zhou, Y.M.; Zhou, X.X. Guidance on Environmental Risk Assessment for Pesticide Registration-Part 7: Non-Target Arthropod; Standards Press of China: Beijing, China, 2016. [Google Scholar]
- Utsumi, T.; Miyamoto, M.; Katagi, T. Ecotoxicological risk assessment of pesticides in terrestrial ecosystems. Sumitomo Kagaku 2011, 1, 1–19. [Google Scholar]
- European Food Safety Authority. Conclusion on the peer review of the pesticide risk assessment of the active substance acetochlor. EFSA J. 2011, 9, 2143. [Google Scholar] [CrossRef]
- EPPO (European and Mediterranean Plant Protection Organization). Guidelines for the efficacy evaluation of plant protection products. PP1/170(4): Side effects on honeybees. IOBC/WPRS Bull. 2000, 23, 51–55. [Google Scholar]
- Xu, H.H. Phytochemical Conservation; China Agric. Press: Beijing, China, 2010. [Google Scholar]
- Slater, R.; Paul, V.L.; Andrews, M.; Garbay, M.; Camblin, P. Identifying the presence of neonicotinoidresistant peach-potato aphid (Myzus persicae) in the peach-growing regions of southern France and northern Spain. Pest Manag. Sci. 2012, 68, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Hung, Y.T.; Cheng, Q. A Review of Sub-lethal Neonicotinoid Insecticides Exposure and Effects on Pollinators. Curr. Pollut. Rep. 2020, 6, 137–151. [Google Scholar] [CrossRef]
- Jactel, H.; Verheggen, F.; Thiéry, D.; Escobar-Gutiérrez, A.J.; Gachet, E.; Desneux, N. Alternatives to neonicotinoids. Environ. Int. 2019, 129, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Seko, T.; Miura, K. Functional response of the lady beetle Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae) on the aphid Myzus persicae (Sulzer) (Homoptera: Aphididae). Appl. Entomol. Zool. 2008, 43, 341–345. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Wan, F. Effect of three diets on development and fecundity of the ladybeetles Harmonia axyridis and Propylea japonica. Chin. J. Biol. Control 2001, 17, 116–120. [Google Scholar]
- van Lenteren, J.C. The state of commercial augmentative biological control: Plenty of natural enemies, but a frustrating lack of uptake. BioControl 2012, 57, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Koch, R.L.; Costamagna, A.C. Reaping benefits from an invasive species: Role of Harmonia axyridis in natural biological control of Aphis glycines in North America. BioControl 2017, 62, 331–340. [Google Scholar] [CrossRef]
- Lin, Q.C.; Chen, H.; Yin, Y.Y.; Zhang, S.C.; Yu, Y.; Zhuang, Q.Y.; Zheng, L.; Zhai, Y.F. Effects of temperature on the development and predation of Aphidoletes aphidimyza (Rondani) larvae. Chin. J. Appl. Entomol. 2019, 56, 79–84. [Google Scholar]
- Kim, T.H.; Kim, J.S. Development and adult life span of Aphidoletes aphidimyza (Rondani) (Diptera: Cecidomyiidae) fed on the melon aphid, Aphis gossypii Glover or the green peach aphid, Myzus persicae. Korean J. Appl. Entomol. 2004, 43, 297–304. [Google Scholar]
- Mottaghinia, L.; Hassanpour, M.; Razmjou, J.; Chamani, E.; Hosseini, M. Intraguild predation on the parasitoid wasp Aphidius colemani by the predator Aphidoletes aphidimyza: Effect of host plant cultivars. J. Agric. Sci. Technol. 2018, 20, 533–542. [Google Scholar]
- Gardiner, M.M.; Landis, D.A. Impact of intraguild predation by adult Harmonia axyridis (Coleoptera: Coccinellidae) on Aphis glycines (Hemiptera: Aphididae) biological control in cage studies. Biol. Control 2007, 40, 386–395. [Google Scholar] [CrossRef]
- Hosseini, M.; Ashouri, A.; Enkegaard, A.; Weisser, W.W.; Goldansaz, S.H.; Mahalati, M.N.; Sarraf Moayeri, H.R. Plant quality effects on intraguild predation between Orius laevigatus and Aphidoletes aphidimyza. Entomol. Exp. Appl. 2010, 135, 208–216. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Pesticides | Insects | Slope ± SE | LC50 (mg a.i.·L−1) | 95% Confidence Interval (mg a.i.·L−1) | Correlation Coefficients (r2) |
|---|---|---|---|---|---|
| Imidacloprid | M. persicae | 0.68 ± 0.064 | 1.847 | 1.019–3.304 | 0.900 |
| H. axyridis | 0.79 ± 0.139 | 0.255 | 0.119–0.533 | 0.954 | |
| A. aphidimyza | 1.01 ± 0108 | 0.209 | 0.125–0.318 | 0.906 | |
| Nitenpyram | M. persicae | 0.49 ± 0.053 | 0.808 | 0.392–1.819 | 0.953 |
| H. axyridis | 1.67 ± 0.311 | 17.067 | 10.916–23.720 | 0.981 | |
| A. aphidimyza | 0.79 ± 0.093 | 0.059 | 0.035–0.096 | 0.993 | |
| Acetamiprid | M. persicae | 1.14 ± 0.136 | 25.867 | 17.193–38.068 | 0.994 |
| H. axyridis | 1.06 ± 0.156 | 0.186 | 0.112–0.335 | 0.968 | |
| A. aphidimyza | 1.11 ± 0.124 | 0.049 | 0.028–0.075 | 0.932 | |
| Thiacloprid | M. persicae | 0.43 ± 0.048 | 0.043 | 0.019–0.096 | 0.911 |
| H. axyridis | 1.43 ± 0.222 | 1.314 | 0.935–2.095 | 0.942 | |
| A. aphidimyza | 0.67 ± 0.072 | 0.128 | 0.067–0.232 | 0.995 | |
| Thiamethoxam | M. persicae | 0.45 ± 0.054 | 1.927 | 0.859–4.792 | 0.995 |
| H. axyridis | 0.80 ± 0.126 | 0.916 | 0.461–1.788 | 0.980 | |
| A. aphidimyza | 1.23 ± 0.130 | 0.116 | 0.077–0.168 | 0.928 | |
| Clothianidin | M. persicae | 0.46 ± 0.050 | 0.860 | 0.404–1.887 | 0.975 |
| H. axyridis | 1.04 ± 0.210 | 0.407 | 0.197–1.54 | 0.905 | |
| A. aphidimyza | 0.95 ± 0.109 | 0.061 | 0.035–0.096 | 0.986 | |
| Dinotefuran | M. persicae | 0.74 ± 0.086 | 20.015 | 11.762–38.174 | 0.984 |
| H. axyridis | 0.78 ± 0.154 | 0.864 | 0.383–3.513 | 0.993 | |
| A. aphidimyza | 0.93 ± 0.108 | 0.065 | 0.037–0.102 | 0.982 | |
| Flupyradifurone | M. persicae | 0.43 ± 0.057 | 7.867 | 2.984–30.072 | 0.984 |
| H. axyridis | 1.16 ± 0.260 | 2.489 | 1.326–3.011 | 0.979 | |
| A. aphidimyza | 0.73 ± 0.078 | 0.340 | 0.206–0.610 | 0.992 |
| Pesticides | Hours | Slope ± SE | LD50 (µg a.i.·bee−1) | 95% Confidence Interval (µg a.i.·bee−1) | Correlation Coefficients (r2) |
|---|---|---|---|---|---|
| Nitenpyram | 24 | 0.64 ± 0.073 | 0.592 | 0.314–1.229 | 0.975 |
| 48 | 0.79 ± 0.151 | 0.565 | 0.218–1.435 | ||
| Thiacloprid | 24 | 1.27 ± 0.251 | 19.825 | 9.254–30.352 | 0.981 |
| 48 | 0.85 ± 0.105 | 17.351 | 6.626–28.624 |
| Pesticides | DT50 (Days) | Number of Applications | Application Interval (Days) | Recommended Application Rates (g a.i.·ha−1) | MAF | PER In-Field (g a.i.·ha−1) | LR50 (g a.i.·ha−1) | HQ In-Field | Risk | Insects |
|---|---|---|---|---|---|---|---|---|---|---|
| Imidacloprid | 10 | 2 | 7 | 63.06 | 1.62 | 101.88 | 2.80 | 36.40 | high | H. axyridis |
| 2.29 | 44.49 | high | A. aphidimyza | |||||||
| Nitenpyram | 10 | 3 | 10 | 29.99 | 1.75 | 52.48 | 187.45 | 0.28 | low | H. axyridis |
| 0.65 | 80.99 | high | A. aphidimyza | |||||||
| Acetamiprid | 10 | 1 | 365 | 29.99 | 1.00 | 29.99 | 2.04 | 14.67 | high | H. axyridis |
| 0.53 | 56.18 | high | A. aphidimyza | |||||||
| Thiacloprid | 10 | 2 | 7 | 9.00 | 1.62 | 14.54 | 14.43 | 1.01 | low | H. axyridis |
| 1.04 | 10.37 | high | A. aphidimyza | |||||||
| Thiamethoxam | 10 | 2 | 7 | 56.31 | 1.62 | 90.97 | 10.06 | 9.04 | high | H. axyridis |
| 1.28 | 71.22 | high | A. aphidimyza | |||||||
| Clothianidin | 10 | 1 | 365 | 48.00 | 1.00 | 48.00 | 4.47 | 10.74 | high | H. axyridis |
| 0.67 | 71.18 | high | A. aphidimyza | |||||||
| Dinotefuran | 10 | 2 | 7 | 120.12 | 1.62 | 194.06 | 9.49 | 20.45 | high | H. axyridis |
| 0.71 | 272.68 | high | A. aphidimyza | |||||||
| Flupyradifurone | 10 | 2 | 7 | 102.00 | 1.62 | 164.79 | 27.34 | 6.03 | high | H. axyridis |
| 3.74 | 44.09 | high | A. aphidimyza |
| Pesticides | Hours | Recommended Application Rates (g a.i.·ha−1) | LD50 (g a.i.·bee−1) | HQ In-Field | Risk |
|---|---|---|---|---|---|
| Nitenpyram | 24 | 29.99 | 0.592 | 50.66 | medium risk |
| 48 | 0.565 | 53.08 | medium risk | ||
| Thiacloprid | 24 | 9 | 19.825 | 0.45 | low risk |
| 48 | 17.351 | 0.52 | low risk |
| Control Measures | Investigation Times | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 (5th) | 3 (10th) | 4 (15th) | 5 (20th) | 6 (25th) | |||||||
| Reduction Rate ± SE | Control Effect | Reduction Rate ± SE | Control Effect | Reduction Rate ± SE | Control Effect | Reduction Rate ± SE | Control Effect | Reduction Rate ± SE | Control Effect | ||
| Control | L(Nov.) | 17.44 ± 0.18 | - | −96.30 ± 26.45 | - | −191.42 ± 88.78 | - | −148.79 ± 57.77 | - | −169.00 ± 68.42 | - |
| H(May) | −130.85 ± 9.31 | - | −204.41 ± 6.95 | - | −291.00 ± 18.06 | - | −461.39 ± 6.88 | - | −595.28 ± 28.24 | - | |
| Thiacloprid | L(Nov.) | 83.98 ± 3.22 | 80.63 | 94.85 ± 1.30 | 97.38 | 85.38 ± 1.72 | 94.98 | 78.51 ± 1.44 | 91.36 | 50.20 ± 12.08 | 81.45 |
| H(May) | 94.84 ± 0.57 | 97.76 | 98.16 ± 0.22 | 99.39 | 95.95 ± 0.67 | 98.96 | 82.56 ± 2.34 | 96.89 | 2.91 ± 10.65 | 86.04 | |
| H. axyridis | L(Nov.) | 63.00 ± 4.23 | 55.24 | 96.13 ± 2.30 | 98.03 | 98.58 ± 0.30 | 99.51 | 98.69 ± 0.75 | 99.47 | 99.55 ± 0.12 | 99.83 |
| H(May) | 12.08 ± 2.87 | 61.89 | 90.13 ± 1.66 | 96.76 | 92.40 ± 1.84 | 98.06 | 95.65 ± 2.14 | 99.22 | 95.03 ± 2.63 | 99.29 | |
| H. axyridis + Thiacloprid | L(Nov.) | 47.91 ± 1.80 | 36.91 | 78.39 ± 10.51 | 88.99 | 94.58 ± 0.76 | 98.13 | 96.11 ± 0.45 | 98.44 | 96.87 ± 0.15 | 98.84 |
| H(May) | −11.49 ± 5.05 | 51.68 | 71.94 ± 0.73 | 90.78 | 68.94 ± 1.54 | 92.06 | 51.85 ± 1.85 | 91.42 | 45.01 ± 3.10 | 92.10 | |
| A. aphidimyza | L(Nov.) | 37.48 ± 0.42 | 24.28 | 4.24 ± 21.91 | 51.22 | −45.10 ± 99.25 | 50.21 | −11.22 ± 92.09 | 55.30 | 44.62 ± 51.81 | 79.41 |
| H(May) | −59.30 ± 6.26 | 30.95 | 16.41 ± 7.80 | 72.55 | 37.12 ± 2.49 | 83.94 | 50.12 ± 4.74 | 91.11 | 93.03 ± 1.31 | 99.00 | |
| A. aphidimyza + Thiacloprid | L(Nov.) | 32.86 ± 0.36 | 18.68 | −15.43 ± 14.86 | 41.20 | −157.07 ± 1.49 | 11.79 | −86.57 ± 30.09 | 25.01 | −32.17 ± 53.64 | 50.87 |
| H(May) | −49.50 ± 10.25 | 35.20 | −11.15 ± 6.35 | 63.50 | −10.73 ± 6.65 | 71.70 | 6.11 ± 6.74 | 83.27 | 84.64 ± 2.64 | 97.79 | |
| Investigation Times | Factor | df | F | p |
|---|---|---|---|---|
| 1 | Treatments | 5 | 0.13 | 0.98 |
| Temp | 1 | 74.43 | <0.0001 | |
| Treatments * Temp | 5 | 0.61 | 0.69 | |
| 2 | Treatments | 5 | 63.42 | <0.0001 |
| Temp | 1 | 187.99 | <0.0001 | |
| Treatments * Temp | 5 | 15.35 | <0.0001 | |
| 3 | Treatments | 5 | 247.21 | <0.0001 |
| Temp | 1 | 6.30 | 0.012 | |
| Treatments * Temp | 5 | 9.45 | <0.0001 | |
| 4 | Treatments | 5 | 238.10 | <0.0001 |
| Temp | 1 | 15.97 | <0.0001 | |
| Treatments * Temp | 5 | 22.81 | <0.0001 | |
| 5 | Treatments | 5 | 540.47 | <0.0001 |
| Temp | 1 | 19.46 | <0.0001 | |
| Treatments * Temp | 5 | 105.14 | <0.0001 | |
| 6 | Treatments | 5 | 613.78 | <0.0001 |
| Temp | 1 | 71.72 | <0.0001 | |
| Treatments * Temp | 5 | 143.99 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Q.; Chen, H.; Dai, X.; Yin, S.; Shi, C.; Yin, Z.; Zhang, J.; Zhang, F.; Zheng, L.; Zhai, Y. Myzus persicae Management through Combined Use of Beneficial Insects and Thiacloprid in Pepper Seedlings. Insects 2021, 12, 791. https://doi.org/10.3390/insects12090791
Lin Q, Chen H, Dai X, Yin S, Shi C, Yin Z, Zhang J, Zhang F, Zheng L, Zhai Y. Myzus persicae Management through Combined Use of Beneficial Insects and Thiacloprid in Pepper Seedlings. Insects. 2021; 12(9):791. https://doi.org/10.3390/insects12090791
Chicago/Turabian StyleLin, Qingcai, Hao Chen, Xiaoyan Dai, Shuyan Yin, Chenghao Shi, Zhenjuan Yin, Jinping Zhang, Feng Zhang, Li Zheng, and Yifan Zhai. 2021. "Myzus persicae Management through Combined Use of Beneficial Insects and Thiacloprid in Pepper Seedlings" Insects 12, no. 9: 791. https://doi.org/10.3390/insects12090791