
Ophelimus bipolaris sp. n. (Hymenoptera, Eulophidae), a New Invasive Eucalyptus Pest and Its Host Plants in China †


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Sampling
2.2. Species Identification
2.3. Photography
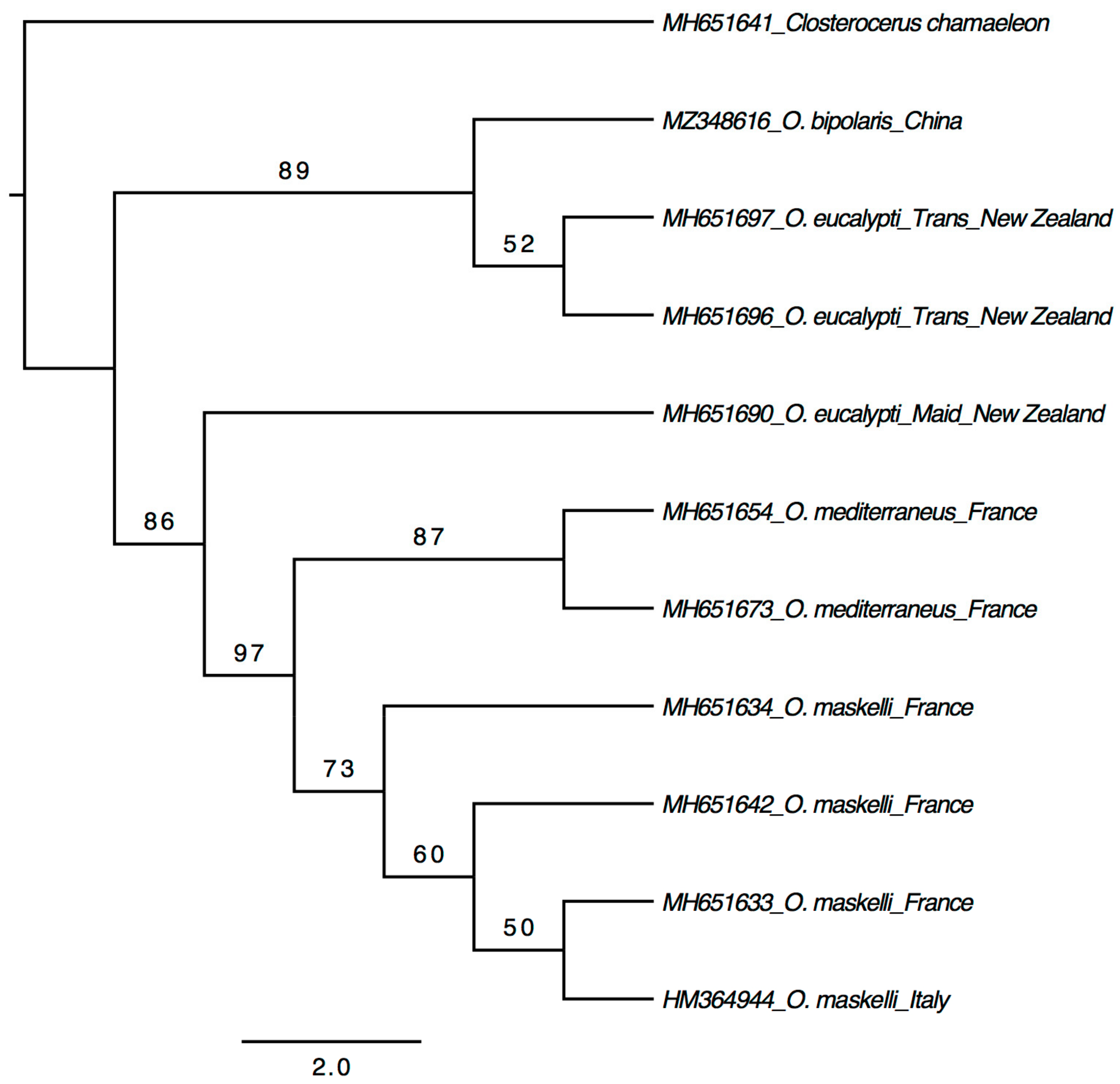
3. Results
Ophelimus bipolaris Chen & Yao, sp. n.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ciesla, V.M. The role of human activities on forest insect outbreaks worldwide. Int. For. Rev. 2015, 17, 269–281. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, Z.; Yan, H. Australian tress in China. In Proceedings of the Australian Tree Species Research in China, Zhangzhou, China, 2–5 November 1992; Brown, A.G., Ed.; Australian Centre or International Agricultural Research: Canberra, Australia, 1994; pp. 19–25. [Google Scholar]
- Arnold, R.J.; Xie, Y.J.; Luo, J.Z.; Wang, H.R.; Midgley, S.J. A tale of two genera: Exotic Eucalyptus and Acacia species in China. 2. Plantation resource development. Int. For. Rev. 2020, 22, 153–168. [Google Scholar]
- Xie, Y.; Arnold, R.J.; Wu, Z.; Chen, S.; Du, A.; Luo, J. Advances in eucalypt research in China. Front. Agric. Sci. Eng. 2017, 4, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.P.; Zeng, L.Q.; Lin, H.Z.; Lin, X.B.; Song, H.T.; Yin, L.Q.; Huang, S.J. Five defoliators on Ecualypts in China. Fujianlinye 2020, 3, 28–32+48. [Google Scholar]
- Tang, C.; Wan, X.J.; Wan, F.H.; Ren, S.X.; Peng, Z.Q. The blue gum chalcid, Leptocybe invasa, invaded Hainan province. Chin. J. Entomol. 2008, 45, 967–971. [Google Scholar]
- Nugnes, F.; Gebiola, M.; Monti, M.M.; Gualtieri, L.; Giorgini, M.; Wang, J.; Bernardo, U. Genetic diversity of the invasive gall wasp Leptocybe invasa (Hymenoptera: Eulophidae) and of its Rickettsia endosymbiont, and associated sex-ratio differences. PLoS ONE 2015, 10, e0124660. [Google Scholar]
- Mendel, Z.; Protasov, A.; Fisher, N.; La Salle, J. Taxonomy and biology of Leptocybe invasa gen. & sp. n. (Hymenoptera: Eulophidae), an invasive gall inducer on Eucalyptus. Aust. J. Entomol. 2004, 43, 101–113. [Google Scholar]
- Borowiec, N.; La Salle, J.; Brancaccio, L.; Thaon, M.; Warot, S.; Branco, M.; Ris, N.; Malausa, J.C.; Burks, R. Ophelimus mediterraneus sp. n. (Hymenoptera, Eulophidae): A new Eucalyptus gall wasp in the Mediterranean region. Bull. Entomol. Res. 2019, 109, 678–694. [Google Scholar] [CrossRef]
- Molina-Mercader, G.; Angulo, A.O.; Olivares, T.S.; Sanfuentes, E.; Castillo-Salazar, M.; Rojas, E.; Toro-Núñez, O.; Benítez, H.A.; Hasbún, R. Ophelimus migdanorum Molina-Mercader sp. nov. (Hymenoptera: Eulophidae): Application of Integrative Taxonomy for Disentangling a Polyphenism Case in Eucalyptus globulus Labill Forest in Chile. Forests 2019, 10, 720. [Google Scholar] [CrossRef] [Green Version]
- Bouček, Z. Australasian Chalcidoidea (Hymenoptera). A Biosystematics Revision of Genera of Fourteen Families, with a Reclassification of Species; CAB International: Wallingford, UK, 1988. [Google Scholar]
- La Salle, J. Biology of gall inducers and evolution of gall induction in Chalcidoidea (Hymenoptera: Eulophidae, Eurytomidae, Pteromalidae, Tanaostigmatidae, Torymidae). In Biology, Ecology and Evolution of Gall-Inducing Arthropods; Raman, A., Schaefer, C.W., Withers, T.M., Eds.; Science Publishers: Enfield, UK, 2005; Volume 2, pp. 503–533. [Google Scholar]
- Withers, T.M.; Raman, A.; Berry, J.A. Host range and biology of Ophelimus eucalypti (Gahan) (Hym.: Eulophidae), a pest of New Zealand eucalypts. N. Z. Plant Prot. 2000, 53, 339–344. [Google Scholar]
- Protasov, A.; La Salle, J.; Blumberg, D.; Brand, D.; Saphir, N.; Assael, F.; Fisher, N.; Mendel, Z. Biology, revised taxonomy and impact on host plants of Ophelimus maskelli, an invasive gall inducer on Eucalyptus spp. in the Mediterranean area. Phytoparasitica 2007, 25, 50–76. [Google Scholar] [CrossRef]
- Dittrich-Schröder, G.; Hurley, B.P.; Wingfield, M.J.; Nahrung, H.F.; Slippers, B. Invasive gall-forming wasps that threaten non-native plantation-grown Eucalyptus: Diversity and invasion patterns. Agric. For. Entomol. 2020, 22, 285–297. [Google Scholar] [CrossRef]
- Gahan, A.B. A list of phytophagous Chalcidoidea with descriptions of two new species. Proc. Entomol. Soc. Wash. 1922, 24, 33–58. [Google Scholar]
- Mendel, Z.; Protasov, A.; Blumberg, D.; Brand, D.; Saphir, N.; Madar, Z.; La Salle, J. Release and recovery of parasitoids of the Eucalyptus gall wasp Ophelimus maskelli in Israel. Phytoparasitica 2007, 35, 330–332. [Google Scholar] [CrossRef]
- Gibson, G.A.P.; Huber, J.T.; Woolley, J.B. Annotated Keys to the Genera of Nearctic Chalcidoidea (Hymenoptera); NRC Research Press: Ottawa, ON, Canada, 1997. [Google Scholar]
- Taekul, C.; Valerio, A.A.; Austin, A.D.; Klompen, H.; Johnson, N.F. Molecular phylogeny of telenomine egg parasitoids (Hymenoptera: Platygastridae s.l.: Telenominae): Evolution of host shifts and implications for classification. Syst. Entomol. 2014, 39, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Gillespie, J.J.; Munro, J.B.; Heraty, J.M.; Yoder, M.J.; Owen, A.K.; Carmichael, A.E. A Secondary Structural Model of the 28S rRNA Expansion Segments D2 and D3 for Chalcidoid Wasps (Hymenoptera: Chalcidoidea). Mol. Biol. Evol. 2005, 22, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- McLaren, P. Ophelimus eucalypti: A recently introduced gall-making insect on eucalypts. Tree Grower May 1989, 10, 26–27. [Google Scholar]
- Clark, A.F. A survey of the insect pests of eucalypts in New Zealand. NZ J. Sci. Tech. 1938, 19, 750–761. [Google Scholar]
- Heimpel, G.E.; de Boer, J.G. Sex determination in the Hymenoptera. Annu. Rev. Entomol. 2008, 53, 209–230. [Google Scholar] [CrossRef] [PubMed]
- Heraty, J.M.; Woolley, J.B.; Hopper, K.R.; Hawks, D.L.; Kim, J.W.; Buffington, M. Molecular phylogenetics and reproductive incompatibility in a complex of cryptic species of aphid parasitoids. Mol. Phylogenet. Evol. 2007, 45, 480–493. [Google Scholar] [CrossRef] [PubMed]
- Gebiola, M.; Bernardo, U.; Burks, R. A reevaluation of the generic limits of Pnigalio Schrank (Hymenoptera: Eulophidae) based on molecular and morphological evidence. Zootaxa 2010, 2484, 35–44. [Google Scholar] [CrossRef]
- Gebiola, M.; Bernardo, U.; Monti, M.M.; Navone, P.; Viggiani, G. Pnigalio agraules (Walker) and Pnigalio mediterraneus Ferrière and Delucchi (Hymenoptera: Eulophidae): Two closely related valid species. J. Nat. Hist. 2009, 43, 2465–2480. [Google Scholar] [CrossRef]
- Dlugosch, K.M.; Parker, I.M. Founding events in species invasions: Genetic variation, adaptive evolution, and the role of multiple introductions. Mol. Ecol. 2008, 17, 431–449. [Google Scholar] [CrossRef]
- Johnstone, R.A.; Hurst, G.D. Maternally inherited male-killing microorganisms may confound interpretation of mitochondrial DNA variability. Biol. J. Linn. Soc. 1996, 58, 453–470. [Google Scholar] [CrossRef]
- Gueguen, G.; Vavre, F.; Gnankine, O.; Peterschmitt, M.; Charif, D.; Chiel, E.; Gottlieb, Y.; Ghanim, M.; Zchori-Fein, E.; Fleury, F. Endosymbiont metacommunities, mtDNA diversity and the evolution of the Bemisia tabaci (Hemiptera: Aleyrodidae) species complex. Mol. Ecol. 2010, 19, 4365–4378. [Google Scholar] [CrossRef]
- Burks, R.A.; Mottern, J.L.; Waterworth, R.; Paine, T.D. First report of the Eucalyptus gall wasp, Ophelimus maskelli (Hymenoptera: Eulophidae), an invasive pest on Eucalyptus, from the Western Hemisphere. Zootaxa 2015, 3926, 448–450. [Google Scholar] [CrossRef] [Green Version]
- Colinet, H.; Boivin, G.; Hance, T. Manipulation of parasitoid size using the temperature-size rule: Fitness consequences. Oecologia 2007, 152, 425–433. [Google Scholar] [CrossRef]
- Garcia, A.; Goncalves, H.; Borowiec, N.; Franco, J.C.; Branco, M. Ophelimus sp., a new invasive gall wasp of Eucalyptus globulus in Europe, escapes the parasitism by Closterocerus chamaeleon due to an asynchronous life cycle. Biol. Control. 2019, 131, 1–7. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Coordinates | Eucalyptus Species |
|---|---|---|
| Guangdong Eco-Engineering Polytechnic (GEEP) | 23°11′58″ N 113°22′35″ E | E. urophylla * |
| Xiaoguwei Island (XI) | 23°4′0″ N 113°22′41″ E | E. urophylla *, E. grandis × E. urophylla *, E. citriodora, E. exserta |
| South China Botanical Garden (SCBG) | 23°10′52″ N 113°21′28″ E | E. tereticornis *, E. citriodora, E. exserta |
| Huolushan Forest Park (HFP) | 23°10′39″ N 113°22′56″ E | E. urophylla *, E. grandis *, E. exserta |
| Code | Locality | Host Plant | Stage | Sex | Submarginal Vein Setae | GenBank Accession Number | |
|---|---|---|---|---|---|---|---|
| COI | 28S | ||||||
| HC739 | GEEP | E. urophylla | adult | female | 4 | MZ348610 | MZ348616 |
| HC740 | GEEP | E. urophylla | larva | NA | NA | MZ348611 | MZ348617 |
| HC745 | GEEP | E. urophylla | adult | male | 3 | MZ348612 | MZ348618 |
| HC746 | GEEP | E. urophylla | adult | male | 4 | MZ348613 | MZ348619 |
| HC747 | GEEP | E. urophylla | adult | female | 5 | MZ348614 | MZ348620 |
| HC785 | XI | E. grandis × E. urophylla | adult | female | 4 | MZ605326 | MZ605354 |
| HC786 | XI | E. grandis × E. urophylla | larva | NA | NA | MZ605327 | MZ605355 |
| HC787 | XI | E. urophylla | adult | female | 5 | MZ605328 | MZ605356 |
| HC788 | XI | E. urophylla | adult | female | 4 | MZ605329 | MZ605357 |
| HC789 | HFP | E. grandis | adult | female | 4 | MZ605330 | MZ605358 |
| HC790 | HFP | E. grandis | adult | female | 3 | MZ605331 | MZ605359 |
| HC791 | HFP | E. urophylla | adult | female | 5 | MZ605332 | MZ605360 |
| HC792 | HFP | E. urophylla | adult | female | 4 | MZ605333 | MZ605361 |
| HC793 | XI | E. urophylla | adult | male | 3 | MZ605334 | MZ605362 |
| HC794 | SCBG | E. tereticornis | adult | female | 4 | MZ605335 | MZ605363 |
| O. bipolaris | O. eucalypti ‘Maid.’ | O. eucalypti ‘Trans.’ | O. maskelli | O. mediterraneus | O. migdnorum | |
|---|---|---|---|---|---|---|
| Head color | Brown with metallic green | Mainly black, frons faintly tinged with metallic green | Mainly black, frons faintly tinged with metallic green | Brown with metallic green | Brown with metallic green | Brown with metallic green |
| Mesosoma color | Brown with metallic green | Mainly black, dorsal mesosoma faintly tinged with purplish | Mainly black, dorsal mesosoma faintly tinged with purplish | Brown with metallic green | Brown with metallic green | Brown with metallic green |
| Body length | Female: 1.1–1.8 mm; male: 1.0–1.2 mm | Female: 2.0–2.5 mm | Female: 2.0–2.5 mm; male: ? | Female: 0.8–1.1 mm | Female: 0.8–1.0 mm | Female: 0.7–1.4 mm; male: 0.7–1.4 mm |
| No. of setae on submarginal vein | 3–5 | 2–4 | ≥5 | 1 | 2–4 | 1–3 |
| Marginal vein/Stigmal vein | About 1.8 × | About 2 × | About 2 × ? | About 0.7× | About 0.7× | 0.7–0.8× |
| Postmarginal vein/Stigmal vein | <0.5× | >1× | >1 × ? | >2× | >1× | >3× |
| No. of setae on mesoscutal midlobe | 5 pairs | 6 pairs | 6 pairs? | 2 pairs | 2 pairs | Unknown |
| Propodeum vs. Metascutellum | Distinctly longer | Subequal | Subequal? | Subequal | Subequal | Unknown |
| Hosts | E. grandis, E. grandis × urophylla, E. tereticornis and E. urophylla | Eucalyptu from section Maidenaria | Eucalyptu from section Transversaria | Eucalyptu from 3 sections: Exsertaria, Latoangulata and Maidenaria | Eucalyptu from section Maidenaria | E. globulus, E. camaldulensis |
| Galls | Only on leaf blade, round and smooth, green then to reddish galls visible on both sides of the leaves | On leaf midribs, leaf blade and shoot axes, round and smooth, green then to reddish galls | Only on leaf blade, females induce circular, protruding galls, males induce pit galls | Only on leaf blade, round and smooth, green then to reddish galls visible on both sides of the leaves | Only on leaf blade, ellipsoidal, conical shaped, brown coloured with rough and racked surface on just the upper side of the leaves | On leaf blade, midrib, secondary rib, petiole, and twigs, amorphous |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.-Y.; Yao, J.-M.; Huang, S.-B.; Pang, H. Ophelimus bipolaris sp. n. (Hymenoptera, Eulophidae), a New Invasive Eucalyptus Pest and Its Host Plants in China. Insects 2021, 12, 778. https://doi.org/10.3390/insects12090778
Chen H-Y, Yao J-M, Huang S-B, Pang H. Ophelimus bipolaris sp. n. (Hymenoptera, Eulophidae), a New Invasive Eucalyptus Pest and Its Host Plants in China. Insects. 2021; 12(9):778. https://doi.org/10.3390/insects12090778
Chicago/Turabian StyleChen, Hua-Yan, Jie-Min Yao, Shao-Bin Huang, and Hong Pang. 2021. "Ophelimus bipolaris sp. n. (Hymenoptera, Eulophidae), a New Invasive Eucalyptus Pest and Its Host Plants in China" Insects 12, no. 9: 778. https://doi.org/10.3390/insects12090778