
Weeds Enhance Pollinator Diversity and Fruit Yield in Mango


Abstract
:Simple Summary
Abstract

1. Introduction
- How do the abundance and diversity of insect species (potential pollinators) differ on mango in the presence or absence of weeds?
- What is the impact of weeds on mango fruit yield?
2. Materials and Methods
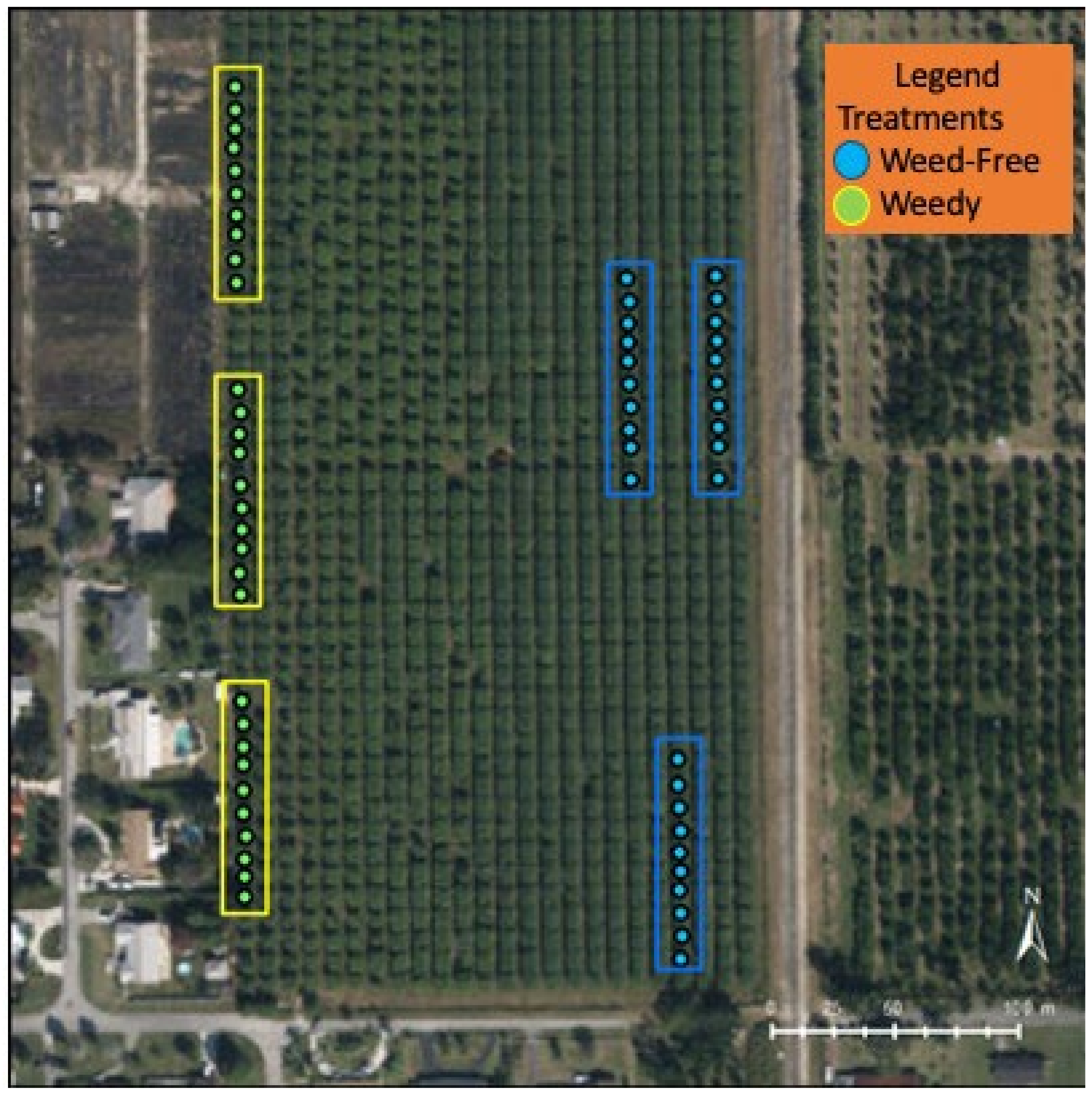
2.1. Site Description
2.2. Farm Maintenance
2.3. Field Data Collection
2.4. Statistical Analyses
2.5. Fruit Yield
3. Results
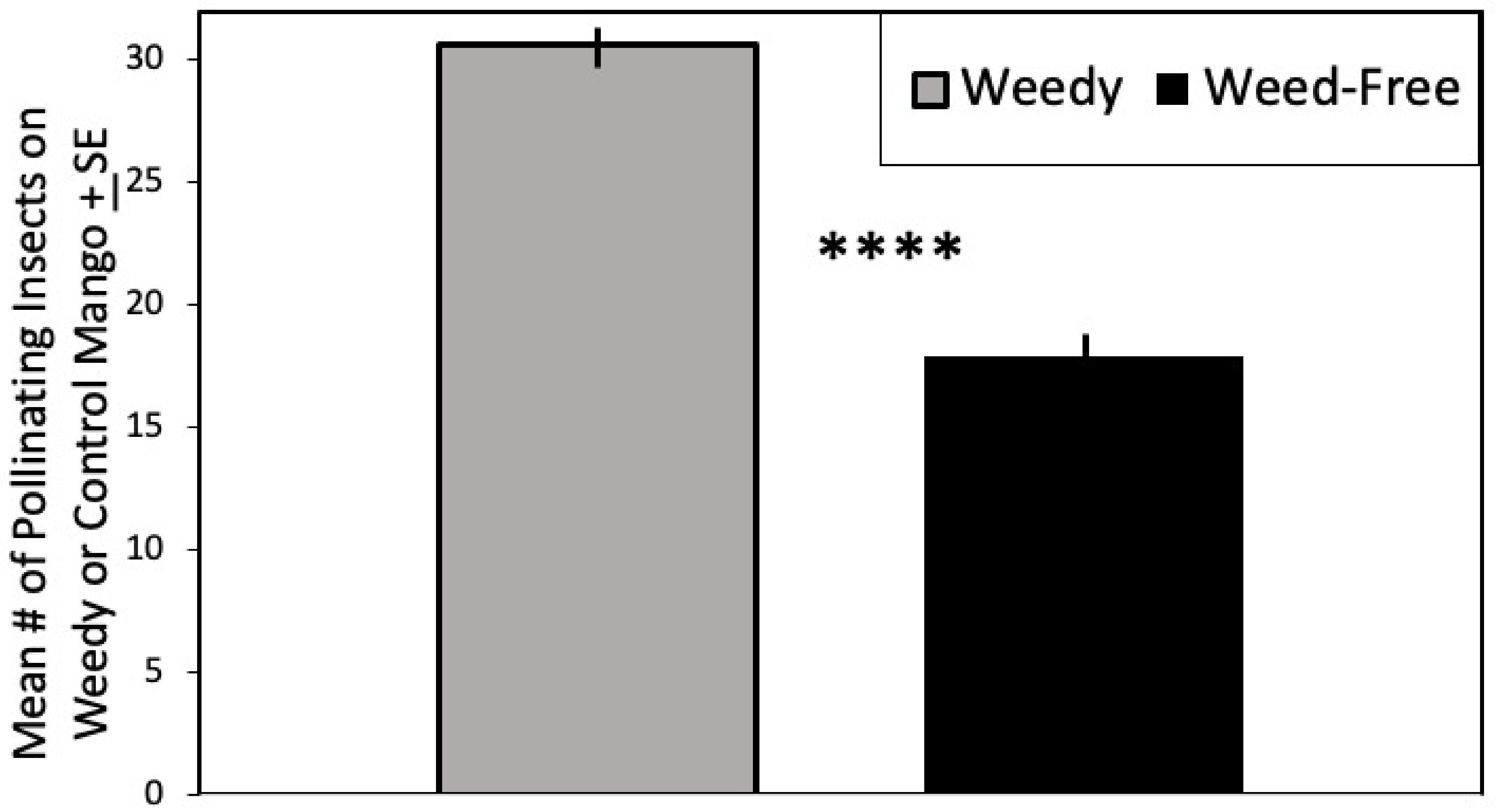
3.1. Flower Visitors of Mango
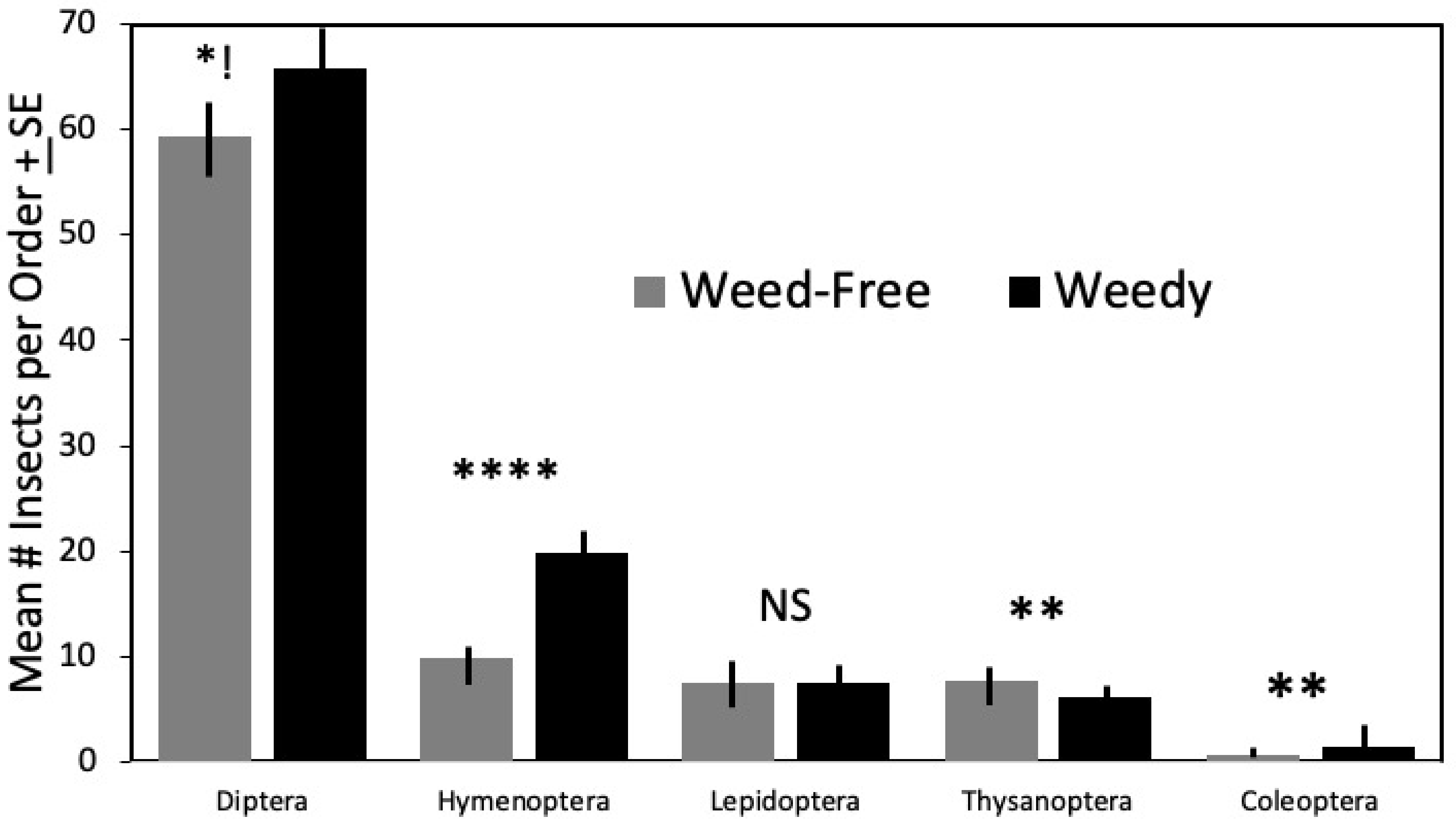
3.2. Families
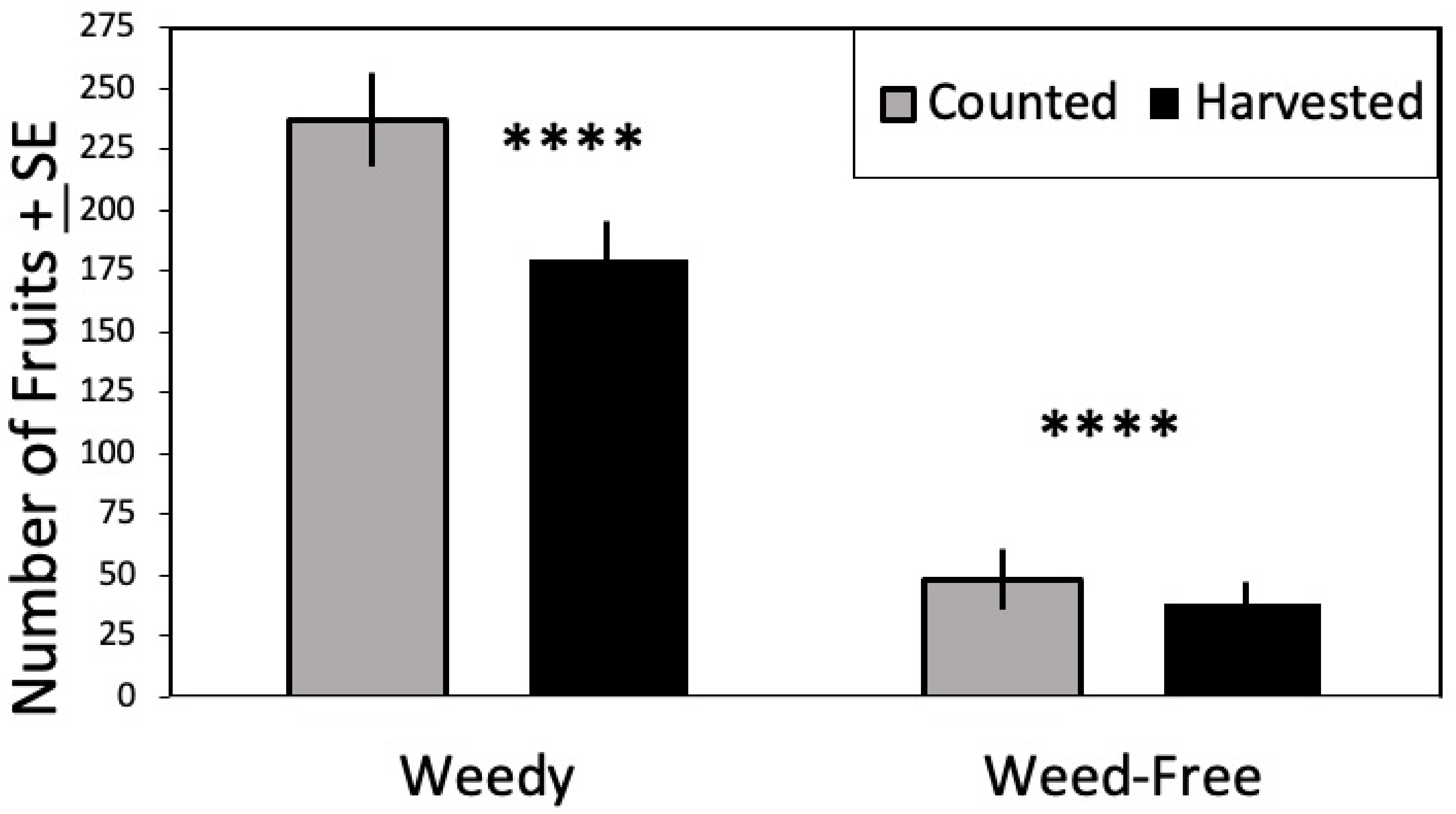
3.3. Fruit Yield
3.4. Weed Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blaauw, B.R.; Isaacs, R. Larger patches of diverse floral resources increase insect pollinator density, diversity, and their pollination of native wildflowers. Basic Appl. Ecol. 2014, 15, 701–711. [Google Scholar] [CrossRef]
- Redhead, J.W.; Powney, G.D.; Woodcock, B.A.; Pywell, R.F. Effects of future agricultural change scenarios on beneficial insects. J. Environ. Manag. 2020, 265, 110550. [Google Scholar] [CrossRef] [PubMed]
- Doré, T.; Makowski, D.; Malézieux, E.; Munier-Jolain, N.; Tchamitchian, M.; Tittonell, P. Facing up to the paradigm of ecological intensification in agronomy: Revisiting methods, concepts and knowledge. Eur. J. Agron. 2011, 34, 197–210. [Google Scholar] [CrossRef]
- Ewel, J.J. Natural systems as models for the design of sustainable systems of land use. Agrofor. Syst. 1999, 45, 1–21. [Google Scholar] [CrossRef]
- Kovács-Hostyánszki, A.; Espíndola, A.; Vanbergen, A.J.; Settele, J.; Kremen, C.; Dicks, L.V.; Irwin, R. Ecological intensification to mitigate impacts of conventional intensive land use on pollinators and pollination. Ecol. Lett. 2017, 20, 673–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleijn, D.; Bommarco, R.; Fijen, T.P.M.; Garibaldi, L.A.; Potts, S.G.; van der Putten, W.H. Ecological Intensification: Bridging the Gap between Science and Practice. Trends Ecol. Evol. 2019, 34, 154–166. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef]
- Altieri, M.A.; Nicholls, C.I. Biodiversity and Pest Management in Agroecosystems; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Hogg, B.N.; Bugg, R.L.; Daane, K.M. Attractiveness of common insectary and harvestable floral resources to beneficial insects. Biol. Control. 2011, 56, 76–84. [Google Scholar] [CrossRef]
- Colley, M.R.; Luna, J.M. Relative Attractiveness of Potential Beneficial Insectary Plants to Aphidophagous Hoverflies (Diptera: Syrphidae). Environ. Èntomol. 2000, 29, 1054–1059. [Google Scholar] [CrossRef]
- Norris, R.F.; Kogan, M. Interactions between weeds, arthropod pests, and their natural enemies in managed ecosystems. Weed Sci. 2000, 48, 94–158. [Google Scholar] [CrossRef]
- Kremen, C.; Williams, N.M.; Thorp, R.W. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. USA 2002, 99, 16812–16816. [Google Scholar] [CrossRef] [Green Version]
- Kleiman, B.; Koptur, S.; Jayachandran, K. Beneficial Interactions of Weeds and Pollinators to Improve Crop Production. J. Res. Weed Sci. 2021, 4, 151–164. [Google Scholar]
- Melin, A.; Rouget, M.; Colville, J.F.; Midgley, J.J.; Donaldson, J.S. Assessing the role of dispersed floral resources for managed bees in providing supporting ecosystem services for crop pollination. PeerJ 2018, 6, e5654. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.A.; Anderson, W.W. Competition for Pollinators between Simultaneously Flowering Species. Am. Nat. 1970, 104, 455–467. [Google Scholar] [CrossRef]
- Ricketts, T.H.; Regetz, J.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Bogdanski, A.; Gemmill-Herren, B.; Greenleaf, S.S.; Klein, A.M.; Mayfield, M.M.; et al. Landscape effects on crop pollination services: Are there general patterns? Ecol. Lett. 2008, 11, 499–515. [Google Scholar] [CrossRef]
- Priess, J.A.; Mimler, M.; Klein, A.-M.; Schwarze, S.; Tscharntke, T.; Steffan-Dewenter, I. Linking deforestation scenarios to pollination services and economic returns in coffee agroforestry systems. Ecol. Appl. 2007, 17, 407–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Veldtman, R.; Shenkute, A.G.; Tesfay, G.B.; Pirk, C.W.W.; Donaldson, J.S.; Nicolson, S.W. Natural and within-farmland biodiversity enhances crop productivity. Ecol. Lett. 2011, 14, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef] [PubMed]
- Dainese, M.; Martin, E.A.; Aizen, M.A.; Albrecht, M.; Bartomeus, I.; Bommarco, R.; Carvalheiro, L.G.; Chaplin-Kramer, R.; Gagic, V.; Garibaldi, L.A.; et al. A global synthesis reveals biodiversity-mediated benefits for crop production. Sci. Adv. 2019, 5, eaax0121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Klink, R.; Bowler, D.E.; Gongalsky, K.B.; Swengel, A.B.; Gentile, A.; Chase, J.M. Meta-analysis reveals declines in terrestrial but increases in freshwater insect abundances. Science 2020, 368, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Steffan-Dewenter, I.; Potts, S.G.; Packer, L. Pollinator diversity and crop pollination services are at risk. Trends Ecol. Evol. 2005, 20, 651–652. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, C.I.; Altieri, M.A. Plant biodiversity enhances bees and other insect pollinators in agroecosystems. A review. Agron. Sustain. Dev. 2013, 33, 257–274. [Google Scholar] [CrossRef] [Green Version]
- Vogel, G. Where have all the insects gone? Science 2017, 356, 576–579. [Google Scholar] [CrossRef]
- Morrison, J.; Izquierdo, J.; Plaza, E.H.; González-Andújar, J.L. The role of field margins in supporting wild bees in Mediterranean cereal agroecosystems: Which biotic and abiotic factors are important? Agric. Ecosyst. Environ. 2017, 247, 216–224. [Google Scholar] [CrossRef]
- Requier, F.; Odoux, J.-F.; Tamic, T.; Moreau, N.; Henry, M.; Decourtye, A.; Bretagnolle, V. Honey bee diet in intensive farmland habitats reveals an unexpectedly high flower richness and a major role of weeds. Ecol. Appl. 2015, 25, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Ghazoul, J. Floral diversity and the facilitation of pollination. J. Ecol. 2006, 94, 295–304. [Google Scholar] [CrossRef]
- Koptur, S.; Barrios, B. Are flowering palms “Pollinator Hogs”? A field experiment in pine rocklands of southern Florida. Nat. Areas J. 2020, 40, 142–149. [Google Scholar] [CrossRef]
- Thomson, J.D. Effects of Stand Composition on Insect Visitation in Two-Species Mixtures of Hieracium. Am. Midl. Nat. 1978, 100, 431–440. [Google Scholar] [CrossRef]
- Gilpin, A.-M.; Denham, A.J.; Ayre, D.J. Do mass flowering agricultural species affect the pollination of Australian native plants through localised depletion of pollinators or pollinator spillover effects? Agric. Ecosyst. Environ. 2019, 277, 83–94. [Google Scholar] [CrossRef]
- Winfree, R.; Williams, N.M.; Gaines, H.; Ascher, J.S.; Kremen, C. Wild bee pollinators provide the majority of crop visitation across land-use gradients in New Jersey and Pennsylvania, USA. J. Appl. Ecol. 2008, 45, 793–802. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Seymour, C.L.; Nicolson, S.W.; Veldtman, R.; Clough, Y. Creating patches of native flowers facilitates crop pollination in large agricultural fields: Mango as a case study. J. Appl. Ecol. 2012, 49, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Bretagnolle, V.; Gaba, S. Weeds for bees? A review. Agron. Sustain. Dev. 2015, 35, 891–909. [Google Scholar] [CrossRef] [Green Version]
- Lowenstein, D.M.; Matteson, K.C.; Minor, E.S. Evaluating the dependence of urban pollinators on ornamental, non-native, and ‘weedy’ floral resources. Urban Ecosyst. 2019, 22, 293–302. [Google Scholar] [CrossRef]
- Kearns, C.; Inouye, D. Pollinators, Flowering Plants, and Conservation Biology. Bioscience 1997, 47, 297–307. [Google Scholar] [CrossRef] [Green Version]
- Willmer, P. Pollination and Floral Ecology; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Dag, A.; Gazit, S. Mango pollinators in Israel. J. Appl. Hortic. 2000, 2, 39–43. [Google Scholar] [CrossRef]
- Heard, T.A. The role of stingless bees in crop pollination. Annu. Rev. Èntomol. 1999, 44, 183–206. [Google Scholar] [CrossRef]
- Kumar, S.; Joshi, P.C.; Nath, P.; Singh, V.K.; Mansotra, D.K. Role of Insects in Pollination of Mango Trees. Int. Res. J. Biol. Sci. 2016, 5, 64. [Google Scholar]
- Kevan, P.G. Pollinators as bioindicators of the state of the environment: Species, activity and diversity. Agric. Ecosyst. Environ. 1999, 74, 373–393. [Google Scholar] [CrossRef]
- Greenleaf, S.S.; Kremen, C. Wild bee species increase tomato production and respond differently to surrounding land use in Northern California. Biol. Conserv. 2006, 133, 81–87. [Google Scholar] [CrossRef]
- Breeze, T.D.; Bailey, A.P.; Balcombe, K.G.; Potts, S.G. Pollination services in the UK: How important are honeybees? Agric. Ecosyst. Environ. 2011, 142, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Free, J.B.; Williams, I.H. Insect pollination of Anacardium occidentale L., Mangifera indica L., Blighia sapida Koenig and Persea americana Mill. Trop. Agric. 1976, 53, 125–139. [Google Scholar]
- Carvalheiro, L.G.; Seymour, C.L.; Veldtman, R.; Nicolson, S.W. Pollination services decline with distance from natural habitat even in biodiversity-rich areas. J. Appl. Ecol. 2010, 47, 810–820. [Google Scholar] [CrossRef]
- Allsopp, M.H.; De Lange, W.J.; Veldtman, R. Valuing Insect Pollination Services with Cost of Replacement. PLoS ONE 2008, 3, e3128. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, R.; Gupta, D.; Chandel, J.S.; Sharma, N.K. Relative abundance of insect visitors on flowers of major subtropical fruits in Himachal Pradesh and their effect on fruit set. Indian J. Agric. Sci. 1995, 65, 907–912. [Google Scholar]
- Evenhuis, N.L.; Pape, T.; Pont, A.C.; Thompson, F.C. Biosystematic Database of World Diptera, Version 10.5; The Catalogue of Life Partnership: Leiden, The Netherlands, 2008. [Google Scholar]
- Ssymank, A. Vegetation and flower-visiting insects in cultivated landscapes. Bonn Bad Godesb. 2001, 64, 513. [Google Scholar]
- Muhammad, F.; Ibrahim, M.; Pervez, M.A. Effect of fungicides on mango malformation. Pak. J. Biol. Sci. 1999, 2, 772–773. [Google Scholar]
- Florida Exotic Pest Plant Council. List of Invasive Plant Species. Available online: www.fleppc.org (accessed on 12 December 2021).
- Lipan, L.; Carbonell-Pedro, A.; Rodríguez, B.C.; Durán-Zuazo, V.H.; Tarifa, D.F.; García-Tejero, I.F.; Ruiz, B.G.; Tavira, S.C.; Muelas, R.; Sendra, E.; et al. Can Sustained Deficit Irrigation Save Water and Meet the Quality Characteristics of Mango? Agriculture 2021, 11, 448. [Google Scholar] [CrossRef]
- Peña, J.E.; Sharp, J.L.; Wysoki, M. Tropical Fruit Pests and Pollinators Biology, Economic Importance, Natural Enemies, and Control; CABI Pub: Wallingford, UK, 2002. [Google Scholar]
- AliAkbarpour, H.; Rawi, C.S.M. The Species Composition of Thrips (Insecta: Thysanoptera) Inhabiting Mango Orchards in Pulau Pinang, Malaysia. Trop. Life Sci. Res. 2012, 23, 45–61. [Google Scholar]
- Nault, B.A.; Speese, J., III; Jolly, D.; Groves, R.L. Seasonal patterns of adult thrips dispersal and implications for management in eastern Virginia tomato fields. Crop. Prot. 2003, 22, 505–512. [Google Scholar] [CrossRef]
- Ananthakrishnan, T.N. The role of thrips in pollination. Curr. Sci. 1993, 65, 262–264. [Google Scholar]
- Retana-Salazar, A.; Rodríguez-Arrieta, A. Potential species of thrips (Insecta: Thysanoptera: Thripidae) in the pollination of mango (Mangifera indica L.) in Costa Rica. Cadiz J. Entomol. 2015, 6, 103–112. [Google Scholar]
- Jiron, L.F.; Hedström, I. Occurrence of Fruit Flies of the Genera Anastrepha and Ceratitis (Diptera: Tephritidae), and Their Host Plant Availability in Costa Rica. Fla. Èntomol. 1988, 71, 62–73. [Google Scholar] [CrossRef]
- Euphoria Sepulcralis (Fabricius) (Insecta: Coleoptera: Scarabaeidae). Available online: https://entnemdept.ufl.edu/creatures/orn/beetles/euphoria_sepulcralis.htm (accessed on 12 December 2021).
- Suhs, R.B.; Somavilla, A.; Köhler, A.; Putzke, J. Vespids (Hymenoptera, Vespidae) pollen vectors of Schinus terebinthifolius Raddi (Anacardiaceae), Santa Cruz do Sul, RS, Brazil. Braz. J. Biosci. 2009, 7, 138–143. [Google Scholar]
- Anderson, D.L.; Sedgley, M.; Short, J.R.T.; Allwood, A.J. Insect pollination of mango in northern Australia. Aust. J. Agric. Res. 1982, 33, 541–548. [Google Scholar] [CrossRef]
- Salas, A. Effects of Host-Plant Density on Herbivores and Their Parasitoids: A Field Experiment with a Native Perennial Legume. Master’s Thesis, Florida International University, Miami, FL, USA, 2016. [Google Scholar]
- Cusser, S.; Pechal, J.L.; Haddad, N.M. Carrion increases pollination service across an urban gradient. Urban Ecosyst. 2021, 24, 243–250. [Google Scholar] [CrossRef]
- Saeed, S.; Naqqash, M.N.; Jaleel, W.; Saeed, Q.; Ghouri, F. The effect of blow flies (Diptera: Calliphoridae) on the size and weight of mangos (Mangifera indica L.). PeerJ 2016, 4, e2076. [Google Scholar] [CrossRef] [Green Version]
- Diehl, E.; Wolters, V.; Birkhofer, K. Arable weeds in organically managed wheat fields foster carabid beetles by resource- and structure-mediated effects. Arthropod Plant Interact. 2012, 6, 75–82. [Google Scholar] [CrossRef]
- Kleiman, B.; Primoli, A.; Koptur, S.; Jayachandran, K. Weeds, pollinators, and parasitoids—Using weeds for insect manipulation in agriculture. J. Res. Weed Sci. 2020, 3, 382–390. [Google Scholar]
- Gibson, R.H.; Nelson, I.L.; Hopkins, G.W.; Hamlett, B.J.; Memmott, J. Pollinator webs, plant communities and the conservation of rare plants: Arable weeds as a case study. J. Appl. Ecol. 2006, 43, 246–257. [Google Scholar] [CrossRef]
- Blaix, C.; Moonen, A.C.; Dostatny, D.F.; Izquierdo, J.; Le Corff, J.; Morrison, J.; Von Redwitz, C.; Schumacher, M.; Westerman, P.R.; Rew, L. Quantification of regulating ecosystem services provided by weeds in annual cropping systems using a systematic map approach. Weed Res. 2018, 58, 151–164. [Google Scholar] [CrossRef]
- Thomas, S.R.; Goulson, D.; Holland, J.M. Resource provision for farmland gamebirds: The value of beetle banks. Ann. Appl. Biol. 2001, 139, 111–118. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The role of weeds in supporting biological diversity within crop fields. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Franke, A.C.; Lotz, L.A.P.; VAN DER Burg, W.J.; VAN Overbeek, L. The role of arable weed seeds for agroecosystem functioning. Weed Res. 2009, 49, 131–141. [Google Scholar] [CrossRef]
- Storkey, J.; Holland, J.M.; Henderson, I.G.; Lutman, P.J.W.; Orson, J.; Baverstock, J.; Pell, J.K. Balancing Food Production and Biodiversity Conservation in Arable Landscapes: Lessons from the Farm4Bio Experiment. Outlooks Pest Manag. 2014, 25, 252–256. [Google Scholar] [CrossRef]
- Altieri, M.A.; Nicholls, C.I.; Henao, A.; Lana, M.A. Agroecology and the design of climate change-resilient farming systems. Agron. Sustain. Dev. 2015, 35, 869–890. [Google Scholar] [CrossRef] [Green Version]
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Dhaliwal, G.S.; Jindal, V.; Mohindru, B. Crop losses to insect pests: Global and Indian scenario. Indian J. Entomol. 2015, 77, 165–168. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.D.; McRoberts, N.; Nelson, A.D. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Capinera, J.L. Relationships between insect pests and weeds: An evolutionary perspective. Weed Sci. 2005, 53, 892–901. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [Green Version]
- Araj, S.-E.; Shields, M.W.; Wratten, S.D. Weed floral resources and commonly used insectary plants to increase the efficacy of a whitefly parasitoid. BioControl 2019, 64, 553–561. [Google Scholar] [CrossRef]
- Provost, C.; Pedneault, K. The organic vineyard as a balanced ecosystem: Improved organic grape management and impacts on wine quality. Sci. Hortic. 2016, 208, 43–56. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Mean | Std Deviation | N | Std Error | F | Df | p Value |
|---|---|---|---|---|---|---|---|
| Pollinators | 31.109 | 1, 57 | <0.0001 | ||||
| Weedy | 30.47 | 1.93 | 30 | 0.35 | |||
| Weed-Free | 18.10 | 3.24 | 30 | 0.59 | |||
| Feeding in Flower | 45.934 | 1, 56 | <0.0001 | ||||
| Weedy | 25.90 | 8.93 | 30 | 1.62 | |||
| Weed-Free | 10.62 | 8.46 | 29 | 1.57 | |||
| Harvested Fruit | 181.317 | 1, 57 | <0.0001 | ||||
| Weedy | 179.37 | 65.34 | 30 | 11.88 | |||
| Weed-Free | 37.67 | 14.63 | 30 | 2.66 | |||
| Counted Fruit | 89.344 | 1, 57 | <0.0001 | ||||
| Weedy | 236.14 | 99.84 | 30 | 18.15 | |||
| Weed-Free | 47.52 | 37.96 | 30 | 6.90 |
| ORDER | Mean | Std Dev | N | Std Error | F | Df | p |
|---|---|---|---|---|---|---|---|
| DIPTERA | 3.792 | 1, 56 | 0.057 | ||||
| Weedy | 65.77 | 10.59 | 30 | 1.93 | |||
| Weed-Free | 59.34 | 14.94 | 29 | 2.77 | |||
| HYMENOPTERA | 80.07 | 1, 56 | <0.0001 | ||||
| Weedy | 19.87 | 5.19 | 30 | 0.94 | |||
| Weed-Free | 9.79 | 3.39 | 29 | 0.63 | |||
| LEPIDOPTERA | 0.001 | 1, 56 | 0.970 | ||||
| Weedy | 7.50 | 2.64 | 30 | 0.48 | |||
| Weed-Free | 7.59 | 4.40 | 29 | 0.82 | |||
| COLEOPTERA | 10.588 | 1, 56 | 0.002 | ||||
| Weedy | 2.47 | 1.31 | 30 | 0.24 | |||
| Weed-Free | 0.55 | 0.83 | 29 | 0.15 | |||
| THYSANOPTERA | 8.208 | 1, 56 | 0.006 | ||||
| Weedy | 6.10 | 2.19 | 30 | 0.40 | |||
| Weed-Free | 7.66 | 2.24 | 29 | 0.42 |
| ON MANGO: | Mean | Std Deviation | N | Std Error Mean | t | Df | p Value |
|---|---|---|---|---|---|---|---|
| Apidae | 4.08 | 55 | 0.000 | ||||
| Weedy | 4.17 | 2.12 | 30 | 0.39 | |||
| Weed-free | 2.04 | 1.79 | 27 | 0.34 | |||
| Calliphoridae | 2.03 | 53 | 0.048 | ||||
| Weedy | 4.07 | 2.14 | 28 | 0.41 | |||
| Weed-free | 3.00 | 1.75 | 27 | 0.34 | |||
| Chalcididae | 2.61 | 29 | 0.003 | ||||
| Weedy | 2.10 | 1.12 | 20 | 0.25 | |||
| Weed-free | 1.18 | 0.41 | 11 | 0.12 | |||
| Chironomidae | −2.5 | 58 | 0.016 | ||||
| Weedy | 4.53 | 1.65 | 30 | 0.30 | |||
| Weed-free | 6.00 | 2.75 | 30 | 0.50 | |||
| Drosophilidae | −2.68 | 33 | 0.011 | ||||
| Weedy | 1.28 | 0.58 | 18 | 0.14 | |||
| Weed-free | 2.06 | 1.09 | 17 | 0.26 | |||
| Lycaenidae | 2.18 | 48 | 0.031 | ||||
| Weedy | 2.42 | 1.17 | 26 | 0.23 | |||
| Weed-free | 1.83 | 0.64 | 24 | 0.13 | |||
| Muscidae | 3.50 | 57 | 0.001 | ||||
| Weedy | 8.27 | 3.81 | 30 | 0.70 | |||
| Weed-free | 5.10 | 3.09 | 29 | 0.57 | |||
| Syrphidae | 3.34 | 44 | 0.002 | ||||
| Weedy | 6.78 | 2.93 | 27 | 0.56 | |||
| Weed-free | 4.57 | 1.91 | 30 | 0.35 | |||
| Vespidae | 2.2 | 18 | 0.041 | ||||
| Weedy | 1.94 | 1.39 | 16 | 0.35 | |||
| Weed-free | 1.13 | 0.35 | 8 | 0.13 |
| Family | Species | Common Name | Native/Invasive |
|---|---|---|---|
| Anacardiaceae | Ruellia blechum | Green Shrimp Plant | Non-native Cat II |
| Ruellia ciliatiflora | Hairy Flower Wild Petunia | Non-native | |
| Schinus terebinthifolia | Brazilian Pepper tree | Non-native Cat I | |
| Arecaceae | Not determined | Various indet. palms | Native & non-native |
| Asteraceae | Bidens alba | Spanish Needles | Native |
| Conoclinium coelestinum | Blue Mistflower | Native | |
| Emilia fosbergii | Florida Tasselflower | Non-native | |
| Parthenium hysterophorus | Santa Maria Feverfew | Non-native | |
| Conyza canadensis | Canadian Horseweed | Native | |
| Ageratum houstonianum | Floss Flower | Non-native | |
| Brassicaceae | Lepidium virginicum | Virginia Pepperweed | Native |
| Lepidium densiflorum | Common Pepperweed | Non-native | |
| Burseraceae | Bursera simaruba | Gumbo Limbo | Native |
| Commelinaceae | Commelina diffusa | Climbing Day flower | Non-native |
| Convolvulaceae | Ipomoea hederifolia | Scarlet Morning Glory | Native |
| Ipomoea indica | Blue Morning Glory | Native | |
| Cucurbitaceae | Melothria pendula | Creeping Cucumber | Native |
| Cyperaceae | Cyperus croceus | Baldwin’s Flatsedge | Native |
| Euphorbiaceae | Acalypha ostryifolia | Hophornbeam Copperleaf | Native |
| Acalypha setosa | Copperleaf | Non-native | |
| Euphorbia heterophylla | Mexican Fireplant | Native | |
| Euphorbia hyssopifolia | Hyssop Spurge | Native | |
| Acalypha arvensis | Field Copperleaf | Non-native | |
| Poinsettia cyathophora | Wild Poinsettia | Native | |
| Euphorbia hirta | Asthma Plant | Native | |
| Fabaceae | Rhynchosia minima | Least Snout-Bean | Native |
| Crotalaria incana | Shake-shake | Non-native | |
| Leucaena leucocephala | White Lead tree | Non-native | |
| Desmanthus virgatus | Wild Tantan | Non-native | |
| Desmodium incanum | Creeping Beggarweed | Non-native | |
| Macroptilium lathyroides | Phasey Bean | Non-native | |
| Indigofera spicata | Creeping Indigo | Non-native | |
| Geraniaceae | Geranium carolinianum | Carolina Geranium | Native |
| Lamiaceae | Salvia occidentalis | West Indian Sage | Native |
| Malvaceae | Sida ulmifolia | Common Wire Weed | Native |
| Moraceae | Ficus aurea | Strangler Fig | Native |
| Oleaceae | Jasminum dichotomum | Gold Coast Jasmine | Non-native Cat I |
| Jasminum fluminense | Brazilian Jasmine | Non-native Cat I | |
| Oxalidaceae | Oxalis debilis | Wood Sorrel | Non-native |
| Papaveraceae | Argemone mexicana | Mexican Prickly Poppy | Native |
| Fumaria officinalis | Common Fumitory | Non-native | |
| Phyllanthaceae | Phyllanthus amarus | Carry-me Seed | Non-native |
| Poaceae | Paspalum conjugatum | Hilo grass | Native |
| Urochloa maxima | Guinea grass | Non-native Cat II | |
| Sporobolus jacquemontii | American Rat’s Tail Grass | Non-native Cat I | |
| Digitaria ciliaris | Southern Crabgrass | Native | |
| Melinis repens | Natal grass | Non-native Cat I | |
| Stenotaphrum secundatum | St. Augustine grass | Native | |
| Sorghum halepense | Johnson grass | Non-native | |
| Panicum maximun | Guinea grass | Non-native | |
| Cenchrus sp. | Sandbur | Native | |
| Neyraudia reynaudiana | Burma Reed | Non-native Cat I | |
| Psilotaceae | Psilotum nudum | Whisk Fern | Native |
| Rubiaceae | Spermacoce remota | Woodland False Buttonweed | Native |
| Spermacoce verticillata | shrubby false buttonweed | Non-native Cat II | |
| Richardia brasiliensis | Brazil Pusley | Non-native | |
| Richardia scabra | Florida Pusley | Non-native | |
| Richardia grandiflora | Large-flower Pusley | Non-native Cat II | |
| Sapindaceae | Cardiospermum corindum | Soapberry | Native |
| Solanaceae | Solanum americanum | American Black Nightshade | Native |
| Urticaceae | Pouzolzia zeylanica | Pouzolzia | Non-native |
| Laportea aestuans | West Indian Wood-nettle | Non-native | |
| Pilea microphylla | Artillery plant | Native | |
| Verbenaceae | Lantana camara | Common Lantana | Non-native |
| Vitaceae | Cissus verticillata | Possum Grape Vine | Native |
| Ampelopsis cordata | Heartleaf Peppervine | Native | |
| Parthenocissus quinquefolia | Virginia Creeper | Non-native | |
| Zamiaceae | Zamia furfuracea | Cardboard Palm | Non-native |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kleiman, B.M.; Koptur, S.; Jayachandran, K. Weeds Enhance Pollinator Diversity and Fruit Yield in Mango. Insects 2021, 12, 1114. https://doi.org/10.3390/insects12121114
Kleiman BM, Koptur S, Jayachandran K. Weeds Enhance Pollinator Diversity and Fruit Yield in Mango. Insects. 2021; 12(12):1114. https://doi.org/10.3390/insects12121114
Chicago/Turabian StyleKleiman, Blaire M., Suzanne Koptur, and Krishnaswamy Jayachandran. 2021. "Weeds Enhance Pollinator Diversity and Fruit Yield in Mango" Insects 12, no. 12: 1114. https://doi.org/10.3390/insects12121114