
Flight and Reproduction Variations of Rice Leaf Roller, Cnaphalocrocis medinalis in Response to Different Rearing Temperatures


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Temperature Treatments
2.3. Larval Development of C. medinalis
2.4. Reproductive Parameter Determination
2.5. Flight Treatment and Capacity Assessments
2.6. Data Analysis
3. Results
3.1. Development of C. medinalis Exposed to Different Rearing Temperatures in the Immature Stage
3.2. Reproductive Performance of C. medinalis Adults from Different Rearing Temperatures in the Immature Stage (the Control Moths without Tethered Flight)
3.3. Flight Capacity of C. medinalis Emergenced from Different Rearing Temperatures in the Immature Stage
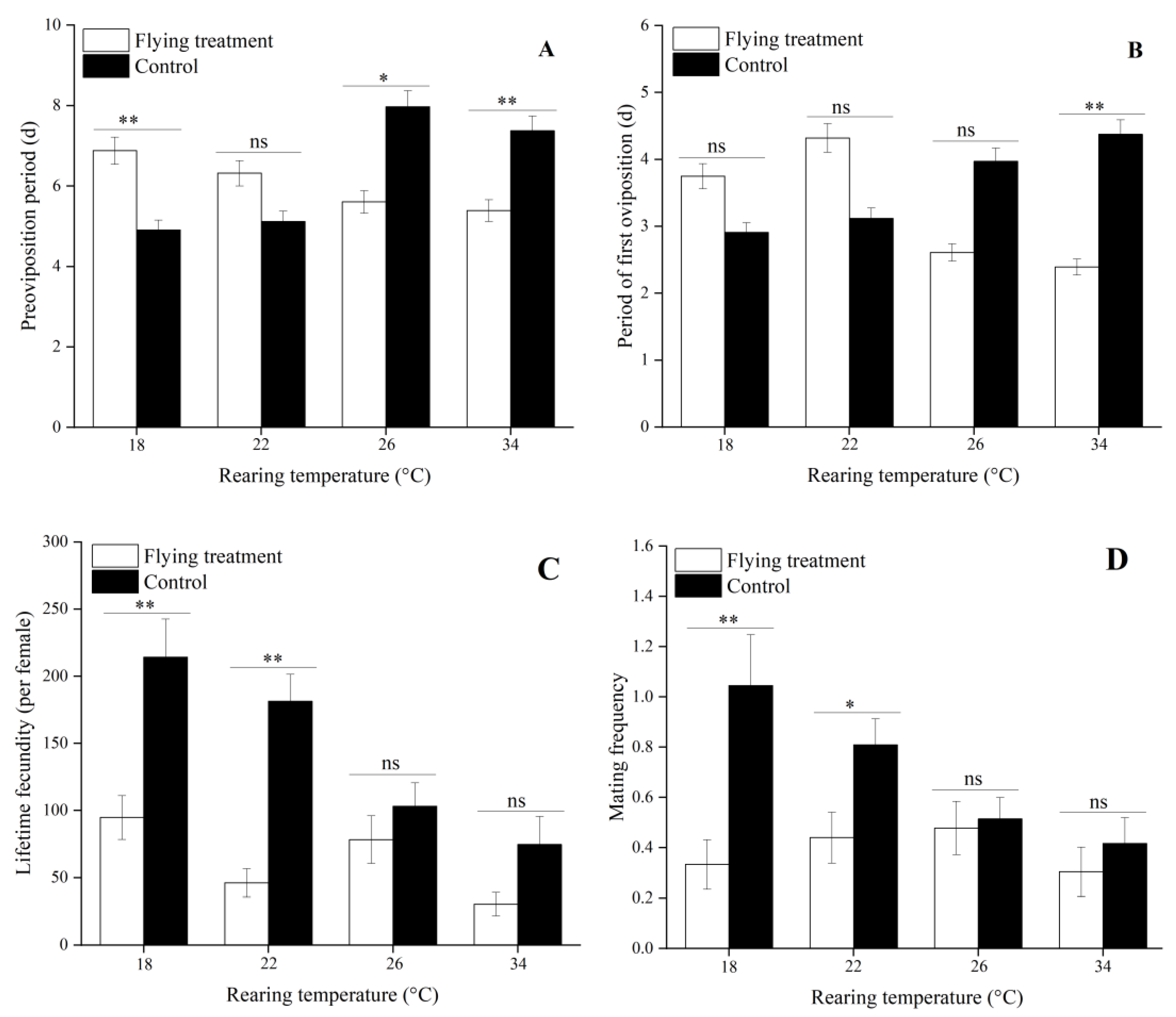
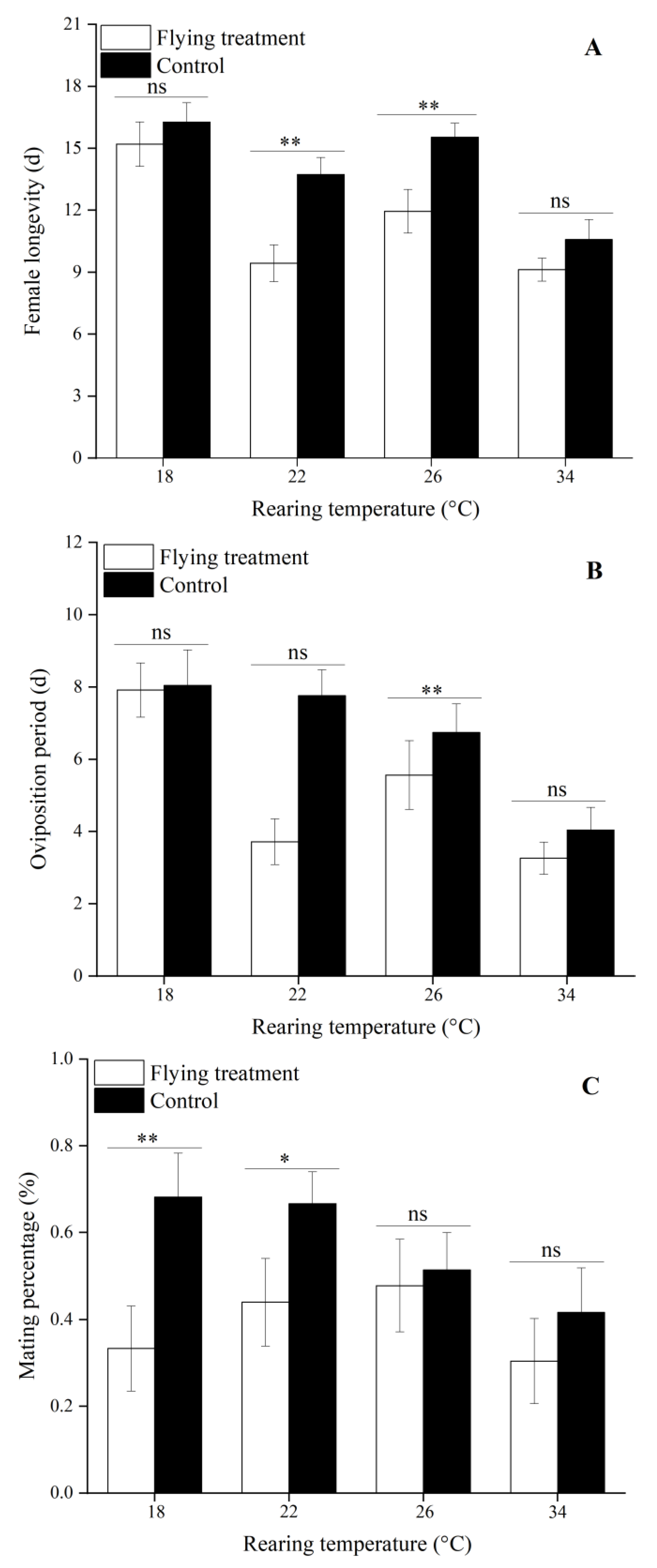
3.4. Influence of Flight on Reproduction Treated by Different Immature Stage Rearing Temperatures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, C.S.; Ma, G.; Pincebourde, S. Survive a Warming Climate: Insect Responses to Extreme High Temperatures. Annu. Rev. Entomol. 2020, 66, 163–184. [Google Scholar] [CrossRef]
- Bale, J.S. Insects and low temperatures: From molecular biology to distributions and abundance. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2002, 357, 849–862. [Google Scholar] [CrossRef] [Green Version]
- Angilletta, M.J., Jr. Thermal Adaptation: A Theoretical and Empirical Synthesis; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Fischer, K.; Kolzow, N.; Holtje, H.; Karl, I. Assay conditions in laboratory experiments: Is the use of constant rather than fluctuating temperatures justified when investigating temperature-induced plasticity? Oecologia 2011, 166, 23–33. [Google Scholar] [CrossRef]
- Piyaphongkul, J.; Pritchard, J.; Bale, J. Heat stress impedes development and lowers fecundity of the brown planthopper Nilaparvata lugens (Stl). PLoS ONE 2012, 7, e47413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pineda, A.; Dicke, M.; Pieterse, C.M.J.; Pozo, M.J. Beneficial microbes in a changing environment: Are they always helping plants to deal with insects? Funct. Ecol. 2013, 27, 574–586. [Google Scholar] [CrossRef] [Green Version]
- Chown, S.L.; Gaston, K.J. Body size variation in insects: A macro ecological perspective. Biol. Rev. 2010, 85, 139–169. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Williams, C.M.; Terblanche, J.S. Variation in thermal performance among insect populations. Physiol. Biochem. Zool. 2012, 85, 594–606. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.X.; Wang, K.; Sappington, T.W.; Luo, L.Z.; Jiang, X.F. Response of reproductive traits and longevity of beet webworm to temperature, and implications for migration. J. Insect Sci. 2015, 15, 154. [Google Scholar] [CrossRef] [Green Version]
- Mironidis, G.K. Development, survivorship and reproduction of Helicoverpa armigera (Lepidoptera: Noctuidae) under fluctuating temperatures. Bull. Entomol. Res. 2014, 104, 751–764. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Higgins, J.K.; Augustine, K.E. Fluctuating temperatures and ectotherm growth: Distinguishing non-linear and time-dependent effects. J. Exp. Biol. 2015, 218, 2218–2225. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.Y.; Liu, Y.Q.; Zhang, L.; Cheng, Y.X.; Luo, L.Z.; Jiang, X.F. Effects of temperatures on the development and reproduction of the armyworm, Mythimna roseilinea: Analysis using an age-stage, two-sex life table. J. Integr. Agric. 2018, 7, 1506–1515. [Google Scholar] [CrossRef]
- Zhou, Z.; Guo, J.; Chen, H.; Wan, F. Effects of temperature on survival, development, longevity, and fecundity of Ophraella communa (Coleoptera: Chrysomelidae), a potential biological control agent against Ambrosia artemisiifolia (Asterales: Asteraceae). Environ. Entomol. 2010, 39, 1021–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regniere, J.; Powell, J.; Bentz, B.; Nealis, V. Effects of temperature on development, survival and reproduction of insects: Experimental design, data analysis and modeling. J. Insect Physiol. 2012, 58, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.Y.; Zhang, L.; Liu, Y.Q.; Sappington, T.W.; Cheng, Y.X.; Luo, L.Z.; Jiang, X.F. Population projection and development of the Mythimna loreyi as affected by temperature: Application of an age-stage, two-sex life table. J. Econ. Entomol. 2017, 110, 1583–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.F.; Luo, L.Z.; Hu, Y. Influence of rearing temperature on flight and reproductive capacity of adult oriental armyworm, Mythimna separata (Walker). Acta Entomol. Sin. 2000, 20, 288–292. [Google Scholar]
- Mourier, H.; Poulsen, K.P. Control of insects and mites in grain using a high temperature/short time (HTST) technique. J. Stored. Prod. Res. 2000, 36, 309–318. [Google Scholar] [CrossRef]
- Li, B.L.; Xu, X.L.; Ji, J.Y.; Wu, J.X. Effects of constant and stage-specific-alternating temperature on the survival, development and reproduction of the oriental armyworm, Mythimna separata (Walker) (Lepidoptera: Noctuidae). J. Integr. Agric. 2018, 7, 1545–1555. [Google Scholar] [CrossRef]
- Hill, J.K.; Gatehouse, A.G. Effect of temperature and photoperiod on development and pre-reproductive period of the silver Y moth Autographa gamma (Lepidoptera: Noctuidae). Bull. Entomol. Res. 1992, 82, 335–341. [Google Scholar] [CrossRef]
- Han, E.N.; Gatehouse, A.G. Effect of temperature and photoperiod on the calling behaviour of a migratory insect, the oriental armyworm Mythimna separata. Physiol. Entomol. 1991, 16, 419–427. [Google Scholar] [CrossRef]
- Mirth, C.K.; Saunders, T.E.; Amourda, C. Growing Up in a Changing World: Environmental Regulation of Development in Insects. Annu. Rev. Entomol. 2021, 66, 81–99. [Google Scholar] [CrossRef] [PubMed]
- Stefanescu, C.; Páramo, F.; Åkesson, S.; Alarcón, M.; Anna, Á.; Brereton, T.; Carnicer, J.; Cassar, L.F.; Fox, R.; Heliölä, J. Multi-generational long-distance migration of insects: Studying the painted lady butterfly in the Western Palaearctic. Ecography 2013, 36, 474–486. [Google Scholar] [CrossRef] [Green Version]
- Rankin, M.A.; Burchsted, J.C.A. The cost of migration in insects. Annu. Rev. Entomol. 1992, 37, 533–559. [Google Scholar] [CrossRef]
- Sonia, A.; Rebecca, B.; Barbara, A.H. Animal migration and infectious disease risk. Science 2011, 331, 296–302. [Google Scholar]
- Dingle, H. Migration: The Biology of Life on the Move, 2nd ed.; Oxford University Press: New York, NY, USA, 2014. [Google Scholar]
- Chapman, J.W.; Reynolds, D.R.; Wilson, K. Long-range seasonal migration in insects: Mechanisms, evolutionary drivers and ecological consequences. Ecol. Lett. 2015, 18, 287–302. [Google Scholar] [CrossRef] [Green Version]
- Zera, A.J.; Denno, R.F. Physiology and ecology of dispersal polymorphism in insects. Annu. Rev. Entomol. 1997, 42, 207–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Pan, P.; Sappington, T.W.; Lu, W.X.; Luo, L.Z.; Jiang, X.F. Accelerated and synchronized oviposition induced by flight of young females may intensify larval outbreaks of the rice leaf roller. PLoS ONE 2015, 10, e0121821. [Google Scholar] [CrossRef] [Green Version]
- Guerra, P.A.; Pollack, G.S. Flight behavior attenuates the trade-off between flight capability and reproduction in a wing polymorphic cricket. Biol. Lett. 2009, 5, 229–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wu, K.M.; Kris, A.G.W.; George, E.H. Trade-off between flight and fecundity in the soybean aphid (Hemiptera: Aphididae). J. Econ Entomol. 2009, 102, 133–138. [Google Scholar] [CrossRef]
- Saglam, I.K.; Roff, D.A.; Fairbairn, D.J. Male sand crickets trade-off flight capability for reproductive potential. J. Evolution Biol. 2010, 21, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.G. Migration and Dispersal of Insects by Flight; Methuen: London, UK, 1969. [Google Scholar]
- Bautista, R.C.; Henirichs, E.A.; Rejesus, R.S. Economic injury levels for the rice leaf folder Cnaphalocrocis medinalis (Lepidoptera: Pyralidae): Insect infestation and artificial leaf removal. Environ. Entomol. 1984, 13, 439–443. [Google Scholar] [CrossRef] [Green Version]
- Riley, J.R.; Reynolds, D.R.; Smith, A.D.; Edwards, A.S.; Zhang, X.X.; Cheng, X.N.; Wang, H.K.; Cheng, J.Y.; Zhai, B.P. Observations of the autumn migration of the rice leaf roller Cnaphalocrocis medinalis (Lepidoptera: Pyralidae) and other moths in eastern China. Bull. Entomol. Res. 1995, 85, 397–414. [Google Scholar] [CrossRef]
- Zhang, X.X.; Lu, Z.Q.; Geng, J.G.; Li, G.Z.; Chen, X.L.; Wu, X.W. Studies on the migration of rice leaf roller Cnaphalocrocis medinalis (Guenée). Acta Entomol. Sin. 1980, 2, 130–140. [Google Scholar]
- Li, G.B.; Wang, H.X.; Hu, W.X. Route of the seasonal migration of the oriental armyworm moth in the eastern part of China as indicated by a three-year result of releasing and recapturing of marked moths. Acta Phytopathol. Sin. 1964, 3, 101–110. [Google Scholar]
- Jiang, X.F.; Luo, L.Z.; Zhang, L.; Sappington, T.W.; Hu, Y. Regulation of migration in Mythimna separata (Walker) in China: A review integrating environmental, physiological, hormonal, genetic, and molecular factors. Environ. Entomol. 2011, 40, 516–533. [Google Scholar] [CrossRef]
- Wang, F.; Yang, Y.F.; Lu, M.H.; Luo, S.Y.; Zhai, B.P.; Lim, K.S. Determining the migration duration of rice leaf folder (cnaphalocrocis medinalis (guenée)) moths using a trajectory analytical approach. Sci. Rep. 2017, 7, 39853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.W.; Li, W.P.; Zhang, J.; Liu, X.D.; Zhai, B.P.; Gao, H. Cnaphalocrocis medinalis moths decide to migrate when suffering nutrient shortage on the first day after emergence. Insects 2019, 10, 364. [Google Scholar] [CrossRef] [Green Version]
- Potter, K.A.; Woods, A.; Pincebourde, S. Microclimatic challenges in global change biology. Glob. Chang. Biol. 2013, 19, 2932–2939. [Google Scholar] [CrossRef]
- Klockmann, M.; Günter, F.; Fischer, K. Heat resistance throughout ontogeny: Body size constrains thermal tolerance. Glob. Chang. Biol. 2017, 23, 686–696. [Google Scholar] [CrossRef]
- Chen, B.; Feder, M.E.; Kang, L. Evolution of heat-shock protein expression underlying adaptive responses to environmental stress. Mol. Ecol. 2018, 27, 3040–3054. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.N.; Zhang, W.; Ma, G.; Hoffmann, A.; Ma, C.S. A single hot event stimulates adult performance but reduces egg survival in the oriental fruit moth, Grapholitha molesta. PLoS ONE 2014, 9, e116339. [Google Scholar] [CrossRef]
- Chintalapati, P.; Gururaj, K.; Sailaja, V.; Padmakumari, A.P.; Jhansilakshmi, V.; Prabhakar, M.; Prasad, Y.G. Temperature Thresholds and Thermal Requirements for the Development of the Rice Leaf Folder, Cnaphalocrocis medinalis. J. Insect Sci. 2013, 1, 1–14. [Google Scholar]
- Guo, J.W.; Yang, F.; Li, P.; Liu, X.D.; Zhai, B.P. Female bias in an immigratory population of Cnaphalocrocis medinalis moths based on field surveys and laboratory tests. Sci. Rep. 2019, 9, 18388. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.F.; Luo, L.Z.; Sappington, T.W. Relationship of flight and reproduction in beet armyworm, Spodoptera exigua (Lepidoptera: Noctuidae), a migrant lacking the oogenesis-flight syndrome. J. Insect Physiol. 2010, 56, 1631–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Cheng, L.L.; Chapman, J.W.; Sappington, T.W.; Liu, J.J.; Cheng, Y.X.; Jiang, X.F. Juvenile hormone regulates the shift from migrants to residents in adult oriental armyworm, Mythimna separata. Sci. Rep. 2020, 10, 11626. [Google Scholar] [CrossRef]
- Cheng, Y.X.; Luo, L.Z.; Jiang, X.F.; Sappington, T.W. Synchronized oviposition triggered by migratory flight intensifies larval outbreaks of beet webworm. PLoS ONE 2012, 7, e31562. [Google Scholar]
- Qin, J.Y.; Liu, Y.Q.; Zhang, L.; Cheng, Y.X.; Sappington, T.W.; Jiang, X.F. Effects of moth age and rearing temperature on the flight performance of the loreyi leafworm, Mythimna loreyi (Lepidoptera: Noctuidae), in tethered and free flight. J. Econ. Entomol. 2018, 111, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.F. Regularity of population occurrence and migration in the oriental armyworm, Mythimna separata (Walker). J. Integr. Agric. 2018, 17, 1482–1505. [Google Scholar] [CrossRef]
- Mironidis, G.K.; Savopoulou-Soultani, M. Development, survivorship, and reproduction of Helicoverpa armigera (Lepidoptera: Noctuidae) under constant and alternating temperatures. Environ. Entomol. 2008, 37, 16–28. [Google Scholar] [CrossRef]
- Ghazanfar, M.U.; Hagenbucher, S.; Romeis, J.; Grabenweger, G.; Meissle, M. Fluctuating temperatures influence the susceptibility of pest insects to biological control agents. J. Pest Sci. 2020, 93, 1007–1018. [Google Scholar] [CrossRef]
- Karimi-Malati, A.; Fathipour, Y.; Talebi, A.A.; Bazoubandi, M. Lifetable parameters and survivorship of Spodoptera exigua (Lepidoptera: Noctuidae) at constant temperatures. Environ. Entomol. 2014, 43, 795–803. [Google Scholar] [CrossRef]
- Sibly, R.M.; Atkinson, D. How rearing temperature affects optimal adult size in ectotherms. Func. Ecol. 1994, 8, 486–493. [Google Scholar] [CrossRef]
- Luo, L.Z.; Jiang, X.F.; Li, K.B.; Hu, Y. Influences of flight on reproduction and longevity of the oriental armyworm, Mythimna separata (Walker). Acta Entomol. Sin. 1999, 2, 150–158. [Google Scholar]
- Zhang, L.; Jiang, X.F.; Luo, L.Z. Determination of sensitive stage for switching migrant oriental armyworms into residents. Environ. Entomol. 2008, 37, 1389–1395. [Google Scholar] [CrossRef]
- Nijhout, H.F. Control mechanisms of polyphenic development in insects. Bioscience 1999, 49, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.L.; Luo, L.Z.; Jiang, X.F.; Zhang, L. Effects of larval density on flight potential of the beet webworm, Loxostege sticticalis (Lepidoptera: Pyralidae). Environ. Entomol. 2010, 39, 1579–1585. [Google Scholar] [CrossRef]
- Jiang, X.F.; Luo, L.Z. Comparison of behavioral and physiological characteristics between the emigrant and immigrant populations of the oriental armyworm, Mythimna separata (Walker). Acta Entomol. Sin. 2005, 48, 61–67. [Google Scholar]
- Wang, F.Y.; Zhang, X.X.; Zhai, B.P. Flight and re-migration capacity of the rice leaf folder moth, Cnaphalocrocis medinalis(Guenée) (Lepidoptera: Crambidae). Acta Entomol. Sin. 2010, 53, 1265–1272. [Google Scholar]
- Taylor, L.R. Analysis of the effect of temperature on insects in flight. J. Anim Ecol. 1963, 32, 99–117. [Google Scholar] [CrossRef]
- Stevenson, R.D.; Josephson, R.K. Effects of operating frequency and temperature on mechanical power output from moth flight muscle. J. Exp. Biol. 1990, 149, 61–78. [Google Scholar] [CrossRef]
- Makumbe, L.M.; Moropa, T.P.; Manrakhan, M.; Weldon, C.W. Effect of sex, age and morphological traits on tethered flight of Bactrocera dorsalis (Hendel) (Diptera:Tephritidae) at different temperatures. Physiol. Entomol. 2020, 7, 110–119. [Google Scholar] [CrossRef]
- Goller, F.; Esch, H. Comparative study of chill-coma temperatures and muscle potentials in insect flight muscles. J. Exp. Biol. 1990, 150, 221–231. [Google Scholar] [CrossRef]
- MacMillan, H.A.; Sinclair, B.J. Mechanisms underlying insect chill-coma. J. Insect Physiol. 2011, 57, 12–20. [Google Scholar] [CrossRef]
- Neven, L.G. Physiological responses of insects to heat. Postharvest Biol. Technol. 2000, 21, 103–111. [Google Scholar] [CrossRef]
- Davis, M.A. The flight and migration ecology of the red milkweed beetle (Tetraopes Tetraophthalmus). Ecology 1984, 65, 230–234. [Google Scholar] [CrossRef]
- Martínez, A.S.; Villacide, J.; Fernández Ajó, A.A.S.; Martinson, J.; Corley, J.C. Sirex noctilio flight behavior: Toward improving current monitoring techniques. Entomol. Exp. Appl. 2014, 152, 135–140. [Google Scholar] [CrossRef]
- Cui, J.; Li, S.; Zhao, P.; Zou, F. Flight capacity of adult Culex pipiens pallens (Diptera: Culicidae) in relation to gender and day-age. J. Med. Entomol. 2013, 50, 1055–1058. [Google Scholar] [CrossRef]
- Sappington, T.W.; Showers, W. Reproductive maturity, mating status, and long-duration flight behavior of Agrotis ipsilon and the conceptual misuse of the oo-genesis flight syndrome by entomologists. Environ. Entomol. 1992, 21, 677–688. [Google Scholar]
- Zhang, N.; Xie, X.C.; Cheng, W.N.; Wu, J.X.; Li, Y.P. Effects of high- or low-temperature treatment time on energy material and flight capacity in the adults of the oriental armyworm, Mythimna separata (Walker). J. Plant Prot. 2018, 45, 243–250. [Google Scholar]
- Sun, B.B.; Zhang, L.; Jiang, X.F.; Luo, L.Z. Effects of temperature in adult period on the reproduction behavior of the rice leaf roller, Cnaphalocrocis medinalis (Guenée). Chin. J. Appl. Entomol. 2013, 50, 622–628. [Google Scholar]
- Wu, J.C. Effect of changing photoperiod, temperature and food quality on the migration of rice leaf folder Cnaphalocrocis medinalis (Guenée). Acta Entomol. Sin. 1985, 28, 398–405. [Google Scholar]
- McCalla, K.A.; Keçeci, M.; Milosavljević, I.; Ratkowsky, D.A. The influence of temperature variation on life history parameters and thermal performance curves of tamarixia radiata (hymenoptera: Eulophidae), a parasitoid of the asian citrus psyllid (hemiptera: Liviidae). J. Econ. Entomol. 2019, 112, 1560–1574. [Google Scholar] [CrossRef] [PubMed]
- Milosavljević, I.; McCalla, K.A.; Ratkowsky, D.A.; Hoddle, M.S. Effects of constant and fluctuating temperatures on development rates and longevity of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae). J. Econ. Entomol. 2019, 112, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Milosavljević, I.; McCalla, K.A.; Morgan, D.J.W.; Hoddle, M.S. The effects of constant and fluctuating temperatures on development of Diaphorina citri (Hemiptera: Liviidae), the Asian Citrus Psyllid. J. Econ. Entomol. 2020, 113, 633–645. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Development Parameters | Rearing Temperatures | |||
|---|---|---|---|---|
| 18 °C | 22 °C | 26 °C | 34 °C | |
| Egg-larval period (d) | 53.80 ± 0.39 a | 30.59 ± 0.23 b | 23.76 ± 0.08 c | 23.25 ± 0.14 c |
| Pupal period (d) | 17.83 ± 0.31 a | 8.87 ± 0.19 b | 5.78 ± 0.09 c | 4.99 ± 0.10 d |
| Larval mortality rate (%) | 74.00 ± 10.51 a | 49.35 ± 12.52 a | 46.00 ± 25.14 a | 53.05 ± 18.49 a |
| Pupal mortality rate (%) | 36.45 ± 23.86 a | 25.62 ± 8.71 a | 18.85 ± 3.45 a | 27.70 ± 5.04 a |
| Pupation rate (%) | 35.14 ± 25.47 a | 50.65 ±12.52 a | 54.00 ± 25.14 a | 46.95 ± 18.49 a |
| Eclosion rate (%) | 71.34 ± 8.15 a | 59.94 ± 23.97 a | 76.64 ± 3.98 a | 68.61 ± 6.28 a |
| Period of synchronized eclosion (d) | 9.40 ± 0.48 a | 5.66 ± 0.24 b | 4.08 ± 0.13 c | 4.61 ± 0.17 c |
| Sex ratio (female/total progeny) | 0.55 ± 0.02 a | 0.56 ± 0.06 a | 0.57 ± 0.04 a | 0.52 ± 0.08 a |
| Reproductive Parameters | Rearing Temperatures | |||
|---|---|---|---|---|
| 18 °C | 22 °C | 26 °C | 34 °C | |
| Preoviposition period (d) | 4.91 ± 0.42 b | 5.12 ± 0.26 b | 7.97 ± 0.90 a | 7.38 ± 0.48 a |
| Period of first oviposition (d) | 2.91 ± 0.42 a | 3.12 ± 0.26 a | 3.97 ± 0.90 a | 4.38 ± 0.48 a |
| Lifetime fecundity | 214.23 ± 28.31 a | 181.40 ± 20.13 a | 103.20 ± 17.38 b | 74.62 ± 20.83 b |
| Mating frequency | 1.05 ± 0.20 a | 0.81 ± 0.10 ab | 0.51 ± 0.08 b | 0.42 ± 0.10 b |
| Female longevity (d) | 16.27 ± 0.93 a | 13.74 ± 0.82 a | 15.54 ± 0.68 a | 10.58 ± 0.97 b |
| Oviposition period (d) | 8.05 ± 0.98 a | 7.76 ± 0.72 a | 6.74 ± 0.79 ab | 4.04 ± 0.63 b |
| Mating percentage (%) | 68.18 ± 10.16 a | 66.67 ± 7.36 a | 51.43 ± 8.57 a | 41.67 ± 10.28 a |
| Parameters | Sex | Rearing Temperatures | |||
|---|---|---|---|---|---|
| 18 °C | 22 °C | 26 °C | 34 °C | ||
| Flight duration (h) | Female | 2.34 ± 0.47 a | 3.37 ± 0.36 a | 3.87 ± 0.37 a | 3.25 ± 0.36 a * |
| Male | 3.09 ± 0.55 ab | 3.04 ± 0.34 ab | 3.74 ± 0.34 a | 2.02 ± 0.34 b | |
| Flight distance (km) | Female | 4.42 ± 1.14 a | 7.13 ± 1.09 a | 8.12 ± 1.13 a | 6.30 ± 0.98 a |
| Male | 6.07 ± 1.29 a | 6.34 ± 0.96 a | 7.92 ± 0.97 a | 4.15 ± 0.91 b | |
| Average velocity | Female | 1.54 ± 0.21 a | 2.24 ± 0.1 7 a | 2.24 ± 0.14 a | 2.76 ± 0.18 a |
| (km/h) | Male | 1.75 ± 0.21 a | 1.82 ± 0.15 a | 1.92 ± 0.13 a | 1.67 ± 0.14 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, W.; Jiang, X.; Chen, X.; Cheng, Y.; Xia, J.; Zhang, L. Flight and Reproduction Variations of Rice Leaf Roller, Cnaphalocrocis medinalis in Response to Different Rearing Temperatures. Insects 2021, 12, 1083. https://doi.org/10.3390/insects12121083
Lv W, Jiang X, Chen X, Cheng Y, Xia J, Zhang L. Flight and Reproduction Variations of Rice Leaf Roller, Cnaphalocrocis medinalis in Response to Different Rearing Temperatures. Insects. 2021; 12(12):1083. https://doi.org/10.3390/insects12121083
Chicago/Turabian StyleLv, Weixiang, Xingfu Jiang, Xiujie Chen, Yunxia Cheng, Jixing Xia, and Lei Zhang. 2021. "Flight and Reproduction Variations of Rice Leaf Roller, Cnaphalocrocis medinalis in Response to Different Rearing Temperatures" Insects 12, no. 12: 1083. https://doi.org/10.3390/insects12121083