A Bioclimate-Based Maximum Entropy Model for Comperiella calauanica Barrion, Almarinez and Amalin (Hymenoptera: Encyrtidae) in the Philippines

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Species Presence, Bioclimatic Variables, and Other Data
2.2. Maxent Species Distribution Modeling for C. calauanica
2.3. Analysis and Assessment of the Species Distribution Model
2.4. Validation of the Predicted Distribution of A. rigidus
3. Results
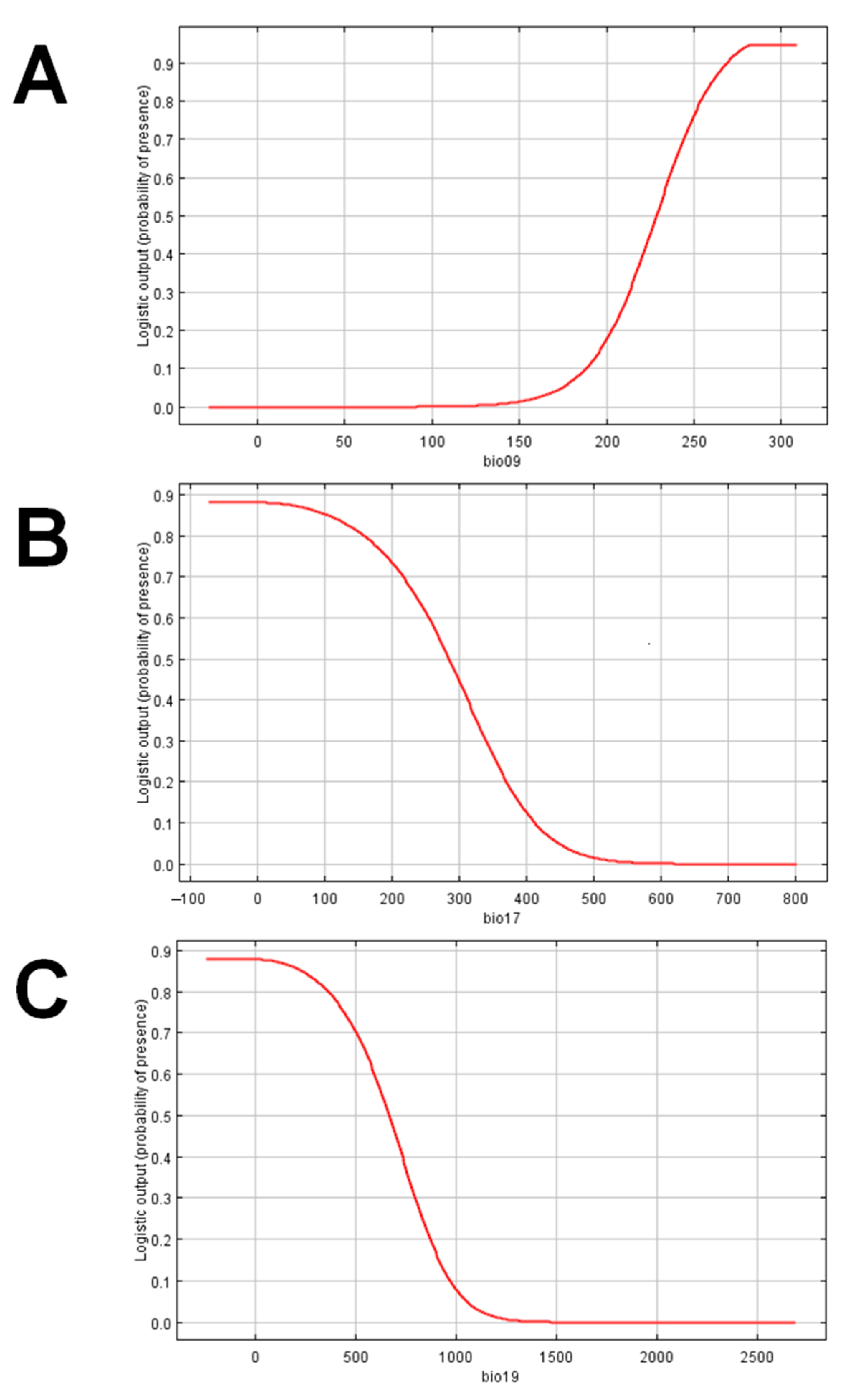
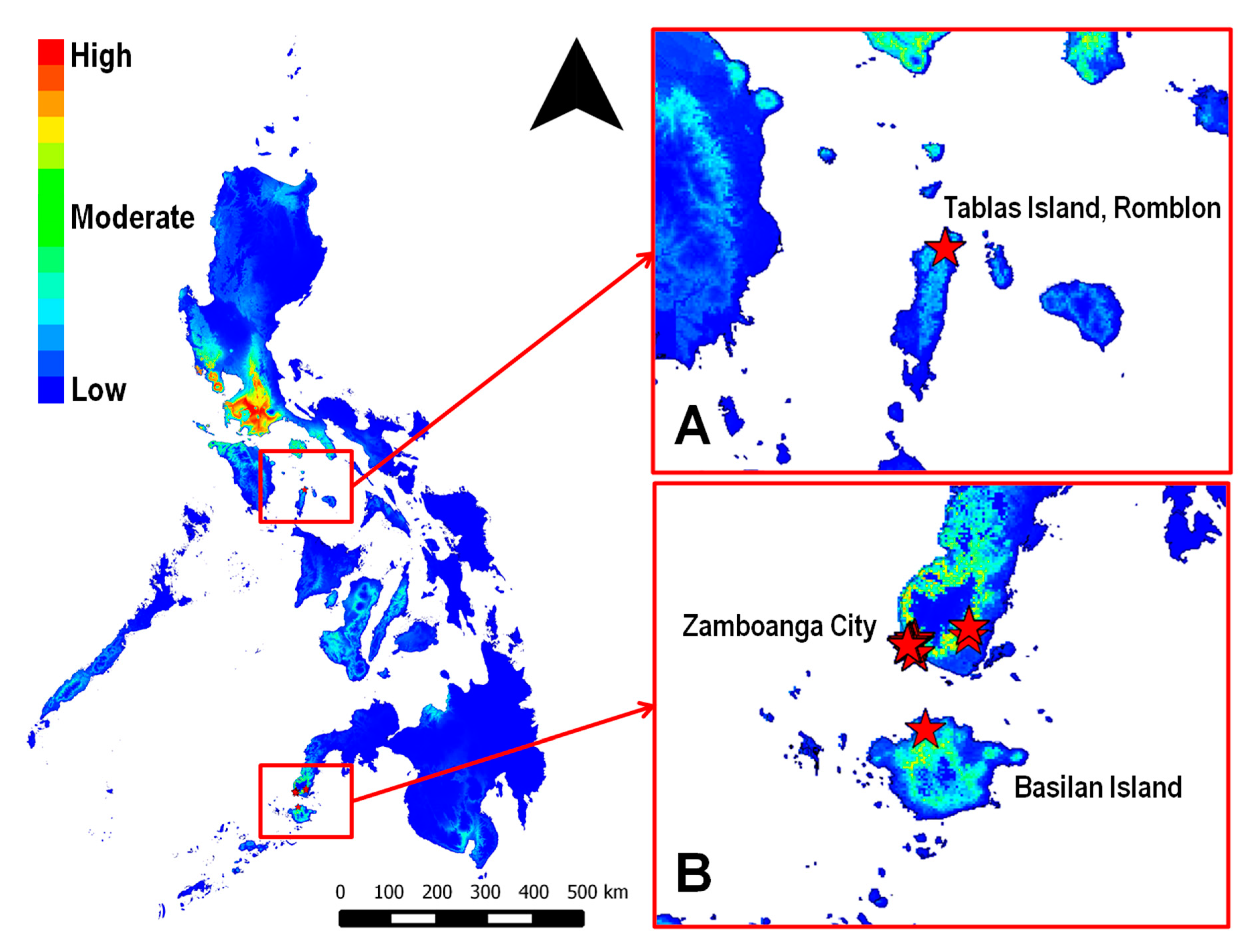
3.1. Maxent Species Distribution Model for C. calauanica
3.2. Analysis and Assessment of the Species Distribution Model
3.3. Validation of the Predicted Distribution of A. rigidus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Bank. Employment in Agriculture (% of Total Employment, Philippines). 2017. Available online: http://data.worldbank.org/indicator/SL.AGR.EMPL.ZS?end=2015&locations=PH&start=2015&view=bar (accessed on 4 March 2019).
- Philippine Statistics Authority. Selected Statistics on Agriculture. 2016. Available online: https://psa.gov.ph/sites/default/files/Selected%20Statistics%20on%20Agriculture%202016.pdf (accessed on 4 March 2019).
- Watson, G.W.; Adalla, C.B.; Shepard, B.M.; Carner, G.R. Aspidiotus rigidus Reyne (Hemiptera: Diaspididae): A devastating pest of coconut in the Philippines. Agric. For. Entomol. 2015, 17, 1–8. [Google Scholar] [CrossRef]
- Almarinez, B.J.M.; Amalin, D.M.; Carandang, J.S.R.; Navasero, M.V.; Navasero, M.M. First Philippine record of the parasitoid, Comperiella sp. (Hymenoptera: Encyrtidae): A potential biological control agent against Aspidiotus rigidus (Hemiptera: Diaspididae). J. Appl. Entomol. 2015, 139, 237–240. [Google Scholar] [CrossRef]
- Barrion, A.T.; Almarinez, B.J.M.; Amalin, D.M.; Carandang, J.S.R. Comperiella calauanica sp. n. (Hymenoptera: Encyrtidae), an endoparasitoid of the invasive coconut scale, Aspidiotus rigidus Reyne (Hemiptera: Diaspididae) on Luzon Island, Philippines. Asia Life Sci. 2016, 25, 1–15. [Google Scholar]
- Palen, D.I.; Almarinez, B.J.M.; Amalin, D.M.; Legaspi, J.C.; David, G. A host-parasitoid model for Aspidiotus rigidus (Hemiptera: Diaspididae) and Comperiella calauanica (Hymenoptera: Encyrtidae). Environ. Entomol. 2019, 48, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Almarinez, B.J.M.; Barrion, A.T.; Navasero, M.V.; Navasero, M.M.; Cayabyab, B.F.; Carandang, J.S.R.; Legaspi, J.C.; Watanabe, K.; Amalin, D.M. Biological Control: A Major Component of the Pest Management Program for the Invasive Coconut Scale Insect, Aspidiotus rigidus Reyne, in the Philippines. Insects 2020, 11, 745. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Schwartz, M.W. Using niche models with climate projections to inform conservation management decisions. Biol. Conserv. 2012, 155, 149–156. [Google Scholar] [CrossRef]
- Smeraldo, S.; Bosso, L.; Fraissinet, M.; Bordignon, L.; Brunelli, M.; Ancilloto, L.; Russo, D. Modelling risks posed by wind turbines and power lines to soaring birds: The black stork (Ciconia nigra) in Italy as a case study. Biodivers. Conserv. 2020, 29, 1959–1976. [Google Scholar] [CrossRef]
- Malahlela, O.E.; Adjorlolo, C.; Olwoch, J.M. Mapping the spatial distribution of Lippia javanica (Burm. f.) Spreng using Sentinel-2 and SRTM-derived topographic data in malaria endemic environment. Ecol. Model. 2019, 392, 147–158. [Google Scholar] [CrossRef]
- Raffini, F.; Bertorelle, G.; Biello, R.; D’Urso, G.; Russo, D.; Bosso, L. From Nucleotides to Satellite Imagery: Approaches to Identify and Manage the Invasive Pathogen Xylella fastidiosa and Its Insect Vectors in Europe. Sustainability 2019, 12, 4508. [Google Scholar] [CrossRef]
- Howse, M.W.F.; Haywood, J.; Lester, P.J. Bioclimatic Modelling Identifies Suitable Habitat for the Establishment of the Invasive European Paper Wasp (Hymenoptera: Vespidae) across the Southern Hemisphere. Insects 2020, 11, 745. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Dudik, M.; Schapire, R.E. A Maximum Entropy Approach to Species Distribution Modeling. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AB, Canada, 4–8 July 2004; Association for Computing Machinery: New York, NY, USA, 2004; pp. 655–662. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Yu, S.; Capinha, C.; Weterings, R.; Gao, T. Using species distribution model to predict the impact of climate change on the potential distribution of Japanese whiting Sillago japonica. Ecol. Indic. 2019, 104, 333–340. [Google Scholar] [CrossRef]
- Schowalter, T.D. Insect Ecology: An Ecosystem Approach, 3rd ed.; Academic Press: London, UK, 2011; pp. 163–165. [Google Scholar]
- Gormley, A.M.; Forsyth, D.M.; Griffioen, P.; Lindeman, M.; Ramsey, D.S.L.; Scroggie, M.P.; Woodford, L. Using presence-only and presence-absence data to estimate the current and potential distributions of established invasive species. J. Appl. Ecol. 2011, 48, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barredo, J.I.; Strona, G.; de Rigo, D.; Caudullo, G.; Stancanelli, G.; San-Miguel-Ayanz, J. Assessing the potential distribution of insect pests: Case studies on the large pine weevil (Hylobius abietis L.) and horse-chestnut leaf miner (Cameraria ohridella) under present and future climate conditions in European forests. EPPO Bull. 2015, 45, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Godefroid, M.; Cruaud, A.; Rossi, J.P.; Rasplus, J.Y. Assessing the risk of invasion by tephritid fruit flies: Intraspecific divergence matters. PLoS ONE 2015, 10, e0135209. [Google Scholar] [CrossRef]
- Li, B.; Wei, W.; Ma, J.; Zhang, R. Maximum entropy niche-based modeling (Maxent) of potential geographical distributions of fruit flies Dacus bivittatus, D. ciliatus and D. vertebrates (Diptera: Tephritidae). Acta Entomol. Sin. 2009, 52, 1122–1131. [Google Scholar]
- Lv, W.; Li, Z.; Wu, X.; Ni, W.; Qv, W. Maximum entropy niche-based modeling (Maxent) of potential geographical distributions of Lobesia botrana (Lepidoptera: Tortricidae) in China. In Computer and Computing Technologies in Agriculture V, Proceedings of the International Conference on Computer and Computing Technologies in Agriculture, Beijing, China, 29–31 October 2011; Li, D., Chen, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 239–246. [Google Scholar]
- Kim, D.; Lee, H.; Kim, M.; Lee, D. Predicting the potential habitat, host plants, and geographical distribution of Pochazia shantungensis (Hemiptera: Ricaniidae) in Korea. Korean J. Appl. Entomol. 2015, 54, 179–189. [Google Scholar] [CrossRef]
- Baek, S.; Kim, M.; Lee, J. Current and Future Distribution of Ricania shantungensis (Hemiptera: Ricaniidae) in Korea: Application of Spatial Analysis to Select Relevant Environmental Variables for Maxent and CLIMAX Modeling. Forests 2019, 10, 490. [Google Scholar] [CrossRef] [Green Version]
- Fand, B.B.; Kumar, M.; Kamble, A.L. Predicting the potential geographic distribution of cotton mealybug Phenacoccus solenopsis in India based on MAXENT ecological niche model. J. Environ. Biol. 2014, 35, 973–982. [Google Scholar]
- Wei, J.; Zhang, H.; Zhao, W.; Zhao, Q. Niche shifts and the potential distribution of Phenacoccus solenopsis (Hemiptera: Pseudococcidae) under climate change. PLoS ONE 2017, 12, e0180913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WorldClim Global Climate Database. Available online: http://worldclim.org/current (accessed on 22 February 2016).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Clim. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Philippine GIS Data Clearinghouse. Available online: http://www.philgis.org (accessed on 22 February 2016).
- Phillips, S.J. A Brief Tutorial on Maxent. Lessons Conserv. 2009, 3, 108–135. [Google Scholar]
- Byeon, D.; Jung, S.; Lee, W.H. Review of CLIMEX and Maxent for studying species distribution in South Korea. J. Asia Pac. Biodivers. 2018, 11, 325–333. [Google Scholar] [CrossRef]
- Imperial, M.L.R.; Pedrajita, L.B.; Manalo, G.G.; Cueto, C.A.; Almarinez, B.J.M.; Amalin, D.M. Updates on the Mass Rearing and Preliminary Field Release Evaluation of Comperiella calauanica Barrion, Almarinez & Amalin (Hymenoptera: Encyrtidae) for Biological Control of the Coconut Scale Insect, Aspidiotus rigidus Reyne (Hemiptera: Diaspididae) in the Bicol Region. In Proceedings of the 51st Pest Management Council of the Philippines Annual Scientific Convention, Coron, Palawan, Philippines, 3 July 2019. [Google Scholar]
- Technical Advisory on the Management of the Coconut Scale Insect (Aspidiotus rigidus). Available online: https://pca.gov.ph/pdf/cocoTech/pestandDiseaseManagement/CoconutScaleInsect.pdf (accessed on 7 April 2020).
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A.; NCEAS Predicting Species Distributions Working Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Point | Coordinate (WGS 84) | |
|---|---|---|
| Longitude (°) | Latitude (°) | |
| Batangas, Luzon | ||
| Malvar | 121.1466 | 14.04903 |
| Talisay | 121.0107 | 14.09334 |
| Tanauan | 121.0913 | 14.09887 |
| Sto. Tomas | 121.2198 | 14.05746 |
| Cavite, Luzon | ||
| Silang A | 120.9729 | 14.21884 |
| Silang B | 121.0305 | 14.21385 |
| Tagaytay | 121.0002 | 14.17002 |
| Laguna, Luzon | ||
| Calauan | 121.2579 | 14.09737 |
| Los Baños | 121.2595 | 14.15006 |
| Nagcarlan | 121.4137 | 14.15893 |
| Rizal | 121.4109 | 14.06585 |
| Candelaria | 121.4513 | 13.92844 |
| Alaminos | 121.2481 | 14.06618 |
| San Pablo A | 121.2948 | 14.06757 |
| San Pablo B | 121.3333 | 14.05642 |
| Bataan, Luzon | ||
| Orani A | 120.4545 | 14.76979 |
| Orani B | 120.4546 | 14.76963 |
| Orani C | 120.4561 | 14.77067 |
| Orani D | 120.4558 | 14.77054 |
| Basilan Island, Mindanao | ||
| Isabela City * | 121.9947 | 6.587794 |
| Bioclimatic Variable | Variable Code |
|---|---|
| Annual mean temperature (°C × 10) | bio01 |
| Mean diurnal range (°C × 10) | bio02 |
| Isothermality | bio03 |
| Temperature seasonality | bio04 |
| Maximum temperature of the warmest month (°C × 10) | bio05 |
| Minimum temperature of the coldest month (°C × 10) | bio06 |
| Temperature annual range (°C × 10) | bio07 |
| Mean temperature of the wettest quarter (°C × 10) | bio08 |
| Mean temperature of the driest quarter (°C × 10) | bio09 |
| Mean temperature of warmest quarter (°C × 10) | bio10 |
| Mean temperature of coldest quarter (°C × 10) | bio11 |
| Annual precipitation (mm) | bio12 |
| Precipitation of the wettest month (mm) | bio13 |
| Precipitation of the driest month (mm) | bio14 |
| Precipitation seasonality | bio15 |
| Precipitation of the wettest quarter (mm) | bio16 |
| Precipitation of the driest quarter (mm) | bio17 |
| Precipitation of the warmest quarter (mm) | bio18 |
| Precipitation of the coldest quarter (mm) | bio19 |
| Species | Number of Occurrence Points Used in Maxent Modeling | Test AUC | Reference |
|---|---|---|---|
| Anastrepha fraterculus | 49 | 0.76 * | Godefroid et al. (2015) |
| A. obliqua | 49 | 0.77 * | Godefroid et al. (2015) |
| Bactrocera cucurbitae | 49 | 0.91 * | Godefroid et al. (2015) |
| B. oleae | 49 | 0.97 * | Godefroid et al. (2015) |
| Cameraria ohridella | 152 | 0.97 | Barredo et al. (2015) |
| Ceratitis fasciventris | 49 | 0.82 * | Godefroid et al. (2015) |
| Comperiella calauanica | 20 | 0.966 | Current study |
| Hylobius abietis | 677 | 0.93 | Barredo et al. (2015) |
| Lobesia botrana | 95 | 0.970 * | Lv et al. (2012) |
| Phenacoccus solenopsis | 111 | 0.895 | Fand et al. (2014) |
| P. solenopsis | 201 | 0.92 | Wei et al. (2017) |
| Polistes dominula | 9246 | 0.982 * | Howse et al. (2020) |
| Ricania shantungensis | 43 | 0.79 | Baek et al. (2019) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almarinez, B.J.M.; Fadri, M.J.A.; Lasina, R.; Tavera, M.A.A.; Carvajal, T.M.; Watanabe, K.; Legaspi, J.C.; Amalin, D.M. A Bioclimate-Based Maximum Entropy Model for Comperiella calauanica Barrion, Almarinez and Amalin (Hymenoptera: Encyrtidae) in the Philippines. Insects 2021, 12, 26. https://doi.org/10.3390/insects12010026
Almarinez BJM, Fadri MJA, Lasina R, Tavera MAA, Carvajal TM, Watanabe K, Legaspi JC, Amalin DM. A Bioclimate-Based Maximum Entropy Model for Comperiella calauanica Barrion, Almarinez and Amalin (Hymenoptera: Encyrtidae) in the Philippines. Insects. 2021; 12(1):26. https://doi.org/10.3390/insects12010026
Chicago/Turabian StyleAlmarinez, Billy Joel M., Mary Jane A. Fadri, Richard Lasina, Mary Angelique A. Tavera, Thaddeus M. Carvajal, Kozo Watanabe, Jesusa C. Legaspi, and Divina M. Amalin. 2021. "A Bioclimate-Based Maximum Entropy Model for Comperiella calauanica Barrion, Almarinez and Amalin (Hymenoptera: Encyrtidae) in the Philippines" Insects 12, no. 1: 26. https://doi.org/10.3390/insects12010026