Monitoring of Target-Site Mutations Conferring Insecticide Resistance in Spodoptera frugiperda


 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
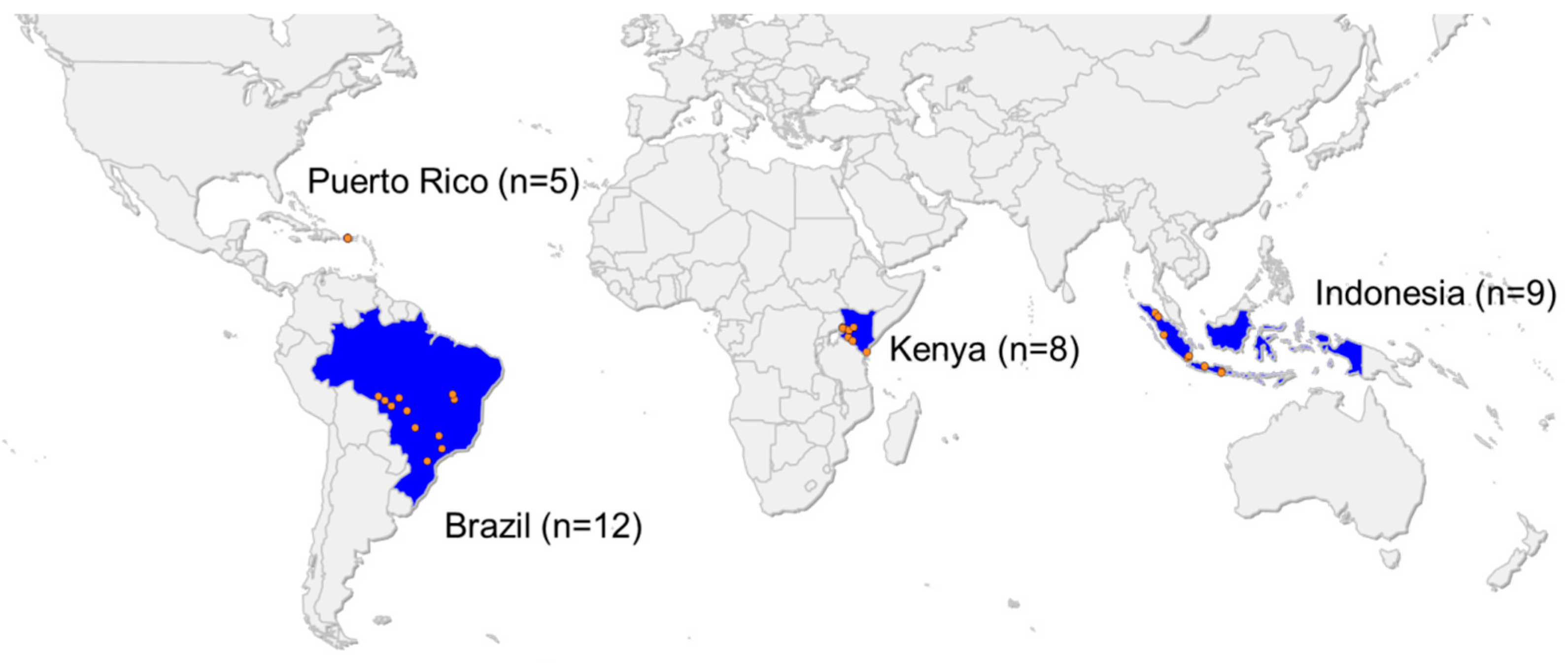
2.1. Insect Collection
2.2. DNA Extraction
2.3. PCR and qPCR Conditions
2.4. Characterization of S. frugiperda Strains
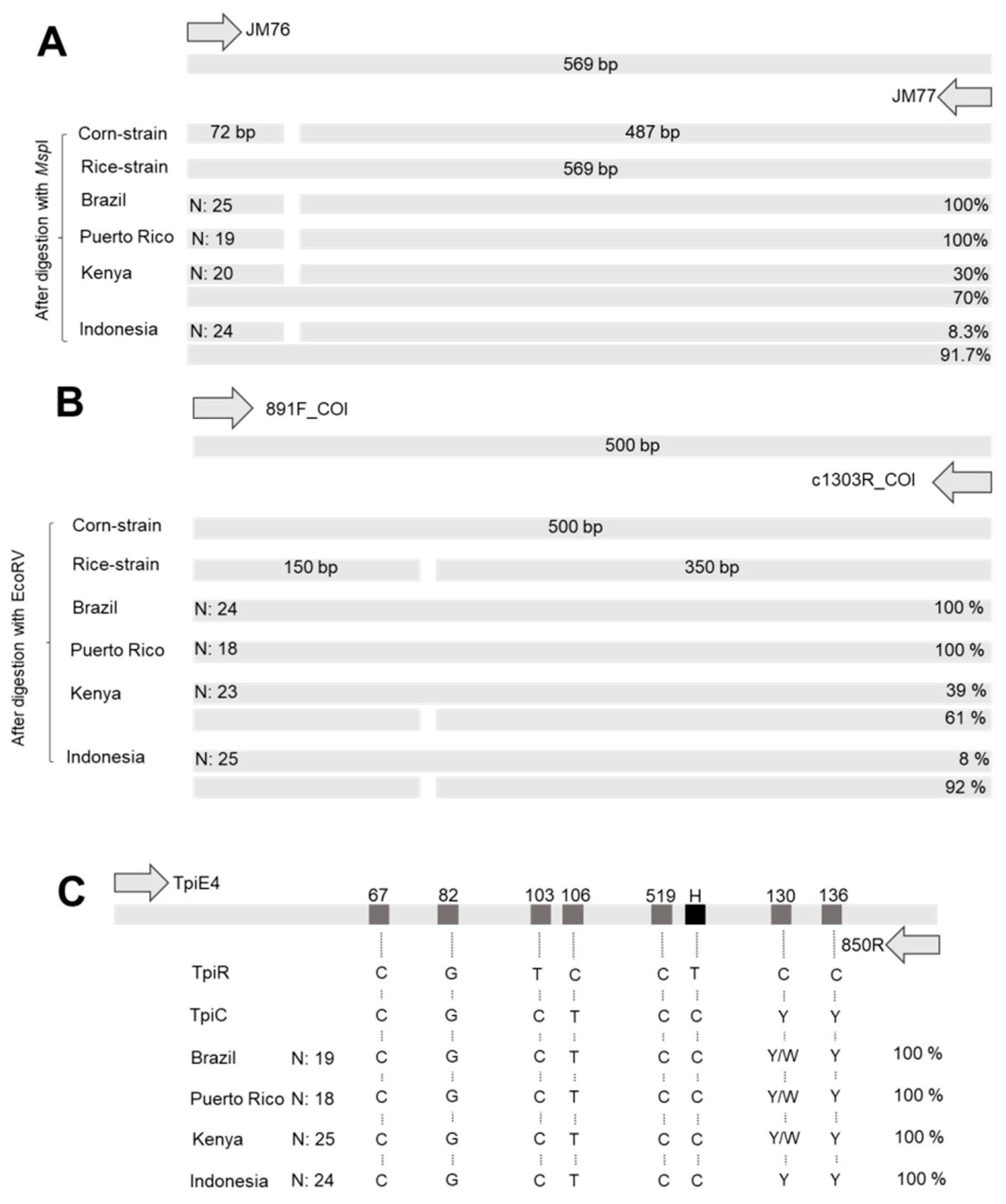
2.4.1. Characterization of COI Haplotypes Using PCR-RFLP
2.4.2. Characterization of Tpi Haplotypes Using DNA Sequencing
2.5. Target-Site Resistance Diagnostics by Pyrosequencing
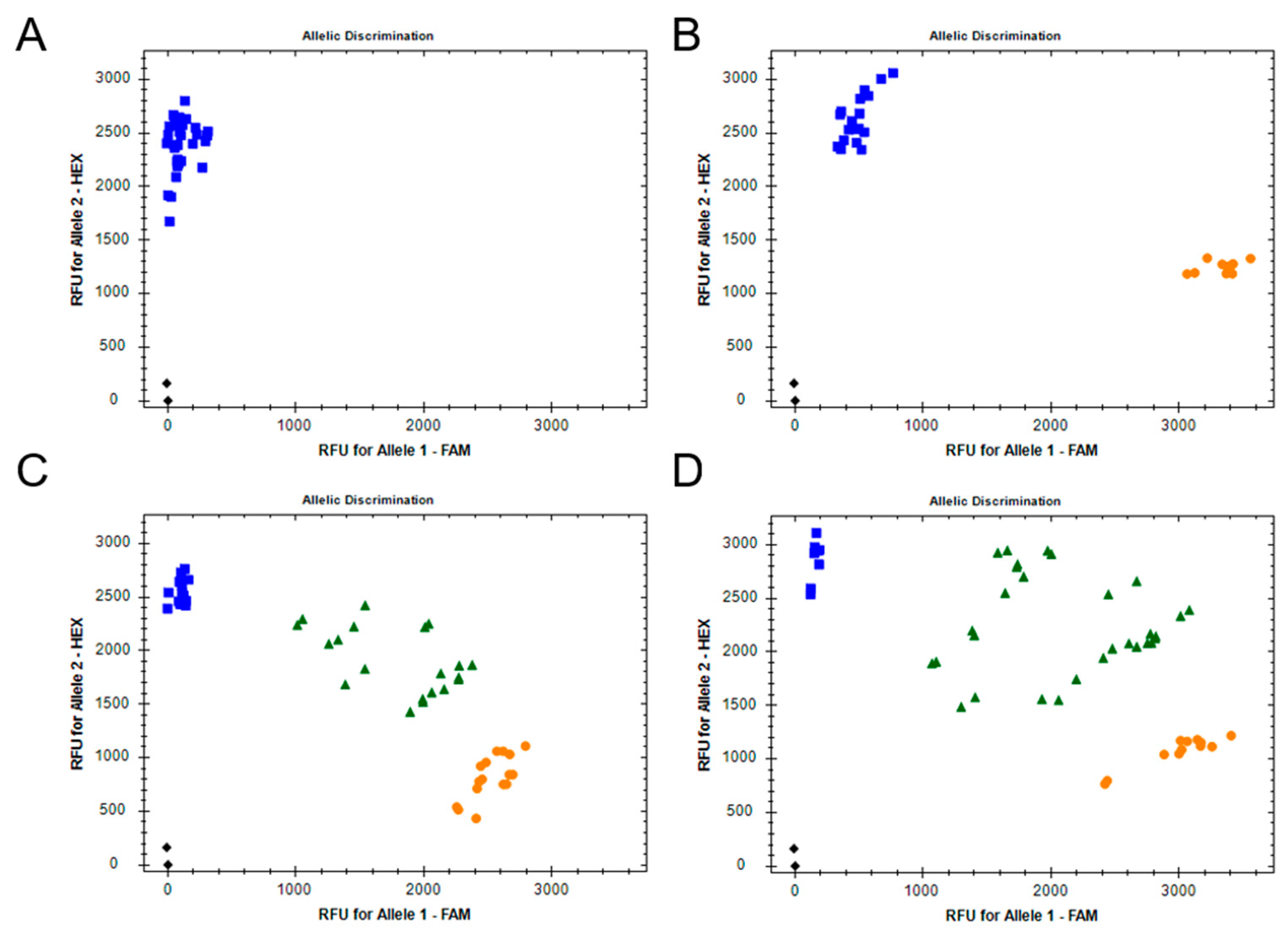
2.6. Fluorescence Based Allelic Discrimination Assays
2.6.1. F290V Mutation in AChE
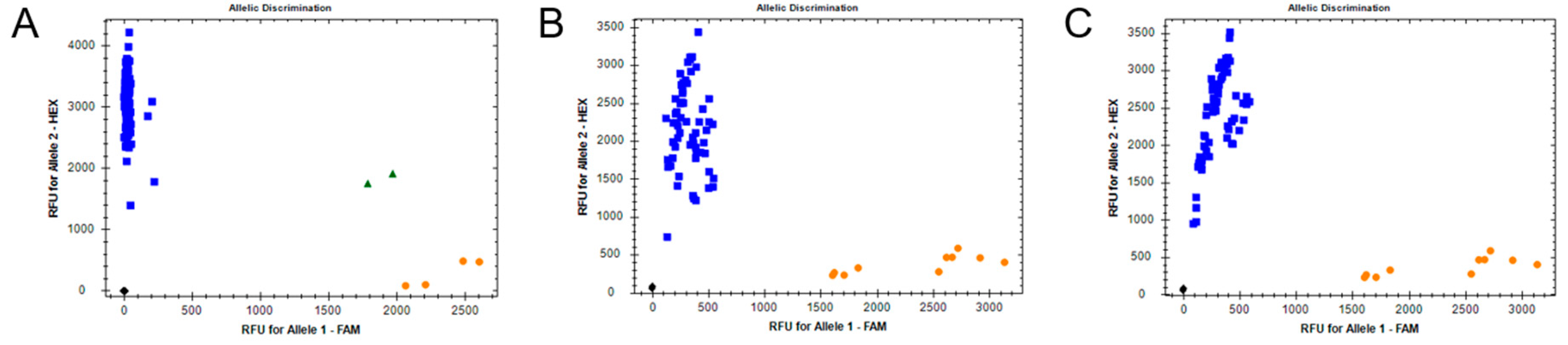
2.6.2. GC Insertion in ABCC2
2.6.3. I4790M Mutation in the RyR
3. Results
3.1. Characterization of S. frugiperda Strains
3.2. Detection of Target-Site Mutations by Pyrosequencing
3.3. Detection of Target-Site Mutations by Fluorescence Based Allelic Discrimination Assays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Luginbill, P. The fall armyworm. U.S. Dept. Agric. Tech. Bull. 1928, 34, 1–91. [Google Scholar]
- Pogue, M.G. A world revision of the genus Spodoptera Gueneé (Lepidoptera: Noctuidae). Mem. Am. Ent. Soc. 2002, 43, 1–202. [Google Scholar]
- EPPO. Available online: https://gd.eppo.int/taxon/LAPHFR/distribution (accessed on 25 March 2020).
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First Report of Outbreaks of the Fall Armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in west and central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalleshwaraswamy, C.M.; Asokan, R.; Swamv, H.M.M.; Marutid, M.S.; Pavithra, H.B.; Hegde’, K.; Navi’, S.; Prabhu’, S.T.; Goergen, G. First report of the Fall armyworm, Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag. Hortic. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- Shylesha, A.N.; Jalali, S.K.; Gupta, A.; Varshney, R.; Venkatesan, T.; Shetty, P.; Ojha, R.; Ganiger, P.C.; Navik, O.; Subaharan, K.; et al. Studies on new invasive pest Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae) and its natural enemies. Biol. Control 2018, 32, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Barros, E.M.; Torres, J.B.; Ruberson, J.R.; Oliveira, M.D. Development of Spodoptera frugiperda on different hosts and damage to reproductive structures in cotton: Fall armyworm performance on different hosts. Entomol. Exp. Appl. 2010, 137, 237–245. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Nagoshi, R.N.; Meagher, R.L.; Fleischer, S.J.; Jairam, S. Modeling seasonal migration of fall armyworm moths. Int. J. Biometeorol. 2016, 60, 255–267. [Google Scholar] [CrossRef]
- Prowell, D.P.; McMichael, M.; Silvain, J.-F. Multilocus Genetic Analysis of Host Use, Introgression, and Speciation in Host Strains of Fall Armyworm (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2004, 97, 1034–1044. [Google Scholar] [CrossRef] [Green Version]
- Veenstra, K.H.; Pashley, D.P.; Ottea, J.A. Host-Plant Adaptation in Fall Armyworm Host Strains: Comparison of Food Consumption, Utilization, and Detoxication Enzyme Activities. Ann. Entomol. Soc. Am. 1995, 88, 80–91. [Google Scholar] [CrossRef]
- Pashley, D.P. Quantitative genetics, development, and physiological adaptation in host strains of fall armyworm. Evolution 1988, 42, 93–102. [Google Scholar] [CrossRef]
- Adamczyk, J.J.; Leonard, B.R.; Graves, J.B. Toxicity of Selected Insecticides to Fall Armyworms (Lepidoptera: Noctuidae) in Laboratory Bioassay Studies. Fla. Entomol. 1999, 82, 230–236. [Google Scholar] [CrossRef]
- Gouin, A.; Bretaudeau, A.; Nam, K.; Gimenez, S.; Aury, J.-M.; Duvic, B.; Hilliou, F.; Durand, N.; Montagné, N.; Darboux, I.; et al. Two genomes of highly polyphagous lepidopteran pests (Spodoptera frugiperda, Noctuidae) with different host-plant ranges. Sci. Rep. 2017, 7, 11816. [Google Scholar] [CrossRef] [PubMed]
- Arias, O.; Cordeiro, E.; Corrêa, A.S.; Domingues, F.A.; Guidolin, A.S.; Omoto, C. Population genetic structure and demographic history of Spodoptera frugiperda (Lepidoptera: Noctuidae): Implications for insect resistance management programs. Pest Manag. Sci. 2019, 75, 2948–2957. [Google Scholar] [CrossRef] [PubMed]
- Silva-Brandão, K.L.; Peruchi, A.; Seraphim, N.; Murad, N.F.; Carvalho, R.A.; Farias, J.R.; Omoto, C.; Cônsoli, F.L.; Figueira, A.; Brandão, M.M. Loci under selection and markers associated with host plant and host-related strains shape the genetic structure of Brazilian populations of Spodoptera frugiperda (Lepidoptera, Noctuidae). PLoS ONE 2018, 13, e0197378. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, R.N.; Htain, N.N.; Boughton, D.; Zhang, L.; Xiao, Y.; Nagoshi, B.Y.; Mota-Sanchez, D. Southeastern Asia fall armyworms are closely related to populations in Africa and India, consistent with common origin and recent migration. Sci. Rep. 2020, 10, 1421. [Google Scholar] [CrossRef] [Green Version]
- Nagoshi, R.N.; Dhanani, I.; Asokan, R.; Mahadevaswamy, H.M.; Kalleshwaraswamy, C.M.; Sharanabasappa; Meagher, R.L. Genetic characterization of fall armyworm infesting South Africa and India indicate recent introduction from a common source population. PLoS ONE 2019, 14, e0217755. [Google Scholar] [CrossRef] [Green Version]
- Nagoshi, R.N.; Goergen, G.; Plessis, H.D.; Van den Berg, J.; Meagher, R. Genetic comparisons of fall armyworm populations from 11 countries spanning sub-Saharan Africa provide insights into strain composition and migratory behaviors. Sci. Rep. 2019, 9, 8311. [Google Scholar] [CrossRef] [Green Version]
- Ingber, D.A.; Mason, C.E.; Flexner, L. Cry1 Bt Susceptibilities of Fall Armyworm (Lepidoptera: Noctuidae) Host Strains. J. Econ. Entomol. 2018, 111, 361–368. [Google Scholar] [CrossRef]
- Pashley, D.P.; Sparks, T.C.; Quisenberry, S.S.; Jamjanya, T. Dowd. Two fall armyworm strains feed on corn, rice and Bermuda-grass. La. Agric. 1987, 30, 8–9. [Google Scholar]
- APRD. Available online: https://www.pesticideresistance.org/ (accessed on 23 March 2020).
- ISAAA. Available online: http://www.argenbio.org/adc/uploads/ISAAA_2017/isaaa-brief-53-2017 (accessed on 23 March 2020).
- Sparks, T.C.; Nauen, R. IRAC: Mode of action classification and insecticide resistance management. Pestic. Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall Armyworm: Impacts and implications for Africa. Outlooks Pest manag. 2017, 28, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Kumela, T.; Simiyu, J.; Sisay, B.; Likhayo, P.; Mendesil, E.; Gohole, L.; Tefera, T. Farmers’ knowledge, perceptions, and management practices of the new invasive pest, fall armyworm (Spodoptera frugiperda) in Ethiopia and Kenya. Int. J. Pest Manag. 2019, 65, 1–9. [Google Scholar] [CrossRef]
- Sisay, B.; Tefera, T.; Wakgari, M.; Ayalew, G.; Mendesil, E. The Efficacy of Selected Synthetic Insecticides and Botanicals against Fall Armyworm, Spodoptera frugiperda, in Maize. Insects 2019, 10, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feyereisen, R. Molecular biology of insecticide resistance. Toxicol. Lett. 1995, 83–90. [Google Scholar] [CrossRef]
- Boaventura, D.; Ulrich, J.; Lueke, B.; Bolzan, A.; Okuma, D.; Gutbrod, O.; Geibel, S.; Zeng, Q.; Dourado, P.M.; Martinelli, S.; et al. Molecular characterization of Cry1F resistance in fall armyworm, Spodoptera frugiperda from Brazil. Insect Biochem. Mol. Biol. 2020, 116, 103280. [Google Scholar] [CrossRef]
- Carvalho, R.A.; Omoto, C.; Field, L.M.; Williamson, M.S.; Bass, C. Investigating the Molecular Mechanisms of Organophosphate and Pyrethroid Resistance in the Fall Armyworm Spodoptera frugiperda. PLoS ONE 2013, 8, e62268. [Google Scholar] [CrossRef] [Green Version]
- Boaventura, D.; Bolzan, A.; Padovez, F.E.; Okuma, D.M.; Omoto, C.; Nauen, R. Detection of a ryanodine receptor target-site mutation in diamide insecticide resistant fall armyworm, Spodoptera frugiperda. Pest. Manag. Sci. 2020, 76, 47–54. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Meagher, R.L.; Hay-Roe, M. Inferring the annual migration patterns of fall armyworm (Lepidoptera: Noctuidae) in the United States from mitochondrial haplotypes: Fall Armyworm Migration. Ecol. Evol. 2012, 2, 1458–1467. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Silvie, P.; Meagher, R.L. Comparison of Haplotype Frequencies Differentiate Fall Armyworm (Lepidoptera: Noctuidae) Corn-Strain Populations from Florida and Brazil. J. Econ. Entomol. 2007, 100, 8. [Google Scholar] [CrossRef]
- Troczka, B.; Zimmer, C.T.; Elias, J.; Schorn, C.; Bass, C.; Davies, T.G.E.; Field, L.M.; Williamson, M.S.; Slater, R.; Nauen, R. Resistance to diamide insecticides in diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae) is associated with a mutation in the membrane-spanning domain of the ryanodine receptor. Insect Biochem. Mol. Biol. 2012, 42, 873–880. [Google Scholar] [CrossRef]
- Banerjee, R.; Hasler, J.; Meagher, R.; Nagoshi, R.; Hietala, L.; Huang, F.; Narva, K.; Jurat-Fuentes, J.L. Mechanism and DNA-based detection of field-evolved resistance to transgenic Bt corn in fall armyworm (Spodoptera frugiperda). Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baudron, F.; Zaman-Allah, M.A.; Chaipa, I.; Chari, N.; Chinwada, P. Understanding the factors influencing fall armyworm (Spodoptera frugiperda J E Smith) damage in African smallholder maize fields and quantifying its impact on yield. A case study in Eastern Zimbabwe. Crop Prot. 2019, 120, 141–150. [Google Scholar] [CrossRef]
- Feldmann, F.; Rieckmann, U.; Winter, S. The spread of the fall armyworm Spodoptera frugiperda in Africa—what should be done next? J. Plant Dis. Prot. 2019, 126, 97–101. [Google Scholar] [CrossRef]
- Gebreziher, H.G. Review on management methods of fall armyworm (Spodoptera frugiperda J E Smith) in Sub-Saharan Africa. ISRN Entomol. 2020, 7, 9–14. [Google Scholar]
- Hruska, A. Fall armyworm (Spodoptera frugiperda) management by smallholders. CAB Rev. 2019, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sharanabasappa, S.; Kalleshwaraswamy, C.M.; Poorani, J.; Maruthi, M.S.; Pavithra, H.B.; Diraviam, J. Natural Enemies of Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), a recent invasive pest on maize in South India. Fla. Entomol. 2019, 102, 619. [Google Scholar] [CrossRef] [Green Version]
- Zibaee, I.; Mahmood, K.; Esmaeily, M.; Bandani, A.R.; Kristensen, M. Organophosphate and pyrethroid resistances in the tomato leaf miner Tuta absoluta (Lepidoptera: Gelechiidae) from Iran. J. Appl. Entomol. 2018, 142, 181–191. [Google Scholar] [CrossRef]
- Blanco, C.; Chiaravalle, W.; Dalla-Rizza, M.; Farias, J.; García-Degano, M.; Gastaminza, G.; Mota-Sánchez, D.; Murúa, M.; Omoto, C.; Pieralisi, B.; et al. Current situation of pests targeted by Bt crops in Latin America. Curr. Opin. Insect Sci. 2016, 15, 131–138. [Google Scholar] [CrossRef]
- Gutiérrez-Moreno, R.; Mota-Sanchez, D.; Blanco, C.A.; Whalon, M.E.; Terán-Santofimio, H.; Rodriguez-Maciel, J.C.; DiFonzo, C. Field-Evolved Resistance of the Fall Armyworm (Lepidoptera: Noctuidae) to Synthetic Insecticides in Puerto Rico and Mexico. J. Econ. Entomol. 2019, 112, 792–802. [Google Scholar] [CrossRef]
- Bolzan, A.; Padovez, F.E.; Nascimento, A.R.; Kaiser, I.S.; Lira, E.C.; Amaral, F.S.; Kanno, R.H.; Malaquias, J.B.; Omoto, C. Selection and characterization of the inheritance of resistance of Spodoptera frugiperda (Lepidoptera: Noctuidae) to chlorantraniliprole and cross-resistance to other diamide insecticides. Pest Manag. Sci. 2019, 75, 2682–2689. [Google Scholar] [CrossRef]
- Diez-Rodríguez, G.I.; Omoto, C. Herança da resistência de Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae) a lambda-cialotrina. Neotrop. Entomol. 2001, 30, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Do Nascimento, A.R.B.; Farias, J.R.; Bernardi, D.; Horikoshi, R.J.; Omoto, C. Genetic basis of Spodoptera frugiperda (Lepidoptera: Noctuidae) resistance to the chitin synthesis inhibitor lufenuron: Inheritance of lufenuron resistance in Spodoptera frugiperda. Pest Manag. Sci. 2016, 72, 810–815. [Google Scholar] [CrossRef] [PubMed]
- Okuma, D.M.; Bernardi, D.; Horikoshi, R.J.; Bernardi, O.; Silva, A.P.; Omoto, C. Inheritance and fitness costs of Spodoptera frugiperda (Lepidoptera: Noctuidae) resistance to spinosad in Brazil. Pest Manag. Sci. 2017, 74, 1441–1448. [Google Scholar] [CrossRef] [PubMed]
- Lira, E.C.; Bolzan, A.; Nascimento, A.R.; Amaral, F.S.; Kanno, R.H.; Kaiser, I.S.; Omoto, C. Resistance of Spodoptera frugiperda (Lepidoptera: Noctuidae) to spinetoram: Inheritance and cross-resistance to spinosad. Pest Manag. Sci. 2020, 76, 2674–2680. [Google Scholar] [CrossRef]
- Nascimento, A.R.B.; Fresia, P.; Cônsoli, F.L.; Omoto, C. Comparative transcriptome analysis of lufenuron-resistant and susceptible strains of Spodoptera frugiperda (Lepidoptera: Noctuidae). BMC Genom. 2015, 16, 985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, A.O.; Khambay, B.P.S.; Williamson, M.S.; Field, L.M.; WAllace, B.A.; Davies, T.G.E. Modelling insecticide-binding sites in the voltage-gated sodium channel. Biochem. J. 2006, 396, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Soderlund, D.M. Molecular mechanisms of pyrethroid insecticide neurotoxicity: Recent advances. Arch. Toxicol. 2012, 86, 165–181. [Google Scholar] [CrossRef] [Green Version]
- Soderlund, D.M.; Knipple, D.C. The molecular biology of knockdown resistance to pyrethroid insecticides. Insect Biochem. Mol. Biol. 2003, 33, 563–577. [Google Scholar] [CrossRef]
- Williamson, M.S.; Martinez-Torres, D.; Hick, C.A.; Devonshire, A.L. Identification of mutations in the housefly para-type sodium channel gene associated with knockdown resistance (kdr) to pyrethroid insecticides. Mol. Genet. Genom. 1996, 252, 51–60. [Google Scholar] [CrossRef]
- Rinkevich, F.D.; Du, Y.; Dong, K. Diversity and convergence of sodium channel mutations involved in resistance to pyrethroids. Pestic. Biochem. Physiol. 2013, 106, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liu, B.; Zheng, W.; Liu, C.; Zhang, D.; Zhao, S.; Xu, P.; Wilson, K.; Withers, A.; Jones, C.M.; et al. High-depth resequencing reveals hybrid population and insecticide resistance characteristics of fall armyworm (Spodoptera frugiperda) invading China. BioRxiv 2019, 813154. [Google Scholar] [CrossRef] [Green Version]
- Harel, M.; Kryger, G.; Rosenberry, T.L.; Mallender, W.D.; Lewis, T.; Fletcher, R.J.; Guss, J.M.; Silman, I.; Sussman, J.L. Three-dimensional structures of Drosophila melanogaster acetylcholinesterase and of its complexes with two potent inhibitors. Protein Sci. 2000, 9, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Kim, J.I.; Lee, D.W.; Chung, B.K.; Miyata, T.; Lee, S.H. Identification and characterization of ace1-type acetylcholinesterase likely associated with organophosphate resistance in Plutella xylostella. Pestic. Biochem. Physiol. 2005, 81, 164–175. [Google Scholar] [CrossRef]
- Cassanelli, S.; Reyes, M.; Rault, M.; Carlo Manicardi, G.; Sauphanor, B. Acetylcholinesterase mutation in an insecticide-resistant population of the codling moth Cydia pomonella (L.). Insect Biochem. Mol. Biol. 2006, 36, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Haddi, K.; Berger, M.; Bielza, P.; Rapisarda, C.; Williamson, M.S.; Moores, G.; Bass, C. Mutation in the ace-1 gene of the tomato leaf miner (Tuta absoluta) associated with organophosphates resistance. J. Appl. Entomol. 2017, 141, 612–619. [Google Scholar] [CrossRef]
- Jiang, D.; Du, Y.; Nomura, Y.; Wang, X.; Wu, Y.; Zhorov, B.S.; Dong, K. Mutations in the transmembrane helix S6 of domain IV confer cockroach sodium channel resistance to sodium channel blocker insecticides and local anesthetics. Insect Biochem. Mol. Biol. 2015, 66, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-W.; Choi, J.Y.; Kim, W.T.; Je, Y.H.; Song, J.T.; Chung, B.K.; Boo, K.S.; Koh, Y.H. Mutations of acetylcholinesterase1 contribute to prothiofos-resistance in Plutella xylostella (L.). Biochem. Biophys. Res. Commun. 2007, 353, 591–597. [Google Scholar] [CrossRef]
- Wang, J.; Wang, B.; Xie, Y.; Sun, S.; Gu, Z.; Ma, L.; Li, F.; Zhao, Y.; Yang, B.; Shen, W.; et al. Functional study on the mutations in the silkworm (Bombyx mori) acetylcholinesterase type 1 gene (Ace1) and its recombinant proteins. Mol. Biol. Rep. 2014, 41, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Richardson, E.B.; Troczka, B.J.; Gutbrod, O.; Davies, T.G.E.; Nauen, R. Diamide resistance: 10 years of lessons from lepidopteran pests. J. Pest Sci. 2020, 93, 911–928. [Google Scholar] [CrossRef] [Green Version]
- Nauen, R.; Steinbach, D. Resistance to Diamide Insecticides in Lepidopteran Pests. In Advances in Insect Control and Resistance Management; Horowitz, A.R., Ishaaya, I., Eds.; Springer: Negev, Palestine, 2016; pp. 219–240. [Google Scholar]
- Flagel, L.; Lee, Y.W.; Wanjugi, H.; Swarup, S.; Brown, A.; Wang, J.; Kraft, E.; Greenplate, J.; Simmons, J.; Adams, N.; et al. Mutational disruption of the Abcc2 gene in fall armyworm, Spodoptera frugiperda, confers resistance to the Cry1Fa and Cry1A.105 insecticidal proteins. Sci. Rep. 2018, 8, 7255. [Google Scholar] [CrossRef] [Green Version]
- Guan, F.; Zhang, J.; Shen, H.; Wang, X.; Padovan, A.; Walsh, T.K.; Tay, W.T.; Gordon, K.H.J.; James, W.; Czepak, C.; et al. Whole-genome sequencing to detect mutations associated with resistance to insecticides and Bt proteins in Spodoptera frugiperda. Insect Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lan, T.; Fang, D.; Gui, F.; Wang, H.; Guo, W.; Cheng, X.; Chang, Y.; He, S.; Lyu, L.; et al. Chromosome level draft genomes of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), an alien invasive pest in China. bioRxiv 2019, 671560. [Google Scholar] [CrossRef] [Green Version]
- Nagoshi, R.N.; Koffi, D.; Agboka, K.; Tounou, K.A.; Banerjee, R.; Jurat-Fuentes, J.L.; Meagher, R.L. Comparative molecular analyses of invasive fall armyworm in Togo reveal strong similarities to populations from the eastern United States and the Greater Antilles. PLoS ONE 2017, 12, e0181982. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, R.N.; Fleischer, S.; Meagher, R.L.; Hay-Roe, M.; Khan, A.; Murúa, M.G.; Silvie, P.; Vergara, C.; Westbrook, J. Fall armyworm migration across the Lesser Antilles and the potential for genetic exchanges between North and South American populations. PLoS ONE 2017, 12, e0171743. [Google Scholar] [CrossRef] [Green Version]
- Guedes, R.N.C. Insecticide resistance, control failure likelihood and the first law of geography. Pest Manag. Sci. 2017, 73, 479–484. [Google Scholar] [CrossRef] [Green Version]
- Jansson, R.K.; Brown, R.; Cartwright, B.; Cox, D.; Dunbar, D.M.; Dybas, R.A.; Eckel, C.; Lasota, J.A.; Mookerjee, P.K.; Norton, J.A.; et al. Emamectin benzoate: A novel avermectin derivative for control of lepidopterous pests. In Proceedings of the 3rd International Workshop on Management of Diamondback Moth and Other Crucifer Pests; MARDI: Kuala Lumpur, Malaysia, 1997; pp. 1–7. [Google Scholar]
- Burtet, L.M.; Bernardi, O.; Melo, A.A.; Pes, M.P.; Strahl, T.T.; Guedes, J.V. Managing fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), with Bt maize and insecticides in southern Brazil: Managing, S. frugiperda with Bt maize and insecticides. Pest Manag. Sci. 2017, 73, 2569–2577. [Google Scholar] [CrossRef]
- Durham, E.W.; Scharf, M.E.; Siegfried, B.D. Toxicity and neurophysiological effects of fipronil and its oxidative sulfone metabolite on European corn borer larvae (Lepidoptera: Crambidae). Pestic. Biochem. Physiol. 2001, 71, 97–106. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Country | Mutation | N | SS (%) | RS (%) | RR (%) |
|---|---|---|---|---|---|---|
| Voltage-gated sodium channel (VGSC) | Brazil | L1014F | 140 | 100.0 | 0.0 | 0.0 |
| Puerto Rico | 70 | 100.0 | 0.0 | 0.0 | ||
| Kenya | 76 | 100.0 | 0.0 | 0.0 | ||
| Indonesia | 110 | 98.2 | 1.8 | 0.0 | ||
| Brazil | L932F | 143 | 100.0 | 0.0 | 0.0 | |
| Puerto Rico | 64 | 100.0 | 0.0 | 0.0 | ||
| Kenya | 75 | 100.0 | 0.0 | 0.0 | ||
| Indonesia | 88 | 100.0 | 0.0 | 0.0 | ||
| Brazil | T929I | 143 | 100.0 | 0.0 | 0.0 | |
| Puerto Rico | 64 | 100.0 | 0.0 | 0.0 | ||
| Kenya | 75 | 100.0 | 0.0 | 0.0 | ||
| Indonesia | 88 | 100.0 | 0.0 | 0.0 | ||
| Acetylcholinesterase (AChE) | Brazil | A201S | 147 | 92.5 | 4.1 | 3.4 |
| Puerto Rico | 29 | 100.0 | 0.0 | 0.0 | ||
| Kenya | 76 | 89.5 | 10.5 | 0.0 | ||
| Indonesia | 85 | 77.6 | 22.4 | 0.0 | ||
| Brazil | F290V | 127 | 55.1 | 44.9 | 0.0 | |
| Puerto Rico | 70 | 4.3 | 10.0 | 85.7 | ||
| Kenya | 76 | 26.3 | 47.4 | 26.3 | ||
| Indonesia | 86 | 19.8 | 55.8 | 24.4 | ||
| Brazil | G227A | 161 | 55.3 | 32.3 | 12.4 | |
| Puerto Rico | 29 | 100.0 | 0.0 | 0.0 | ||
| Kenya | 76 | 100.0 | 0.0 | 0.0 | ||
| Indonesia | 86 | 83.7 | 16.3 | 0.0 | ||
| Ryanodine receptor (RyR) | Brazil | G4946E | 140 | 100.0 | 0.0 | 0.0 |
| Puerto Rico | 70 | 100.0 | 0.0 | 0.0 | ||
| Kenya | 76 | 100.0 | 0.0 | 0.0 | ||
| Indonesia | 90 | 100.0 | 0.0 | 0.0 | ||
| Brazil a | I4790M | 140 | 100.0 | 0.0 | 0.0 | |
| Puerto Rico | 70 | 100.0 | 0.0 | 0.0 | ||
| Kenya | 76 | 100.0 | 0.0 | 0.0 | ||
| Indonesia | 90 | 100.0 | 0.0 | 0.0 | ||
| ATP-binding cassette transporter subfamily C (ABCC2) | Brazil b | GY del | 211 | 39.83 | 14.30 | 45.87 |
| Puerto Rico | 19 | 100.0 | 0.0 | 0.0 | ||
| Kenya | 70 | 100.0 | 0.0 | 0.0 | ||
| Indonesia | 79 | 100.0 | 0.0 | 0.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boaventura, D.; Martin, M.; Pozzebon, A.; Mota-Sanchez, D.; Nauen, R. Monitoring of Target-Site Mutations Conferring Insecticide Resistance in Spodoptera frugiperda. Insects 2020, 11, 545. https://doi.org/10.3390/insects11080545
Boaventura D, Martin M, Pozzebon A, Mota-Sanchez D, Nauen R. Monitoring of Target-Site Mutations Conferring Insecticide Resistance in Spodoptera frugiperda. Insects. 2020; 11(8):545. https://doi.org/10.3390/insects11080545
Chicago/Turabian StyleBoaventura, Debora, Macarena Martin, Alberto Pozzebon, David Mota-Sanchez, and Ralf Nauen. 2020. "Monitoring of Target-Site Mutations Conferring Insecticide Resistance in Spodoptera frugiperda" Insects 11, no. 8: 545. https://doi.org/10.3390/insects11080545