Host Plant Affects Symbiont Abundance in Bemisia tabaci (Hemiptera: Aleyrodidae)


1
College of Plant Protection, Shanxi Agricultural University, Taigu 030801, China
2
State Key Laboratory of Crop Stress Biology in Arid Areas, and Key Laboratory of Integrated Pest Management on the Loess Plateau of Ministry of Agriculture, Northwest A & F University, Yangling 712100, China
3
Wheat Research Center, Bangladesh Agricultural Research Institute, Dinajpur 5200, Bangladesh
*
Author to whom correspondence should be addressed.
Insects 2020, 11(8), 501; https://doi.org/10.3390/insects11080501
Submission received: 5 July 2020
/
Revised: 31 July 2020
/
Accepted: 1 August 2020
/
Published: 4 August 2020
(This article belongs to the Collection Improving IPM of Specialty Crop Pests and Global Food Security)
Abstract
:Simple Summary
The nutritional contributions of symbionts facilitate herbivores’ plant utilization, promoting insects infecting and spreading on host plants. In this study we investigated the effects of host plants on the symbionts of Bemisia tabaci Middle East-Asia Minor 1 (MEAM1) from a nutritional aspect. We found that three host plant-adapted whitefly populations harbored the same symbiont taxa in different quantities. The amount of the primary symbiont Portiera decreased with increasing host-plant essential amino acid proportions in whitefly populations and even in those transferred to different host-plant species to meet the nutritional demands of whiteflies. However, the abundance of the secondary symbionts in whiteflies after host-plant-shifting for one generation showed little correlation with essential amino acid levels of host plants. It demonstrates that host-plant nitrogen nutrition—mainly, essential amino acids—influences the abundance of symbionts, especially Portiera, to meet whiteflies’ nutritional demands, and whiteflies manipulate their symbionts’ quantity governed by the host plant. The nutrient exchanges in symbioses involving multiple partners could provide new ideas for pest control.
Abstract
Symbionts contribute nutrients that allow insects to feed on plants. The whitefly Bemisia tabaci Middle East-Asia Minor 1 (MEAM1) is a polyphagous pest that depends on symbionts to provide key nutrients that are deficient in the diet. Here, we established three whitefly populations on eggplants, cucumbers, and tomatoes and observed that they harbored the same symbiont taxa in different quantities. The amount of the primary symbiont, Portiera, decreased with increasing concentrations of host-plant essential amino acids (EAAs). Whitefly populations transferred to different plant species exhibited fluctuations in Portiera amounts in the first three or four generations; the amount of Portiera increased when whitefly populations were transferred to plant species with lower EAAs proportions. As for the secondary symbionts, the whitefly population of eggplants exhibited lower quantities of Hamiltonella and higher quantities of Rickettsia than the other two populations. The changes of both symbionts’ abundance in whitefly populations after host-plant-shifting for one generation showed little correlation with the EAAs’ proportions of host plants. These findings suggest that host-plant nitrogen nutrition, mainly in the form of EAAs, influences the abundance of symbionts, especially Portiera, to meet the nutritional demands of whiteflies. The results will inform efforts to control pests through manipulating symbionts in insect–symbiont associations.
1. Introduction
The sweet potato whitefly, Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae), is a cosmopolitan pest of over 500 species of horticultural and agronomic crops in fields and greenhouses [1,2]. It feeds on phloem sap, excretes honeydew, and transmits more than 300 plant pathogenic viruses that cause over 40 different diseases in vegetables and fiber crops around the world [3,4]. B. tabaci is responsible for an estimated one to two billion dollars in annual losses through direct or indirect damage [5]. It is a species complex with wide-ranging genetic diversity and is composed of at least 34–36 species that were defined in the past as biotypes [6,7]. Some of the species are restricted to a small number of host plants in specific areas; others are highly polyphagous and global. Of these, B. tabaci Middle East-Asia Minor 1 (MEAM1) (previously termed “biotype B”) is invasive, polyphagous, viruliferous, and the most widespread and damaging group [3,8]. It adapts to a variety of host plants in different habitats.
Plant usage by polyphagous insects may be promoted by microbial symbionts that provide the hosts with nutrients or detoxify plant allelochemicals [9]. The bacterial symbionts in whiteflies are composed of a primary symbiont (P-symbiont), Portiera aleyrodidarum (hereafter, Portiera), occurring in all individuals, and one or more secondary symbionts (S-symbionts) found in some of the individuals of a population [10]. B. tabaci contains at least seven S-symbionts: Hamiltonella, Rickettsia, Wolbachia, Cardinium, Arsenophonus, Fritschea, and Hemipteriphilus [10,11,12,13].
Portiera is essential for the development and reproduction of whiteflies. It is located in specialized bacteriocytes in the body cavity and synthesizes nutrients, such as essential amino acids (EAAs) and carotenoids, which are missing from the phloem diet [4,14]. Although S-symbionts are facultative for whitefly survival and reproduction, they can influence whitefly fitness by mediating a resistance to parasitoids, thermotolerance, viral transmission, and hosts’ susceptibility to insecticides [15]. In addition, some S-symbionts supply vitamins, cofactors, and EAAs to their hosts [16]. The genomes of Hamiltonella and Rickettsia retain some genes involved in the biosynthesis of several EAAs, cofactors, and vitamins [4,17,18,19,20]. While the presence of these genes hints at metabolic cooperation between the S-symbionts and whiteflies [18,21,22], there have been a few studies focusing on the relationship between the amount of S-symbionts and EAAs synthesis.
Variations in the symbiont quantity influences insect fitness. In the aphid Aphis craccivora, the titer of its symbiont, Buchnera, decreases under low and high temperature stress, negatively influencing aphid reproduction [23]. Excessive symbiont density may amplify the negative or positive effects on the insect by influencing its fitness or by causing pathological damage [24]. Symbiont proliferation consumes the resources of the insect body, which are in turn obtained from the host plants. Thus, host plants may significantly influence symbiont numbers. Plants differ in their amino-acid profiles, which may affect symbiont density in polyphagous insects [25]. The fluctuation of symbiont density in whiteflies in response to differences in nitrogen nutrient levels among host plants remains unknown.
Here, we characterized the symbionts in three laboratory-grown whitefly populations that were originally derived from the same parental population and then established on eggplants, tomatoes, or cucumbers. We monitored the symbiont abundance in whiteflies transferred to natal or novel host-plant species for five generations. We also measured the amino acid compositions in the phloem sap of the three plant species and in their corresponding whitefly populations to interpret the dynamics of Portiera density in response to nitrogen nutrition, especially EAAs levels, in different host plants. We found that, in our laboratory-grown whiteflies, different B. tabaci populations harbored the same symbiont taxa in different quantities, and the amounts of Portiera decreased with increasing concentrations of host-plant EAAs in whitefly populations or after host-plant shifting to meet the nutritional demands of whiteflies.
2. Materials and Methods
2.1. Plants and Insects
Three economically important vegetables were used in this study: eggplants (Solanum melongena L. (Solanaceae) var. “Zichangqie”), tomatoes (Solanum lycopersicum L. (Solanaceae) var. “Florida Lanai”), and cucumbers (Cucumis sativus L. (Cucurbitaceae) var. “Jinchun”). Seeds were germinated, and seedlings individually transplanted into plastic pots (12 cm in diameter) with potting mix (a mixer of peat moss, vermiculite, and perlite at a 7:1:1 ratio by volume) in an insectary at 25 ± 1 °C, 60% ± 5% relative humidity (RH), and a photoperiod of 16L:8D (light:dark) at a light intensity of 1400–1725 lux [25]. Seedlings were fertilized with a dry soluble fertilizer “Harvest More 20-20-20+TE” at the rate of 1-g/L water/week. When plants had five to six leaves, some were used to rear whiteflies, and others were used to extract amino acids.
B. tabaci was collected from a greenhouse (Key Laboratory of Applied Entomology, Northwest A&F University, Yangling, Shaanxi, China) in 2011 and subsequently reared on tomatoes (Solanum lycopersicum var. “Florida Lanai”) in the insectary. The whitefly adults were identified as the MEAM1 species (GenBank accession No.: KF773139) using the mitochondrial COI gene [26]. Three laboratory whitefly populations were raised for more than 30 generations in cages (65 × 65 × 65 cm) on eggplants, tomatoes, and cucumbers, separately. These populations were labeled Bemisia-eggplant, Bemisia-tomato, and Bemisia-cucumber, respectively. They had identical genetic backgrounds. The whiteflies were maintained at 26 ± 1 °C, 65% ± 5% RH, with a photoperiod of 16L:8D and a light intensity of 1400–1725 lux [27].
2.2. Identification of Symbionts in Different Whitefly Populations
Total DNA for each sample was extracted from 20 female adults using the Wizard Genomic DNA Purification Kit (Promega Corporation, Madison, WI, USA). The quality of the DNA template was verified by PCR amplification of the mitochondrial COI gene of B. tabaci. Diagnostic PCR with specific primers was used to identify the symbionts infecting the whitefly populations. The 16S rRNA gene for Portiera, Hamiltonella, Rickettsia, Wolbachia, Cardinium, and Hemipteriphilus and the 23S rRNA gene for Arsenophonus and Fritschea were amplified. The corresponding primer sequences are listed in Table 1. PCR reactions were performed with 25-μL reaction mixtures containing 12.5-μL Taq Premix (TaKaRa), 1-μL forward and reverse primers (10 μM) each, 2-μL DNA template, and 8.5-μL ddH2O under the following cycling conditions: 95 °C for 4 min, then 35 cycles of 95 °C for 30 s, 60 °C for 30 s (except for Wolbachia, which was at 55 °C for 30 s), and 72 °C for 1 min, followed by a final extension at 72 °C for 10 min. Amplified DNA products were subjected to electrophoreses on 1% agarose gels, and a specific target band was regarded as diagnostic for the symbiont infection. The species of symbionts detected in the samples were further confirmed by sequencing (Invitrogen, Beijing, China).
2.3. Transfer of Whitefly Populations to Natal and Novel Host-Plant Species
From each population, about 150 five-day-old female adults (termed G0) were collected to measure the symbiont abundance. Between 150 and 200 adults were transferred to the natal or novel species for 24 h to lay eggs. The adults were then removed, and the eggs were allowed to develop. About 150 five-day-old G1 female adults were collected and the symbionts quantified. The remaining G1 adults were transferred to the three plant species to allow egg-laying for 24 h, following which, the adults were removed. About one month later, 150 five-day-old female adults (G2) were collected and stored in ethanol at −20 °C. The remaining G2 adults were transferred to fresh plants for the production of the third generation. The third (G3), fourth (G4), and fifth (G5) generation samples were collected similarly. The plant switch experiments were performed as shown in Figure 1. The G0 and G1 samples with six replicates for each treatment were used to quantify all detected symbionts, and the G2–G5 samples with six replicates for each treatment were sampled to quantify Portiera.
2.4. Quantification of Symbionts
The relative abundance of symbionts in Bemisia-eggplants, Bemisia-tomatoes, Bemisia-cucumbers (G0), and in the three whitefly populations transferred to the natal and novel plant species (G1–G5) was determined by quantitative PCR (qPCR). Total DNA was extracted from 20 female adults for each sample using the Wizard Genomic DNA Purification Kit, according to the manufacturer’s instructions. qPCR was performed on the iQ5 multicolor real-time PCR machine (Bio-Rad) with 20-μL mixtures containing SYBR Premix Ex Taq II (TaKaRa) 10 μL, forward and reverse primers (10 μM) each for 0.8 μL, DNA template 2 μL, and ddH2O 6.4 μL. The reaction program was 95 °C for 3 min, followed by 40 cycles of 95 °C for 10 s and 55 °C for 30 s. Portiera, Hamiltonella, and Rickettsia were quantified by qPCR using the number of 16S rRNA, 16S rRNA, and gltA genes, respectively. β-actin was used as the reference gene. The primers used here are presented in Table 1. All quantifications included six replicates, with three technical repeats for each sample. The relative amount of each symbiont was normalized using β-actin and calculated by the 2-ΔCt method [36,37,38].
2.5. Amino-Acid Analysis
Free amino acids in plant phloem sap were collected using the EDTA-exudation technique [25,39,40]. When eggplants, tomatoes, and cucumbers grew to the 5–6-leaf stage, the second and third fully expanded fresh leaves from the top of the plant were cut. The petioles were immersed in 1-mL EDTA solution (10 mM, pH 7.0) in a 1.5-mL Eppendorf tube and incubated in darkness at 25 °C for 4 h with > 90% RH. Leaves were removed from the tubes, and any droplets attached to the petioles were tapped back into the tube. The exudates were centrifuged at 4500 g for 5 min at 4 °C. The supernatant was filtered through a sterile syringe filter and stored at −80 °C for further analysis. Amino acids were extracted from six biological replicates of each plant species.
Free amino acids in whiteflies were extracted using ethanol and hydrochloric acid [39]. Fifty newly emerged female adults from every population (G0) were ground in a 1.5-mL Eppendorf tube containing a 600-µL buffer of 0.1-M HCl and 100% ethanol (1:1), followed by centrifugation at 12,000 g for 20 min. The supernatant was filtered and kept at −80 °C for subsequent analysis. Amino acids were extracted from nine biological replicates of each whitefly population.
Amino acids in the extracts were analyzed by LC-MS. Liquid chromatography separations were carried out with a Inertsil OSD-4 C18 Column (250 mm × 3.0 mm; GL Sciences Inc., Tokyo, Japan). Amino-acid elution was performed by applying a three-step gradient: 100% A for 8 min, 0–100% B linear for 2 min, 100% B for additional 5 min, and 0–100% A linear for 1 min, holding the system at 100% A for 8 min with a flow rate of 0.3 mL/min. Mobile phase A was an aqueous solution containing 5% acetonitrile and 0.1% formic acid, and mobile phase B was 100% acetonitrile with 0.1% formic acid. The elution pattern of free amino acids was further confirmed by GC-MS using an identical column and the same procedure as described above. LTQ XLTM linear ion trap mass spectrometer (Thermo Scientific, Waltham, MA, USA) was used in the positive electrospray ionization (ESI) mode. Nitrogen was used as the sheath gas (30.0 arbitrary units) and auxiliary gas (5.0 arbitrary units). The spray voltage was 4.5 kV, and the ion transfer capillary temperature was 275 °C. The amino acids were scanned and fragmented by data-dependent MS/MS. Masses of precursor and product ions and collision energy for each amino acid were based on those in Liu et al. [20]. Acquired data were processed by Xcalibur 2.1 software (Thermo Scientific, Waltham, MA, USA). Quantification was based on an external standard amino-acid mixture of a known concentration.
2.6. Data Analysis
All data related to symbiont abundance, and percentages of individual amino acids and entire EAAs (arginine, histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan, and valine) were tested for normality using the Shapiro-Wilk test [41]. The statistical significance of differences between the experimental groups was tested with one-way ANOVA and Tukey’s honest significant difference (HSD) test at p < 0.05 using IBM SPSS Statistics package v. 20.0 (SPSS Inc., Chicago, IL, USA). To evaluate the effects of different factors on the amount of symbionts, two- or three-way ANOVA with Tukey’s test was used, with post facto multiple comparisons of means. Variations among amino acid–composition profiles in phloem sap and whitefly populations were measured by multiple-dimensional principal component analysis (PCA) using SAS version 9.2. Each amino-acid concentration was converted to mole percentage (mol%) and subjected to PCA. Each spot in the PCA plot represented an individual sample, and distances among the spot groups defined the variation in the amino-acid profiles among the treatments. Correlation between the abundance of Portiera and the proportion of EAAs in plants was tested by Spearman’s rho rank correlation coefficient analysis using SPSS 20.0.
3. Results
3.1. Different Quantities of the Same Symbiont Taxa in Three Whitefly Populations
We identified the P-symbiont Portiera and two S-symbionts taxa, Hamiltonella and Rickettsia, in the three whitefly populations established on eggplants, cucumbers, and tomatoes by diagnostic PCR (GenBank accession No.: KF773136–KF773138).
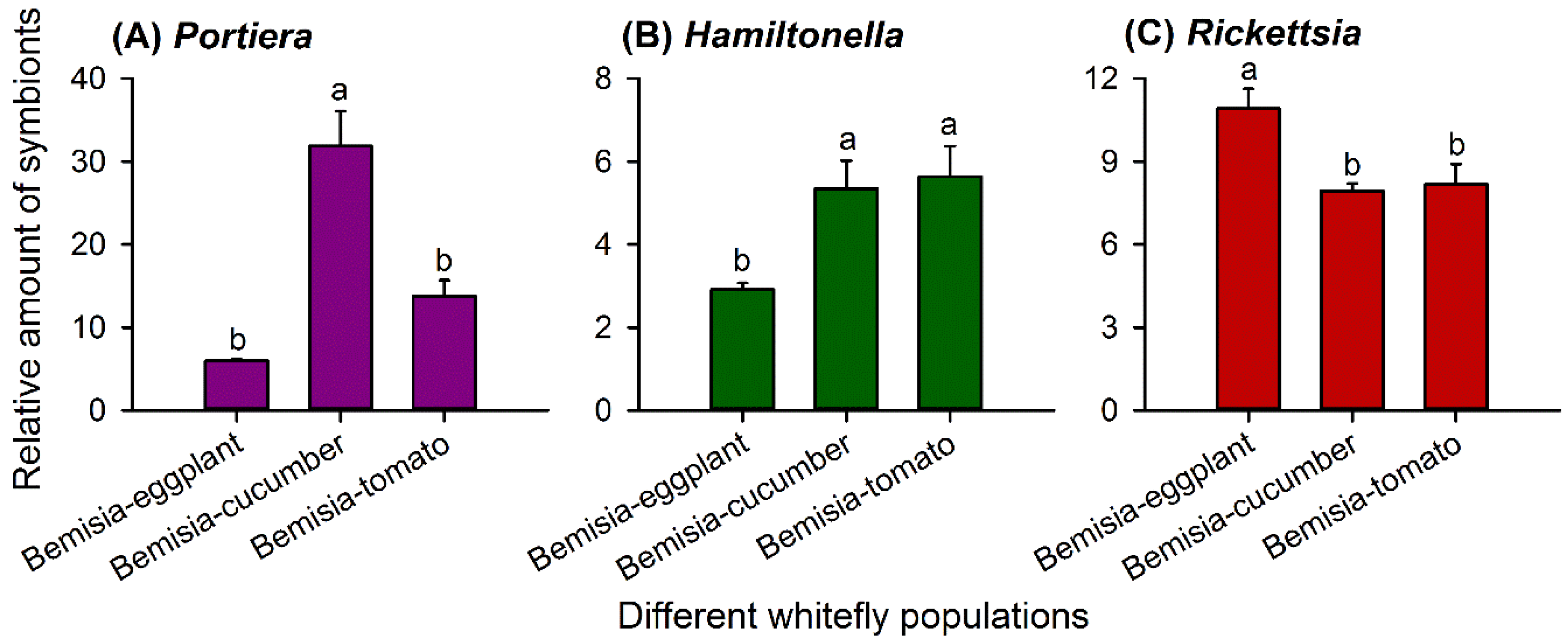
The relative abundance of Portiera differed significantly among the three G0 populations (F2,15 = 24.532, p < 0.001). Bemisia-cucumbers harbored significantly more Portiera than Bemisia-eggplants and Bemisia-tomatoes (Figure 2A).
Similarly, the relative amount of Hamiltonella (F2,15 = 6.558, p = 0.009) and Rickettsia (F2,15 = 7.255, p = 0.006) differed significantly among the three populations. Bemisia-eggplants hosted a lower amount of Hamiltonella (Figure 2B) but significantly higher amounts of Rickettsia (Figure 2C) than the other two populations.
3.2. Host-Plant-Switching Affects Portiera Abundance
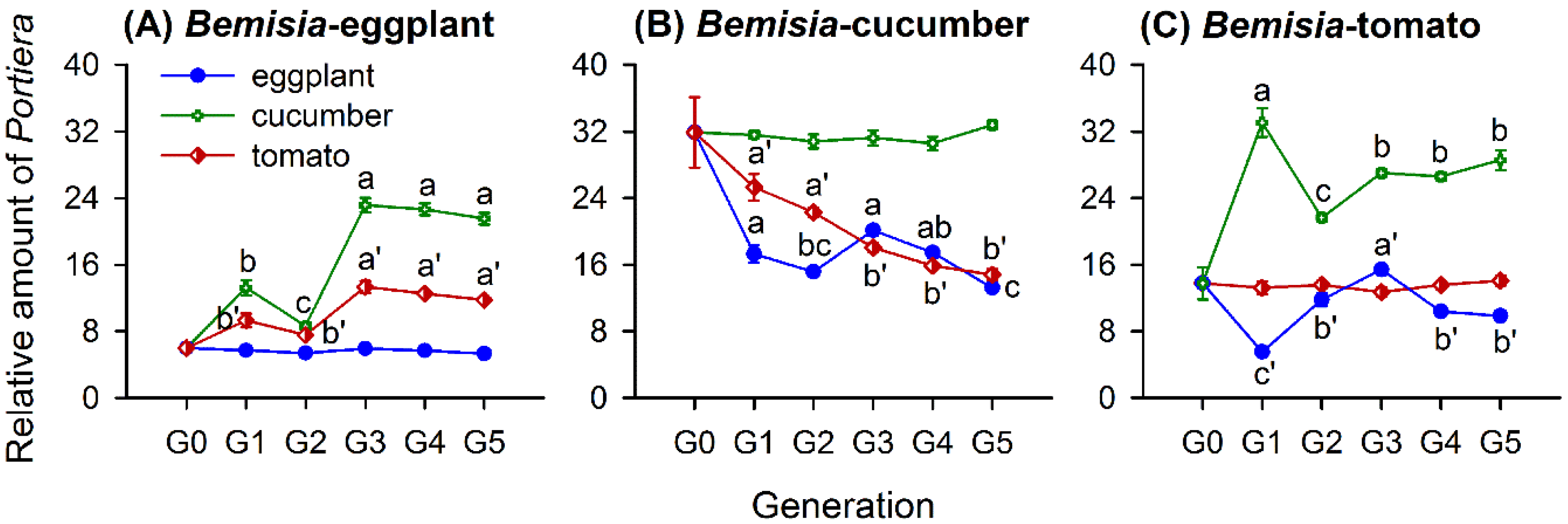
The whitefly population, host-plant species, and the generation number since transfer all affected the relative amount of Portiera, with significant interactions among them (Table 2). In Figure 3, the relative amount of Portiera in the population of Bemisia-eggplants differed significantly across the five generations (G1–G5) when transferred to the cucumbers (F4,25 = 77.198, p < 0.001) and tomatoes (F4,25 = 19.407, p < 0.001), but on the natal plant species, it contained similar amounts of Portiera from G0 to G5 (F5,30 = 1.857, p = 0.132). The Portiera amounts in Bemisia-eggplants increased after a switch to a novel plant species (Figure 3A). It varied greatly in the first three generations and stabilized in G3, G4, and G5 on the cucumbers and tomatoes (Figure 3A).
The amount of Portiera in the Bemisia-cucumbers population showed significant variations across G1 to G5 when transferred to the eggplants (F4,25 = 15.026, p < 0.001) and tomatoes (F4,25 = 25.799, p < 0.001) and remained similar when shifted to other cucumber plants across G0 to G5 (F5,30 = 0.189, p = 0.964). The Portiera levels in Bemisia-cucumbers decreased over the course of five generations on the eggplants and the first three generations on the tomatoes but stabilized to similar levels among G3, G4, and G5 on the tomatoes (Figure 3B).
In Bemisia-tomatoes, the amount of Portiera varied among the five generations when transferred to the eggplants (F4,25 = 39.090, p < 0.001) and cucumbers (F4,25 = 16.526, p < 0.001), while there was no difference from G0 to G5 on the natal plant species (F5,30 = 0.255, p = 0.934). When shifted to the eggplants, the amount of Portiera decreased firstly and then increased, with large variations in the first three generations, followed by stable levels in G4 and G5 (Figure 3C). When transferred to the cucumbers, the Portiera amounts showed fluctuating increases in the first three generations but with similar levels in G3, G4, and G5 (Figure 3C).
3.3. Host-Plant-Switching Affects S-Symbiont Abundance
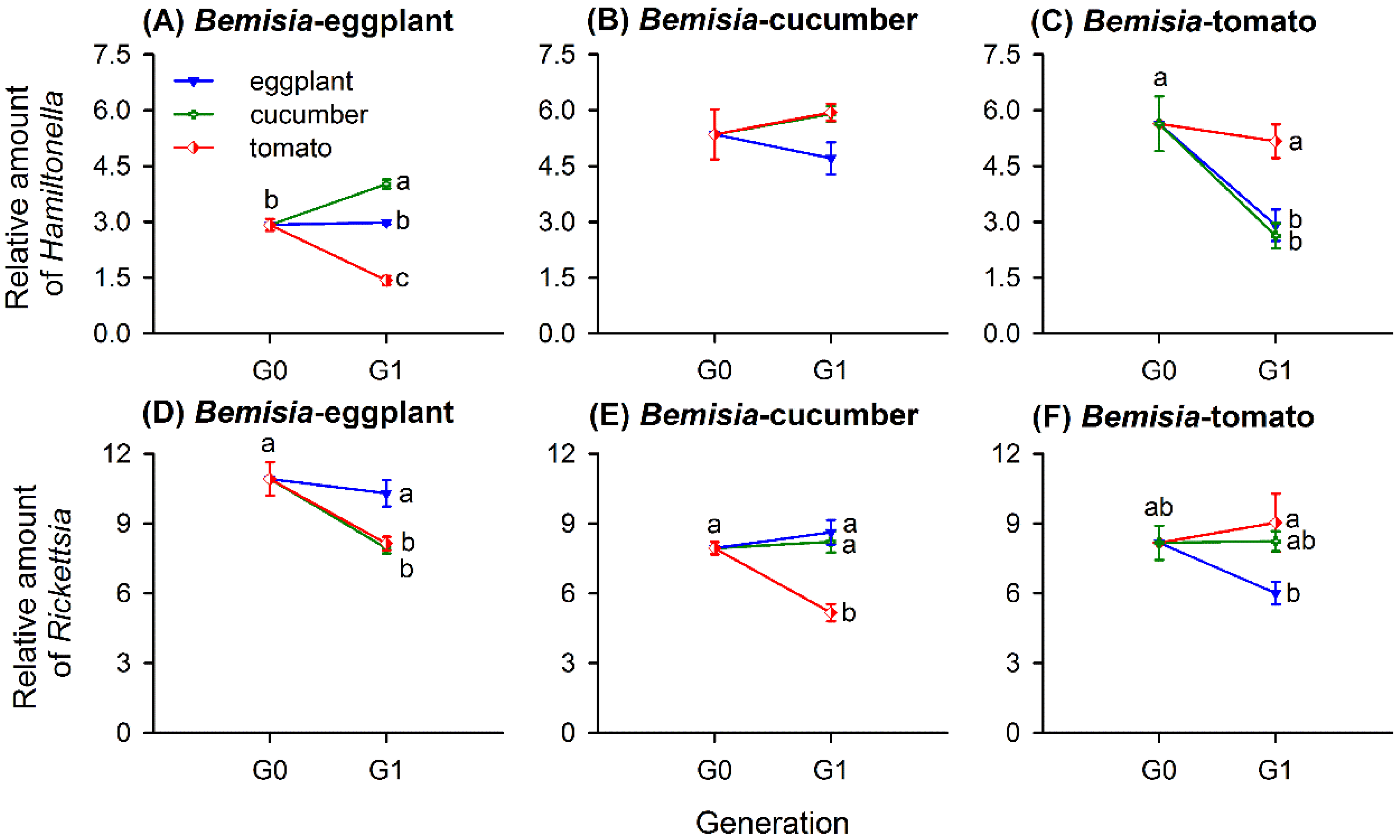
When transferred to novel host-plant species, Hamiltonella amounts changed significantly in a natal population-dependent manner (Table 3). However, Rickettsia amounts changed based on both the natal whitefly population and on the novel host, with a significant interaction term (Table 3). When transferred to novel plant species, Bemisia-eggplants greatly changed in their Hamiltonella (F3,20 = 83.668, p < 0.001) and Rickettsia (F3,20 = 11.584, p < 0.001) quantities but did not differ when shifted to a natal plant species. Figure 4A describes Hamiltonella amounts in Bemisia-eggplants significantly increased when transferred to cucumbers but decreased greatly when moved to tomatoes. Rickettsia amounts reduced considerably when transferred to either cucumbers or tomatoes (Figure 4D).
Host-plant-shifting exerted no effect on Hamiltonella amounts of Bemisia-cucumbers (F3,20 = 1.887, p = 0.164; Figure 4B) but reduced Rickettsia amounts when moved to tomatoes (F3,20 = 17.368, p < 0.001; Figure 4E).
In Bemisia-tomatoes, host-plant-switching influenced the amounts of both Hamiltonella (F3,20 = 10.310, p < 0.001) and Rickettsia (F3,20 = 4.260, p = 0.018). The amount of Hamiltonella in Bemisia-tomatoes decreased when transferred to either novel plant species but exhibited similar levels on other tomatoes (Figure 4C). The amount of Rickettsia remained stable when transferred to the cucumbers and the natal species tomatoes but was significantly reduced when moved to the eggplants (Figure 4F).
3.4. Phloem Sap Amino Acid–Composition Profiles Differ among Plant Species
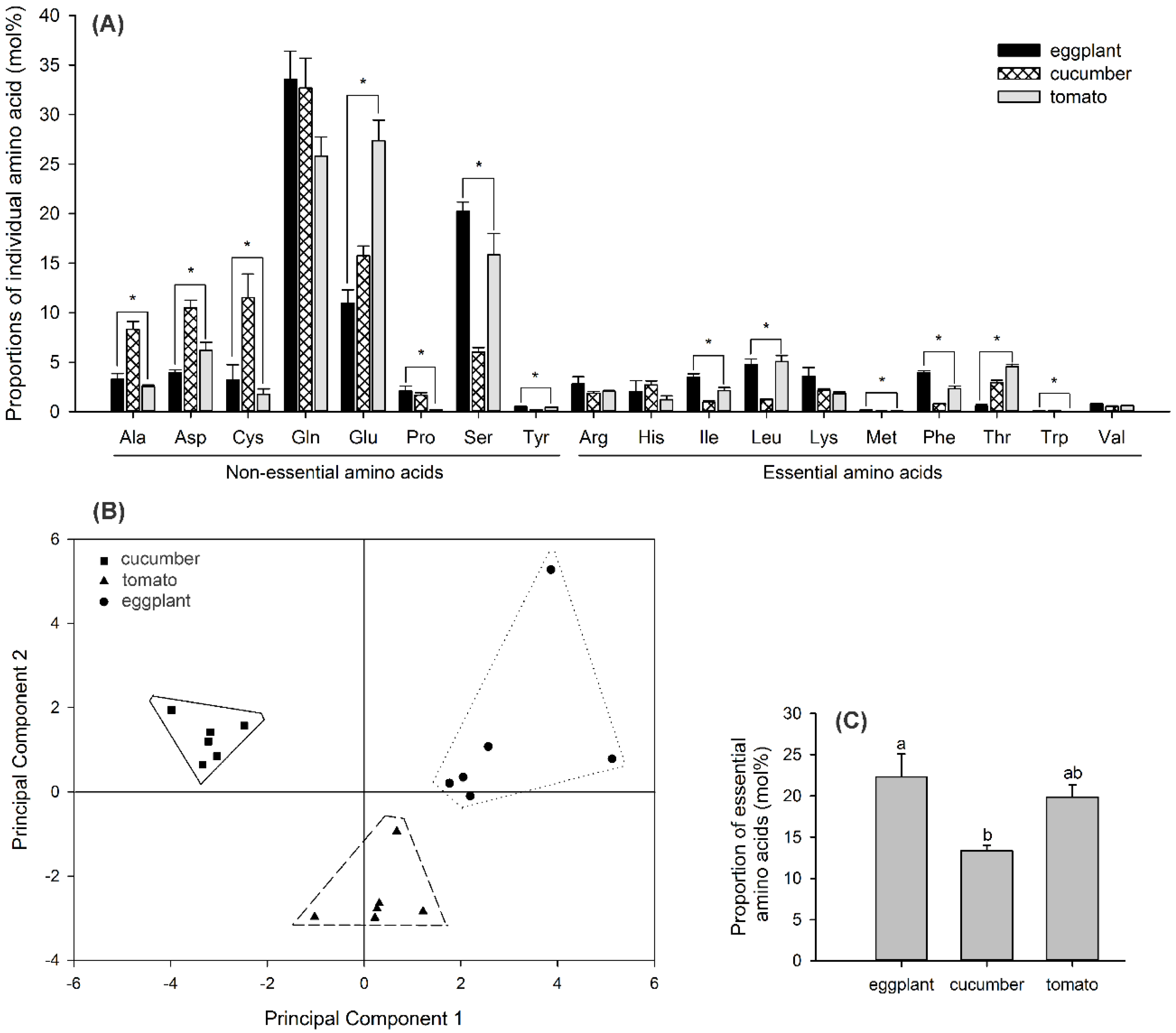
There are 18 amino acids detected in the phloem sap of the eggplant, cucumber, and tomato leaves. The proportions of most individual amino acids varied significantly among the different plant species (Figure 5A), and the statistical analyses of these amino acids are shown in Table 4. For each plant species, the three most abundant amino acids were non-EAAs: glutamine (33.6%), serine (20.2%), and glutamic acid (11.0%) in eggplants; glutamine (32.7%), glutamic acid (15.7%), and cysteine (11.5%) in cucumbers; and glutamic acid (27.4%), glutamine (25.8%), and serine (15.8%) in tomatoes. Among the EAAs, isoleucine, leucine, methionine, and phenylalanine occurred in larger amounts in eggplants than in cucumbers (Figure 5A). PCA indicated lower variations in the amino acid–composition profiles within the sample replicates than among the plant species (Figure 5B). The first principal component explained 41% of the total variation and showed a strong positive association with leucine, isoleucine, phenylalanine, methionine, tyrosine, and serine levels and a negative association with alanine and aspartic acid levels. The second principal component accounted for 27% of the total variation and showed a strong positive association with tryptophan, histidine, lysine, and cysteine levels and a negative association with glutamic acid levels. The percentage of the entire EAAs differed significantly among the plant species (F2,15 = 6.065, p = 0.012). Eggplants had the highest percentage of the entire EAAs, cucumbers had the lowest, and tomatoes had an intermediate percentage that was not significantly different from either (Figure 5C). The percentage of the entire EAAs in the three plant species correlated negatively with the Portiera amount in the corresponding whitefly populations (Spearman’s rho: r = −0.473, p = 0.048).
3.5. No Significant Differences in Amino Acid–Composition Profiles among Whitefly Populations
A total of 17 amino acids detected in the whitefly populations. The proportions of most individual amino acids differed significantly among the whitefly populations (Figure 6A), and the statistical analyses of these amino acids are shown in Table 4. PCA revealed no significant differences in the amino acid–composition profiles among the three whitefly populations. Bemisia-cucumbers were separable from Bemisia-eggplants and Bemisia-tomatoes, but the latter two populations could not be distinguished based on the amino-acid profiles (Figure 6B). The first principal component accounted for 36% of the total variation and demonstrated a strong positive association with isoleucine, tryptophan, proline, and tyrosine levels and a negative association with arginine levels. The second principal component explained 20% of the total variation and had a strong positive association with alanine, glutamic acid, and serine levels and a negative association with leucine and lysine levels. However, the percentage of the entire EAAs differed significantly among the whitefly populations (F2,24 = 88.777, p < 0.001). Bemisia-cucumbers contained a significantly higher percentage of the entire EAAs than Bemisia-tomatoes and Bemisia-eggplants. Bemisia-eggplants had the lowest percentage of the three populations (Figure 6C).
4. Discussion
Host-plant nutrition is important for the performance of herbivorous insects, especially sap suckers [25]. These insects are highly dependent on nutrient provisioning by intracellular symbionts [42]. Our study indicated that host plants substantially influence the abundance, but not the taxa, of symbionts in B. tabaci MEAM1. The S-symbionts infecting our study population are Hamiltonella and Rickettsia, which are comparable to those documented in the same B. tabaci species in China, Israel, and Brazil [6,11,12,43]. We established three experimental populations from the same parental population and detected the same symbiont taxa in all three experimental populations. Usually, the change of the symbiont taxa in the insect host is associated with horizontal transmission and environmental factors. A new infection of S-symbionts is through occasional events of horizontal transmissions among different insect species or different individuals mediated by plant, parasitoids, or copulation [44,45,46,47,48]. High temperatures can eliminate the mutualistic partners from insect hosts; many bacteriocyte-associated symbionts have reduced densities or are lost entirely when the insect host is exposed to high temperatures [49,50,51]. Our experimental populations did not contact other infectious populations, species, or host plants fed by other infectious populations and were kept in controlled conditions in the laboratory. Accordingly, we speculate that coinfections of new symbionts or the elimination of existing symbionts might not occur without contact with different infectious populations or without changing the environmental factors.
The host-plant species strongly affected the abundance of different symbionts in B. tabaci. In our study, the amount of symbionts, including Portiera, Hamiltonella, and Rickettsia differed significantly in the three whitefly populations grown on different host-plant species (Figure 2) and after transferring to novel host-plant species (Figure 3 and Figure 4). Likewise, previous studies reported that the relative amount of Portiera, Hamiltonella, Rickettsia, and Cardinium in MEAM1 and Portiera and Hamiltonella amounts in B. tabaci Mediterranean (MED) (previously termed “biotype Q”) were greatly influenced by the host plants, even with the same parental whitefly [36,52]. The host-adapted MEAM1 population on the cucumbers harbored more Portiera than the populations on the cabbages and cotton, and the cabbage population harbored more Hamiltonella, Rickettsia, and Cardinium than the cucumber and cotton populations [36]. However, the MED species of the cabbage population harbored more Portiera than those of the cucumber and cotton populations, and the cucumber and cabbage populations harbored more Hamiltonella than that of the cotton population [52]. In the green peach aphid Myzus persicae, the quantities of the symbionts Buchnera and Serratia symbiotica differed significantly among the aphid populations that fed on different host-plant species [25]. The amount of Buchnera differed significantly in populations of the cotton aphid Aphis gossypii, that had been reared on different host plants over a long duration [53]. Thus, long-term associations between phloem sap-sucking insects and their host plants affect the amount of symbionts that they host.
Although the total nitrogen content of the plant tissue is commonly used as the index of the nutritional value of plants for insects, the nitrogen quality—mainly, the ten EAAs—can be of crucial nutritional importance [54]. We observed that amino acids in different host-plant phloem saps predominantly comprised non-EAAs (EAAs: 13.4–22.3%, Figure 5A,C), but amino acids in different whitefly populations mainly comprised EAAs (51.3–65.0%, Figure 6A,C). PCAs showed that amino acid–composition profiles differed among host-plant species (Figure 5B) but not among the whitefly populations (Figure 6B). These data imply an essential role for the symbionts in providing hosts with EAAs. Eggplant was a more nutritious host plant—due to the higher proportion of EAAs—than tomato and cucumber, and that cucumber was the least nutritious host plant, with the lowest percentage of EAAs (Figure 5C). In an earlier study, we showed that B. tabaci MEAM1 had better fitness, i.e., a shorter developmental time and higher immature survival rate, on eggplants than on other plants [27]. Studies consistently indicate that eggplant is the most suitable host plant for different populations of B. argentifolii (B. tabaci MEAM1) among cucumber, sweet pepper, tomato, and garden bean, based on life-table analyses [55,56]. While the suitability for tomatoes and cucumbers differed depending on B. argentifolii populations and tested cultivars [55,56]. For the Japanese population, tomatoes appeared to be the least suitable host plant among eggplants, cucumbers, and sweet peppers [55], while the Florida B. tabaci population that fed on tomatoes had a higher intrinsic rate of increase than that on cucumbers [56]. These results highlight the importance of nitrogen as a limiting factor in the nutrition of whiteflies.
The nitrogen nutrition of the host plant impacts the population density of symbionts. We observed that the whitefly population established on eggplants with a higher EAA proportion harbored a lower abundance of Portiera (Figure 2 and Figure 5). After host-plant-shifting, the Portiera amount in whiteflies increased when transferred to new plant species with lower percentages of EAAs (Figure 3 and Figure 5). It demonstrates that the whitefly could manipulate its symbiont density to compensate for the deficiencies in the nitrogen nutrition of the host plant. The proliferation of symbionts relies on the consumption of resources from the insect body; consequently, infection densities may be significantly influenced by the nutritional condition of host plants fed by the insect [54]. Previous studies also indicate the important role of host-plant or diet nutrition in symbiont regulation [9,24,54,57]. Studies on aphids show that Buchnera densities in Acyrthosiphon pisum and M. persicae increase with increasing nitrogen levels in the aphid diet or host plants [19,20], while Aphis fabae has elevated Buchnera densities on Lamium purpureum, with a lower amino acid content than Vicia faba [57]. These inconsistencies among whitefly and aphids and among different aphid species demonstrate that the density of the P-symbionts is not maintained at a fixed association with host plants and can vary, possibly to meet the nutritional demands of the insect host, and, ultimately, influences insects’ fitness [24,58]. Even so, we should find out some more evidence to support fine-tuned symbiont densities in B. tabaci MEAM1 in response to the changes of nitrogen nutrition in plants or diets in our future work. Symbionts control the host-plant utilization of insects by regulating the host plant-derived carbon and nitrogen inputs to bacteriocytes [42,53]. Compared with the bacteriocytes of aphids, which are inhabited only by Buchnera, in whiteflies, most of the S-symbionts share bacteriocytes with Portiera [43]. Therefore, Portiera competes with the coresident S-symbionts for nutrients and space, which may explain the different responses observed with Buchnera in aphids. The impacts of symbionts on their insect hosts are determined by the balance between the cost of nutrients consumed by the symbionts and the benefits of nutrients released back to the host [20]. Therefore, elucidating the mechanisms underlying the nutrient exchange in symbioses involving multiple symbionts could provide new ideas for pest control.
Since host-plant-shifting, the relative amount of Portiera in whitefly populations changed on the new host-plant species but did not differ on the natal plant species with which the whitefly populations had over 3 years of interactions (Figure 3). Judging from the dynamics of the Portiera amounts, we assumed that, after transferring to a new host-plant species for many more generations, Portiera in whiteflies will reach an amount similar to the corresponding whitefly populations established on this plant species. When A. gossypii are transferred to novel host plants, Buchnera densities fluctuated in the first two generations and became stable if they continued feeding on that plant species [53]. Therefore, variations in the Portiera abundance after transferring to new host plants may reflect a process of nutritional compensation critical to whitefly survival, since it confers whiteflies with the ability to expand their host-plant range.
With regard to the S-symbionts, both Hamiltonella and Rickettsia genomes keep most of genes involved in synthesis of two EAAs: phenylalanine (Phe) and lysine (Lys), which are missing in the genome of Portiera [4,17,18,19,20,21]. In the present study, lower quantities of Hamiltonella and higher quantities of Rickettsia were found in the whitefly populations on eggplants, a more-nitrogen diet (Figure 2 and Figure 5); the former showed similar changes with Portiera amounts in response to different host plants. In A. pisum, the population density of the S-symbiont S. symbiotica increased when reared on low-nitrogen diets, indicating distinct regulatory mechanisms with the P-symbiont Buchnera [24]. However, similar with Buchnera, the density of Regiella insecticola and Hamiltonella defensa in A. fabae increased in L. purpureum, a low-nitrogen diet [57]. Therefore, variations of the S-symbionts in response to diet nutrition may be attributed to differences between symbionts in metabolic capabilities or access to insect nutrients. In our study, Rickettsia showed an opposite trend in abundance with Hamiltonella, which was inconsistent with the previous study [36]. The relative amounts of different S-symbionts, i.e., Hamiltonella, Rickettsia, and Cardinium, had the same trend in different whitefly populations established on cotton, cucumbers, and cabbages [36]. The discrepancy may be caused by the whitefly genetic background, interactions between S-symbiont taxa, and population inertia. Recent research indicated that Hamiltonella can synthesize biotin; biotin provisioned by whitefly horizontally transferred genes (HTGs) affects the survival and fecundity of adult whiteflies [22]. A Hamiltonella deficiency reduced the level of B vitamin but not EAAs and affected the sex ratio, so this symbiont affects sex ratios in B. tabaci MEAM1 by regulating the fertilization and supplying B vitamins [21]. Except for the nutritional functions, S-symbionts may confer ecologically important traits, e.g., a resistance to parasitoids, thermotolerance, viral transmission, and hosts’ susceptibility to insecticides [15]. B. tabaci MED infected with Hamiltonella grow faster under nutritional stress conditions [14]. MEAM1 species infected with Rickettsia were substantially fitter than uninfected ones: they produced more offspring, especially daughters, had a higher survival to adult, and developed faster [59,60]. We found that after host-plant-shifting for one generation, the changes of both S-symbionts’ abundance showed little correlation with the EAA proportions of the host plants (Figure 4 and Figure 5). Therefore, many generations of host-plant-shifting need to be observed to clarify the correlation. We speculate that the S-symbionts infecting our whitefly populations may confer nutrition or fitness advantages by complicated interactions among Hamiltonella, Rickettsia, and Portiera and even insect hosts when feeding on different host plants.
Host plants influence symbionts in two major ways: nutrients and phytotoxins [53]. Phytotoxins, or secondary metabolites, can exert positive or negative effects on the proliferation of symbionts. Many plant secondary metabolites have bacteriostatic and bactericidal activities [61]. These vary across host plants [53,61]. Future studies may explore the effects of phytotoxins on the abundance of symbionts to explain the changes of symbionts’ abundance after host-plant-shifting.
5. Conclusions
Host plants affect the abundance, but not the taxa, of symbionts in three host-plant-adapted whitefly populations. The abundance of the P-symbiont Portiera in the whitefly populations decreased with the increase of the nitrogen levels of the host plant. Whitefly populations transferred to different host-plant species exhibited fluctuations in Portiera amounts in the first three or four generations; the amount of Portiera increased when the whitefly populations were transferred to lower-nitrogen nutrition plant species. The amount of S-symbionts changed based on a natal population or both the natal population and novel host species after host-plant-shifting, exhibiting little correlation with the nutritional quality of the host plants for just one generation. The whitefly could manipulate its symbionts’ quantity governed by the host plants. Further investigations are required to better understand the cooperative metabolism in multi-partner associations in insect, symbionts and host plants.
Author Contributions
Conceived and designed the experiments: Y.-H.L., M.M.R.S., and T.-X.L. Performed the experiments: Y.-H.L. and M.M.R.S. Analyzed the data: Y.-H.L. and Y.S. Wrote the paper: Y.-H.L., M.M.R.S., and T.-X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the China Agriculture Research System (CARS-25), the Open Foundation of Key Laboratory of Integrated Pest Management on Crops in Northwestern Loss Plateau, Ministry of Agriculture, P.R. China (KFJJ20180108), the Applied Basic Research Program of Shanxi Province (201901D211354), the Excellent Doctor Introduction Award Program of Shanxi Province (SXYBKY2018016), and the Talent Support Program of Shanxi Agricultural University (2018YJ13).
Acknowledgments
We sincerely thank Thomas James Ridsdill-Smith (University of Western Australia, Perth) for reviewing an early version of this manuscript and Zhan-Feng Zhang (Northwest A & F University) for his technical assistance with mass spectrometry for measuring the amino acid levels. We also thank the assistance of all staff in the Key Laboratory of Applied Entomology, Northwest A & F University, Yangling, Shaanxi, China.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Cock, M.J.W. Bemisia tabaci–A Literature Survey on the Cotton Whitefly with an Annotated Bibliography; C.A.B International Institute of Biological Control: Ascot, UK, 1986; pp. 1–121. [Google Scholar]
- Li, S.J.; Xue, X.; Ahmed, M.Z.; Ren, S.X.; Du, Y.Z.; Wu, J.H.; Cuthbertson, A.G.S.; Qiu, B.L. Host Plants and Natural Enemies of Bemisia tabaci (Hemiptera: Aleyrodidae) in China. Insect Sci. 2011, 18, 101–120. [Google Scholar] [CrossRef]
- Barro, P.J.D.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A Statement of Species Status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Hasegawa, D.K.; Kaur, N.; Kliot, A.; Pinheiro, P.V.; Luan, J.; Stensmyr, M.C.; Zheng, Y.; Liu, W.; Sun, H.; et al. The Draft Genome of Whitefly Bemisia tabaci MEAM1, a Global Crop Pest, Provides Novel Insights into Virus Transmission, Host Adaptation, and Insecticide Resistance. BMC Biol. 2016, 14, 110. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.F.; Xia, F.; Johnson, K.W.; Bartom, E.; Tuteja, J.H.; Stevens, R.; Grossman, R.L.; Brumin, M.; White, K.P.; Ghanim, M. Genome Sequences of the Primary Endosymbiont “Candidatus Portiera aleyrodidarum” in the Whitefly Bemisia tabaci B and Q Biotypes. J. Bacteriol. 2012, 194, 6678–6679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marubayashi, J.M.; Kliot, A.; Yuki, V.A.; Rezende, J.A.M.; Krause-Sakate, R.; Pavan, M.A.; Ghanim, M. Diversity and Localization of Bacterial Endosymbionts from Whitefly Species Collected in Brazil. PLoS ONE 2014, 9, e108363. [Google Scholar] [CrossRef]
- Macfadyen, S.; Paull, C.; Boykin, L.M.; De Barro, P.; Maruthi, M.N.; Otim, M.; Kalyebi, A.; Vassão, D.G.; Sseruwagi, P.; Tay, W.T.; et al. Cassava Whitefly, Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in East African Farming Landscapes: A Review of the Factors Determining Abundance. Bull. Entomol. Res. 2018, 108, 565–582. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.X. Population Dynamics of Bemisia argentifolii (Homoptera: Aleyrodidae) on Spring Collard and Relationship to Yield in the Lower Rio Grande Valley of Texas. J. Econ. Entomol. 2000, 93, 750–756. [Google Scholar] [CrossRef]
- Wilkinson, T.L.; Adams, D.; Minto, L.B.; Douglas, A.E. The Impact of Host Plant on the Abundance and Function of Symbiotic Bacteria in an Aphid. J. Exp. Biol. 2001, 204, 3027. [Google Scholar]
- Ghosh, S.; Bouvaine, S.; Richardson, S.C.W.; Ghanim, M.; Maruthi, M.N. Fitness Costs Associated with Infections of Secondary Endosymbionts in the Cassava Whitefly Species Bemisia. tabaci. J. Pest Sci. 2018, 91, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Chiel, E.; Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Katzir, N.; Inbar, M.M. Biotype-dependent Secondary Symbiont Communities in Sympatric Populations of Bemisia tabaci. Bull. Entomol. Res. 2007, 97, 407–413. [Google Scholar] [CrossRef] [Green Version]
- Bing, X.L.; Ruan, Y.M.; Rao, Q.; Wang, X.W.; Liu, S.S. Diversity of Secondary Endosymbionts among Different Putative Species of the Whitefly Bemisia tabaci. Insect Sci. 2013, 20, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Zchori-Fein, E.; Lahav, T.; Freilich, S. Variations in the Identity and Complexity of Endosymbiont Combinations in Whitefly Hosts. Front. Microbiol. 2014, 5, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Q.; Xie, W.; Wang, S.L.; Wu, Q.J.; Liu, B.M.; Fang, Y.; Xu, B.Y.; Zhang, Y.J. The Endosymbiont Hamiltonella Increases the Growth Rate of its Host Bemisia tabaci during Periods of Nutritional Stress. PLoS ONE 2014, 9, e89002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bing, X.L.; Yang, J.; Zchori-Fein, E.; Wang, X.W.; Liu, S.S. Characterization of a Newly Discovered Symbiont of the Whitefly Bemisia tabaci (Hemiptera: Aleyrodidae). Appl. Environ. Microbiol. 2013, 79, 569–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Garcia, D.; Juravel, K.; Freilich, S.; Zchori-Fein, E.; Latorre, A.; Moya, A.; Morin, S.; Silva, F. To B or not to B: Comparative Genomics Suggests Arsenophonus as a Source of B Vitamins in Whiteflies. Front. Microbiol. 2018, 9, 2254. [Google Scholar] [CrossRef]
- Rao, Q.; Wang, S.; Su, Y.L.; Bing, X.L.; Liu, S.S.; Wang, X.W. Draft Genome Sequence of “Candidatus Hamiltonella defensa,” an Endosymbiont of the Whitefly Bemisia tabaci. J. Bacteriol. 2012, 194, 3558. [Google Scholar] [CrossRef] [Green Version]
- Luan, J.B.; Chen, W.B.; Hasegawa, D.K.; Simmons, A.M.; Wintermantel, W.M.; Ling, K.S.; Fei, Z.J.; Liu, S.S.; Douglas, A.E. Metabolic Coevolution in the Bacterial Symbiosis of Whiteflies and Related Plant Sap-Feeding Insects. Genome Biol. Evol. 2015, 7, 2635–2647. [Google Scholar] [CrossRef] [Green Version]
- Rao, Q.; Rollat-Farnier, P.A.; Zhu, D.T.; Santos-Garcia, D.; Silva, F.J.; Moya, A.; Latorre, A.; Klein, C.C.; Vavre, F.; Sagot, M.F.; et al. Genome Reduction and Potential Metabolic Complementation of the Dual Endosymbionts in the whItefly Bemisia tabaci. BMC Genom. 2015, 16, 226. [Google Scholar] [CrossRef] [Green Version]
- Ankrah, N.Y.D.; Luan, J.B.; Douglas, A.E. Cooperative Metabolism in a Three-Partner Insect-Bacterial Symbiosis Revealed by Metabolic Modeling. J. Bacteriol. 2017, 199, e00872-16. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.B.; Ren, F.R.; Yao, Y.L.; Sun, X.; Walling, L.L.; Li, N.N.; Bai, B.; Bao, X.Y.; Xu, X.R.; Luan, J.B. Intracellular Symbionts Drive Sex Ratio in the Whitefly by Facilitating Fertilization and Provisioning of B Vitamins. ISME J. 2020. [Google Scholar] [CrossRef]
- Ren, F.R.; Sun, X.; Wang, T.Y.; Yao, Y.L.; Huang, Y.Z.; Zhang, X.; Luan, J.B. Biotin Provisioning by Horizontally Transferred Genes from Bacteria Confers Animal Fitness Benefits. ISME J. 2020. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Lai, C.Y.; Kuo, M.H. Temperature Effect on the Growth of Buchnera Endosymbiont in Aphis craccivora (Hemiptera: Aphididae). Symbiosis 2009, 49, 53. [Google Scholar] [CrossRef]
- Wilkinson, T.L.; Koga, R.; Fukatsu, T. Role of Host Nutrition in Symbiont Regulation: Impact of Dietary Nitrogen on Proliferation of Obligate and Facultative Bacterial Endosymbionts of the Pea aphid Acyrthosiphon pisum. Appl. Environ. Microbiol. 2007, 73, 1362–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.H.; Kang, Z.W.; Guo, Y.; Zhu, G.S.; Shah, M.M.R.; Song, Y.; Fan, Y.L.; Jing, X.F.; Liu, T.X. Nitrogen Hurdle of Host Alternation for a Polyphagous Aphid and the Associated Changes of Endosymbionts. Sci. Rep. 2016, 6, 24781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frohlich, D.R.; Torresjerez, I.I.; Bedford, I.D.; Markham, P.G.; Brown, J.K. A Phylogeographical Analysis of the Bemisia tabaci Species Complex Based on Mitochondrial DNA Markers. Mol. Ecol. 1999, 8, 1683–1691. [Google Scholar] [CrossRef]
- Shah, M.M.R.; Liu, T.X. Feeding Experience of Bemisia tabaci (Hemiptera: Aleyrodidae) Affects their Performance on Different Host Plants. PLoS ONE 2013, 8, e77368. [Google Scholar] [CrossRef] [Green Version]
- Zchori-Fein, E.; Brown, J.K. Diversity of Prokaryotes Associated with Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). Ann. Entomol. Soc. Am. 2002, 95, 711–718. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, Y.; Ghanim, M.; Chiel, E.; Gerling, D.; Portnoy, V.; Steinberg, S.; Tzuri, G.; Horowitz, A.R.; Belausov, E.; Mozesdaube, N. Identification and Localization of a Rickettsia sp. in Bemisia tabaci (Homoptera: Aleyrodidae). Appl. Environ. Microbiol. 2006, 72, 3646–3652. [Google Scholar] [CrossRef] [Green Version]
- Heddi, A.; Grenier, A.M.; Khatchadourian, C.; Charles, H.; Nardon, P. Four Intracellular Genomes Direct Weevil Biology: Nuclear, Mitochondrial, Principal Endosymbiont, and Wolbachia. Proc. Natl. Acad. Sci. USA 1999, 96, 6814–6819. [Google Scholar] [CrossRef] [Green Version]
- Zchorifein, E.; Perlman, S.J.; Kelly, S.E.; Katzir, N.; Hunter, M.S. Characterization of a ‘Bacteroidetes’ Symbiont in Encarsia wasps (Hymenoptera: Aphelinidae): Proposal of ‘Candidatus Cardinium Hertigii’. Int. J. Syst. Evol. Microbiol. 2004, 54, 961–968. [Google Scholar] [CrossRef]
- Thao, M.L.; Baumann, P. Evolutionary Relationships of Primary Prokaryotic Endosymbionts of Whiteflies and their Hosts. Appl. Environ. Microbiol. 2004, 70, 3401–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everett, K.D.; Thao, M.; Horn, M.; Dyszynski, G.E.; Baumann, P. Novel Chlamydiae in Whiteflies and Scale Insects: Endosymbionts ‘Candidatus Fritschea bemisiae’ Strain Falk and ‘Candidatus Fritschea eriococci’ Strain Elm. Int. J. Syst. Evol. Microbiol. 2005, 55, 1581–1587. [Google Scholar] [CrossRef] [Green Version]
- Ghanim, M.; Kontsedalov, S. Susceptibility to Insecticides in the Q Biotype of Bemisia tabaci is Correlated with Bacterial Symbiont Densities. Pest Manag. Sci. 2009, 65, 939–942. [Google Scholar] [CrossRef] [PubMed]
- Brumin, M.; Kontsedalov, S.; Ghanim, M. Rickettsia Influences Thermotolerance in the Whitefly Bemisia tabaci B Biotype. Insect Sci. 2011, 18, 57–66. [Google Scholar] [CrossRef]
- Pan, H.P.; Chu, D.; Liu, B.M.; Xie, W.; Wang, S.L.; Wu, Q.J.; Xu, B.Y.; Zhang, Y.J. Relative Amount of Symbionts in Insect Hosts Changes with Host-Plant Adaptation and Insecticide Resistance. Environ. Entomol. 2013, 42, 74–78. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-∆∆Ct Method. Methods 2000, 25, 402–408. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Weibull, J.; Ronquist, F.; Brishammar, S. Free Amino Acid Composition of Leaf Exudates and Phloem Sap: A Comparative Study in Oats and Barley. Plant Physiol. 1990, 92, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. The Nutritional Quality of Phloem Sap Utilized by Natural Aphid Populations. Ecol. Entomol. 1993, 18, 31–38. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Hansen, A.K.; Moran, N.A. The impact of Microbial Symbionts on Host Plant Utilization by Herbivorous Insects. Mol. Ecol. 2014, 23, 1473–1496. [Google Scholar] [CrossRef] [PubMed]
- Shan, H.W.; Lu, Y.H.; Bing, X.L.; Liu, S.S.; Liu, Y.Q. Differential Responses of the Whitefly Bemisia tabaci Symbionts to Unfavorable Low and High Temperatures. Microb. Ecol. 2014, 68, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.A.; Moran, N.A. Horizontal Transfer of Bacterial Symbionts: Heritability and Fitness Effects in a Novel Aphid Host. Appl. Environ. Microbiol. 2005, 71, 7987–7994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, N.A.; Dunbar, H.E. Sexual Acquisition of Beneficial Symbionts in Aphids. Proc. Natl. Acad. Sci. USA 2006, 103, 12803–12806. [Google Scholar] [CrossRef] [Green Version]
- Caspi-Fluger, A.; Inbar, M.; Mozes-Daube, N.; Katzir, N.; Portnoy, V.; Belausov, E.; Hunter, M.S.; Zchori-Fein, E. Horizontal Transmission of the Insect Symbiont Rickettsia is Plant-Mediated. Proc. Biol. Sci. 2012, 279, 1791–1796. [Google Scholar] [CrossRef] [Green Version]
- Gonella, E.; Pajoro, M.; Marzorati, M.; Crotti, E.; Mandrioli, M.; Pontini, M.; Bulgari, D.; Negri, I.; Sacchi, L.; Chouaia, B.; et al. Plant-Mediated Interspecific Horizontal Transmission of an Intracellular Symbiont in Insects. Sci. Rep. 2015, 5, 15811. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.H.; Ahmed, M.Z.; Li, S.J.; Lv, N.; Shi, P.Q.; Chen, X.S.; Qiu, B.L. Plant-Mediated Horizontal Transmission of Rickettsia Endosymbiont Between Different Whitefly Species. FEMS Microbiol. Ecol. 2017, 93, fix138. [Google Scholar] [CrossRef] [Green Version]
- Wernegreen, J.J. Mutualism Meltdown in Insects: Bacteria Constrain Thermal Adaptation. Curr. Opin. Microbiol. 2012, 15, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Shan, H.W.; Deng, W.H.; Luan, J.B.; Zhang, M.J.; Zhang, Z.; Liu, S.S.; Liu, Y.Q. Thermal Sensitivity of Bacteriocytes Constrains the Persistence of Intracellular Bacteria in Whitefly Symbiosis under Heat Stress. Environ. Microbiol. Rep. 2017, 9, 706–716. [Google Scholar] [CrossRef]
- Lemoine, M.M.; Engl, T.; Kaltenpoth, M. Microbial Symbionts Expanding or Constraining Abiotic Niche Space in Insects. Curr. Opin. Insect Sci. 2020, 39, 14–20. [Google Scholar] [CrossRef]
- Pan, H.P.; Su, Q.; Jiao, X.G.; Zhou, L.; Liu, B.M.; Xie, W.; Wang, S.L.; Wu, Q.J.; Xu, B.Y.; Zhang, Y.J. Relative Amount of Symbionts in Bemisia tabaci (Gennadius) Q Changes with Host Plant and Establishing the Method of Analyzing Free Amino Acid in B. tabaci. Commun. Integr. Biol. 2013, 6, e23397. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Cao, W.J.; Zhong, L.R.; Godfray, H.C.J.; Liu, X.D. Host Plant Determines the Population Size of an Obligate Symbiont (Buchnera aphidicola) in Aphids. Appl. Environ. Microbiol. 2016, 82, 2336–2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, T.L.; Douglas, A.E. Phloem Amino Acids and the Host Plant Range of the Polyphagous Aphid, Aphis. fabae. Entomol. Exp. Appl. 2003, 106, 103–113. [Google Scholar] [CrossRef]
- Kakimoto, K.; Inoue, H.; Yamaguchi, T.; Ueda, S.; Honda, K.i.; Yano, E. Host Plant Effect on Development and Reproduction of Bemisia argentifolii Bellows et Perring (B. tabaci [Gennadius] B-biotype) (Homoptera: Aleyrodidae). Appl. Entomol. Zool. 2007, 42, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Tsai, J.H.; Wang, K.H. Development and Reproduction of Bemisia argentifolii (Homoptera: Aleyrodidae) on Five Host Plants. Environ. Entomol. 1996, 25, 810–816. [Google Scholar] [CrossRef]
- Chandler, S.M.; Wilkinson, T.L.; Douglas, A.E. Impact of Plant Nutrients on the Relationship between a Herbivorous Insect and its Symbiotic Bacteria. Proc. Biol. Sci. 2008, 275, 565–570. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E.; Minto, L.B.; Wilkinson, T.L. Quantifying Nutrient Production by the Microbial Symbionts in an Aphid. J. Exp. Biol. 2001, 204, 349–358. [Google Scholar]
- Chiel, E.; Inbar, M.; Mozes-Daube, N.; White, J.A.; Hunter, M.S.; Zchori-Fein, E. Assessments of Fitness Effects by the Facultative Symbiont Rickettsia in the Sweetpotato Whitefly (Hemiptera: Aleyrodidae). Ann. Entomol. Soc. Am. 2009, 102, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Himler, A.G.; Adachi-Hagimori, T.; Bergen, J.E.; Kozuch, A.; Kelly, S.E.; Tabashnik, B.E.; Chiel, E.; Duckworth, V.E.; Dennehy, T.J.; Zchori-Fein, E. Rapid Spread of a Bacterial Symbiont in an Invasive Whitefly is Driven by Fitness Benefits and Female Bias. Science 2011, 332, 254–256. [Google Scholar] [CrossRef] [Green Version]
- Harborne, J.B. Introduction to Ecological Biochemistry, 4th ed.; Academic Press: London, UK, 1993; pp. 128–158. [Google Scholar]
Figure 1.
Schematic diagram of the plant switch experiments designed in this study. MEAM1: Middle East-Asia Minor 1.
Figure 1.
Schematic diagram of the plant switch experiments designed in this study. MEAM1: Middle East-Asia Minor 1.

Figure 2.
Relative abundance of Portiera (A), Hamiltonella (B), and Rickettsia (C) (mean ± SE) in whitefly populations G0 of Bemisia-eggplants, Bemisia-cucumbers, and Bemisia-tomatoes. The relative amount of each symbiont taxon was normalized using β-actin and calculated by the 2-ΔCt method. Statistical analyses were carried out separately for each symbiont taxon; different letters above the error bars indicate significant differences for that taxon among the three whitefly populations (p < 0.05, Tukey’s honest significant difference (HSD) test).
Figure 2.
Relative abundance of Portiera (A), Hamiltonella (B), and Rickettsia (C) (mean ± SE) in whitefly populations G0 of Bemisia-eggplants, Bemisia-cucumbers, and Bemisia-tomatoes. The relative amount of each symbiont taxon was normalized using β-actin and calculated by the 2-ΔCt method. Statistical analyses were carried out separately for each symbiont taxon; different letters above the error bars indicate significant differences for that taxon among the three whitefly populations (p < 0.05, Tukey’s honest significant difference (HSD) test).

Figure 3.
Relative amounts of Portiera (mean ± SE) in whitefly populations of Bemisia-eggplants (A), Bemisia-cucumbers (B), and Bemisia-tomatoes (C) transferred to natal and novel host-plant species for different generations, ranging from G0 to G5. The relative amount of each symbiont taxon was normalized using β-actin and calculated by the 2-ΔCt method. Different lowercase letters and lowercase letter variants, i.e., a′, b’ and c′, around the error bars indicate significant differences among different generations after a transfer to a novel host-plant species (p < 0.05, Tukey’s HSD test).
Figure 3.
Relative amounts of Portiera (mean ± SE) in whitefly populations of Bemisia-eggplants (A), Bemisia-cucumbers (B), and Bemisia-tomatoes (C) transferred to natal and novel host-plant species for different generations, ranging from G0 to G5. The relative amount of each symbiont taxon was normalized using β-actin and calculated by the 2-ΔCt method. Different lowercase letters and lowercase letter variants, i.e., a′, b’ and c′, around the error bars indicate significant differences among different generations after a transfer to a novel host-plant species (p < 0.05, Tukey’s HSD test).

Figure 4.
Relative amounts of Hamiltonella and Rickettsia (mean ± SE) in whitefly populations of Bemisia-eggplants (A,D), Bemisia-cucumbers (B,E), and Bemisia-tomatoes (C,F) transferred to natal and two novel host-plant species for one generation. The relative amounts of each symbiont taxon were normalized using β-actin and calculated by the 2-ΔCt method. Different lowercase letters around the error bars of each whitefly population indicate significant difference of Hamiltonella or Rickettsia when transferred to different plant species (p < 0.05, Tukey’s HSD test).
Figure 4.
Relative amounts of Hamiltonella and Rickettsia (mean ± SE) in whitefly populations of Bemisia-eggplants (A,D), Bemisia-cucumbers (B,E), and Bemisia-tomatoes (C,F) transferred to natal and two novel host-plant species for one generation. The relative amounts of each symbiont taxon were normalized using β-actin and calculated by the 2-ΔCt method. Different lowercase letters around the error bars of each whitefly population indicate significant difference of Hamiltonella or Rickettsia when transferred to different plant species (p < 0.05, Tukey’s HSD test).

Figure 5.
Proportions of individual amino acids (mean ± SE) (mol%; A), amino acid–composition analysis by principle component analysis (PCA) (B), and percentage of the entire essential amino acids (EAAs) in the phloem sap of eggplant, cucumber, and tomato leaves (mol%; C). Non-EAAs: Ala, alanine; Asp, aspartic acid; Cys, cysteine; Gln, glutamine; Glu, glutamic acid; Pro, proline; Ser, serine; and Tyr, tyrosine. EAAs: Arg, arginine; His, histidine; Ile, isoleucine; Leu, leucine; Lys, lysine; Met, methionine; Phe, phenylalanine; Thr, threonine; Trp, tryptophan; and Val, valine. Asterisk or different letters above the error bars indicate significant differences in individual amino acids or percentage of the entire EAAs among the three plant species (p < 0.05, Tukey’s HSD test).
Figure 5.
Proportions of individual amino acids (mean ± SE) (mol%; A), amino acid–composition analysis by principle component analysis (PCA) (B), and percentage of the entire essential amino acids (EAAs) in the phloem sap of eggplant, cucumber, and tomato leaves (mol%; C). Non-EAAs: Ala, alanine; Asp, aspartic acid; Cys, cysteine; Gln, glutamine; Glu, glutamic acid; Pro, proline; Ser, serine; and Tyr, tyrosine. EAAs: Arg, arginine; His, histidine; Ile, isoleucine; Leu, leucine; Lys, lysine; Met, methionine; Phe, phenylalanine; Thr, threonine; Trp, tryptophan; and Val, valine. Asterisk or different letters above the error bars indicate significant differences in individual amino acids or percentage of the entire EAAs among the three plant species (p < 0.05, Tukey’s HSD test).

Figure 6.
Proportions of individual amino acids (mean ± SE; mol%; A), amino acid composition–profile analysis by PCA (B), and percentage of the entire EAAs in three whitefly populations (G0), termed Bemisia-eggplants, Bemisia-cucumbers, and Bemisia-tomatoes (mol%) (C). Asterisks or different letters above the error bars indicate significant differences in individual amino-acid levels or percentage of the entire EAAs among the whitefly populations (p < 0.05, Tukey’s HSD test).
Figure 6.
Proportions of individual amino acids (mean ± SE; mol%; A), amino acid composition–profile analysis by PCA (B), and percentage of the entire EAAs in three whitefly populations (G0), termed Bemisia-eggplants, Bemisia-cucumbers, and Bemisia-tomatoes (mol%) (C). Asterisks or different letters above the error bars indicate significant differences in individual amino-acid levels or percentage of the entire EAAs among the whitefly populations (p < 0.05, Tukey’s HSD test).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers used for symbiont identification and quantification.
| Organism and Target Gene | Primer Sequences (5′ to 3′) | Product Length (bp) | References |
|---|---|---|---|
| Diagnostic PCR | |||
| Bemisia tabaci mtCOI | C1-J-2195: TTGATTTTTTGGTCATCCAGAAGT L2-N-3014: TCCAATGCACTAATCTGCCATATTA | 800 | [26] |
| Portiera 16S rRNA | Por-F: TGCAAGTCGAGCGGCATCAT Por-R: AAAGTTCCCGCCTTATGCGT | 1000 | [28] |
| Hamiltonella 16S rRNA | Ham-F: TGAGTAAAGTCTGGGAATCTGG Ham-R: CCCGGGAACGTATTCACCGTAG | 1000 | [28] |
| Rickettsia 16S rRNA | Rb-F: GCTCAGAACGAACGCTATC Rb-R: GAAGGAAAGCATCTCTGC | 900 | [29] |
| Wolbachia 16S rRNA | Wol-16S-F: CGGGGGAAAAATTTATTGCT Wol-16S-R: AGCTGTAATACAGAAAGTAAA | 700 | [11,30] |
| Cardinium 16S rRNA | Ch-F: TACTGTAAGAATAAGCACCGGC Ch-R: GTGGATCACTTAACGCTTTCG | 400 | [31] |
| Hemipteriphilus 16S rRNA | Hem-F: GCTCAGAACGAACGCTRKC Hem-R: TTCGCCACTGGTGTTCCTC | 670 | [15] |
| Arsenophonus 23S rRNA | Ars23S-1: CGTTTGATGAATTCATAGTCAAA Ars23S-2: GGTCCTCCAGTTAGTGTTACCCAAC | 900 | [32] |
| Fritschea 23S rRNA | U23F: GATGCCTTGGCATTGATAGGCGATGAAGGA 23SIGR: TGGCTCATCATGCAAAAGGCA | 600 | [11,33] |
| qPCR | |||
| B. tabaci β-actin | qActinF: TCTTCCAGCCATCCTTCTTG qActinR: CGGTGATTT CCTTCTGCATT | 130 | [34,35] |
| Portiera 16S rRNA | qPor-F: TAGTCCACGCTGTAAACG qPor-R: AGGCACCCTTCCATCT | 229 | [36] |
| Hamiltonella 16S rRNA | qHam-F: GCATCGAGTGAGCACAGTTT qHam-R: TATCCTCTCAGACCCGCTAGA | 243 | [35,36] |
| Rickettsia gltA | qgltA-F: AAAGGTTGCTCATCATGCGTT qgltA-R: GCCATAGGATGCGAAGAGCT | 80 | [34,35] |
Table 2.
Effects of the B. tabaci population, host plant, and generation number since host-plant-shifting on Portiera abundance, determined by three-way ANOVA. df: degrees of freedom.
Table 2.
Effects of the B. tabaci population, host plant, and generation number since host-plant-shifting on Portiera abundance, determined by three-way ANOVA. df: degrees of freedom.
| Effect | Portiera Amount | |||
|---|---|---|---|---|
| df | Mean Square | F | p | |
| Population (A) | 2 | 2713.206 | 943.388 | <0.001 |
| Host plant (B) | 2 | 5202.316 | 1808.857 | <0.001 |
| Generation (C) | 4 | 77.291 | 26.874 | <0.001 |
| A × B | 4 | 103.224 | 35.891 | <0.001 |
| A × C | 8 | 78.090 | 27.152 | <0.001 |
| B × C | 8 | 75.856 | 26.375 | <0.001 |
| A × B × C | 16 | 62.033 | 21.569 | <0.001 |
Table 3.
Effects of the B. tabaci population and host plant since host-plant-shifting on Hamiltonella and Rickettsia abundance, determined by two-way ANOVA.
Table 3.
Effects of the B. tabaci population and host plant since host-plant-shifting on Hamiltonella and Rickettsia abundance, determined by two-way ANOVA.
| Effect | Hamiltonella Amount | Rickettsia Amount | ||||||
|---|---|---|---|---|---|---|---|---|
| df | Mean Square | F | p | df | Mean Square | F | p | |
| Population (A) | 2 | 29.861 | 28.377 | <0.001 | 2 | 16.368 | 11.706 | <0.001 |
| Host plant (B) | 2 | 3.084 | 2.931 | 0.064 | 2 | 8.526 | 6.098 | 0.005 |
| A × B | 4 | 12.951 | 12.307 | <0.001 | 4 | 18.938 | 13.544 | <0.001 |
Table 4.
Statistical analyses of individual amino acids detected from plant phloem sap and from whitefly populations.
Table 4.
Statistical analyses of individual amino acids detected from plant phloem sap and from whitefly populations.
| Amino Acid | Plant Leaves | Whitefly | ||
|---|---|---|---|---|
| F2,15 Value | p-Value | F2,24 Value | p-Value | |
| Ala | 30.19 | <0.001 | 12.08 | <0.001 |
| Asp | 23.10 | <0.001 | undetected | |
| Cys | 9.93 | 0.002 | 0.45 | 0.642 |
| Gln | 2.61 | 0.107 | undetected | |
| Glu | 30.52 | <0.001 | 12.85 | <0.001 |
| Gly | undetected | 1.61 | 0.216 | |
| Pro | 9.20 | 0.003 | 6.26 | 0.005 |
| Ser | 27.04 | <0.001 | 32.64 | <0.001 |
| Tyr | 16.12 | <0.001 | 8.32 | 0.001 |
| Arg | 1.40 | 0.277 | 10.88 | <0.001 |
| His | 1.11 | 0.354 | 12.21 | <0.001 |
| Ile | 20.62 | <0.001 | 1.25 | 0.299 |
| Leu | 19.76 | <0.001 | 3.73 | 0.035 |
| Lys | 3.40 | 0.061 | 85.69 | <0.001 |
| Met | 35.09 | <0.001 | 0.44 | 0.646 |
| Phe | 68.71 | <0.001 | 0.02 | 0.984 |
| Thr | 80.39 | <0.001 | 0.54 | 0.588 |
| Trp | 13.22 | <0.001 | 0.12 | 0.890 |
| Val | 2.65 | 0.104 | 1.19 | 0.316 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, Y.-H.; Shah, M.M.R.; Song, Y.; Liu, T.-X. Host Plant Affects Symbiont Abundance in Bemisia tabaci (Hemiptera: Aleyrodidae). Insects 2020, 11, 501. https://doi.org/10.3390/insects11080501
AMA Style
Liu Y-H, Shah MMR, Song Y, Liu T-X. Host Plant Affects Symbiont Abundance in Bemisia tabaci (Hemiptera: Aleyrodidae). Insects. 2020; 11(8):501. https://doi.org/10.3390/insects11080501
Chicago/Turabian StyleLiu, Yan-Hong, M. Mostafizur Rahman Shah, Yue Song, and Tong-Xian Liu. 2020. "Host Plant Affects Symbiont Abundance in Bemisia tabaci (Hemiptera: Aleyrodidae)" Insects 11, no. 8: 501. https://doi.org/10.3390/insects11080501
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.