Stingless Bee Honey: Evaluating Its Antibacterial Activity and Bacterial Diversity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling of Honey
2.2. Antimicrobial Test
2.3. DNA Extraction
2.4. PCR Amplification of 16S rRNA, Library Preparation and Sequencing
2.5. Sequence Data Analysis
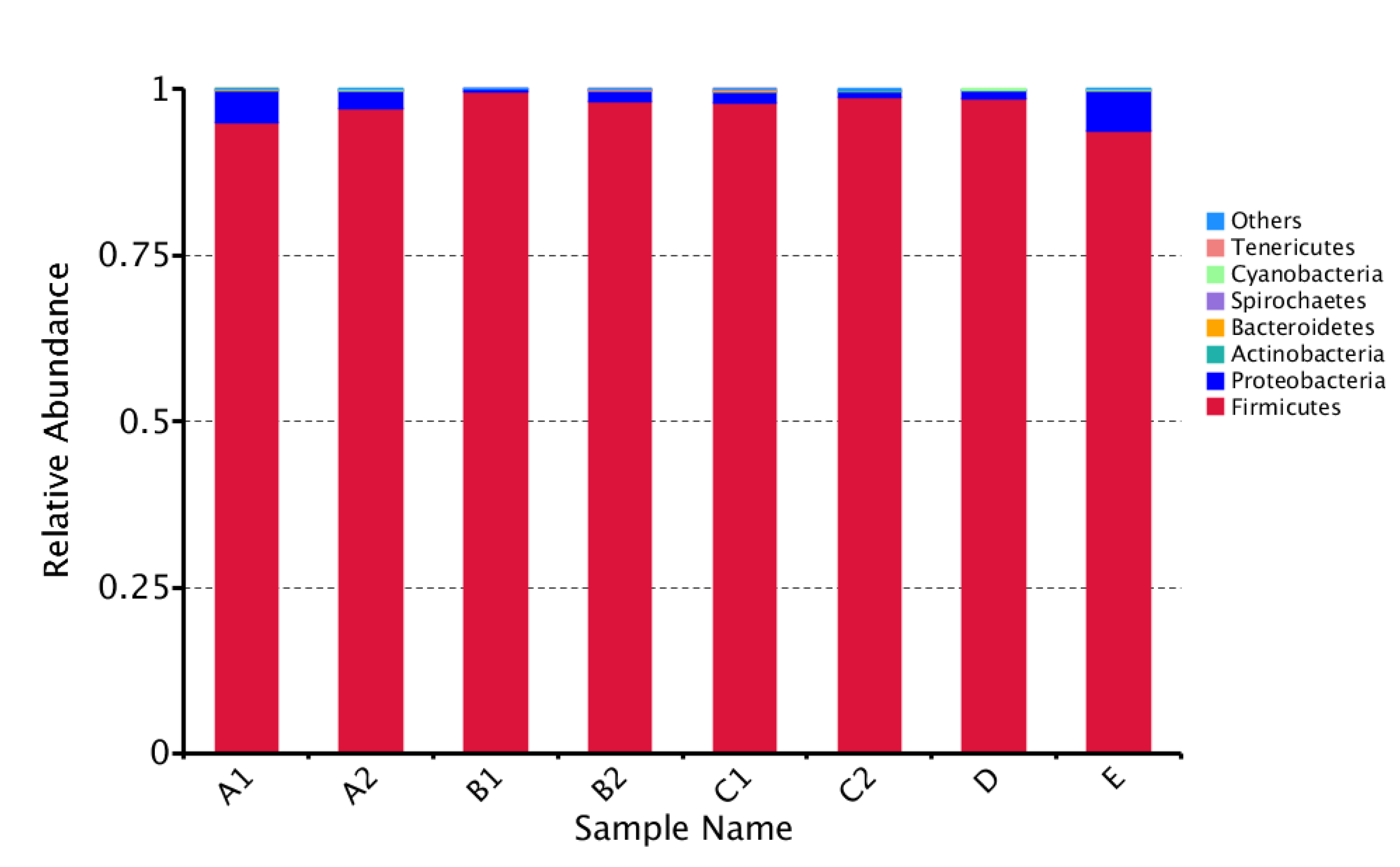
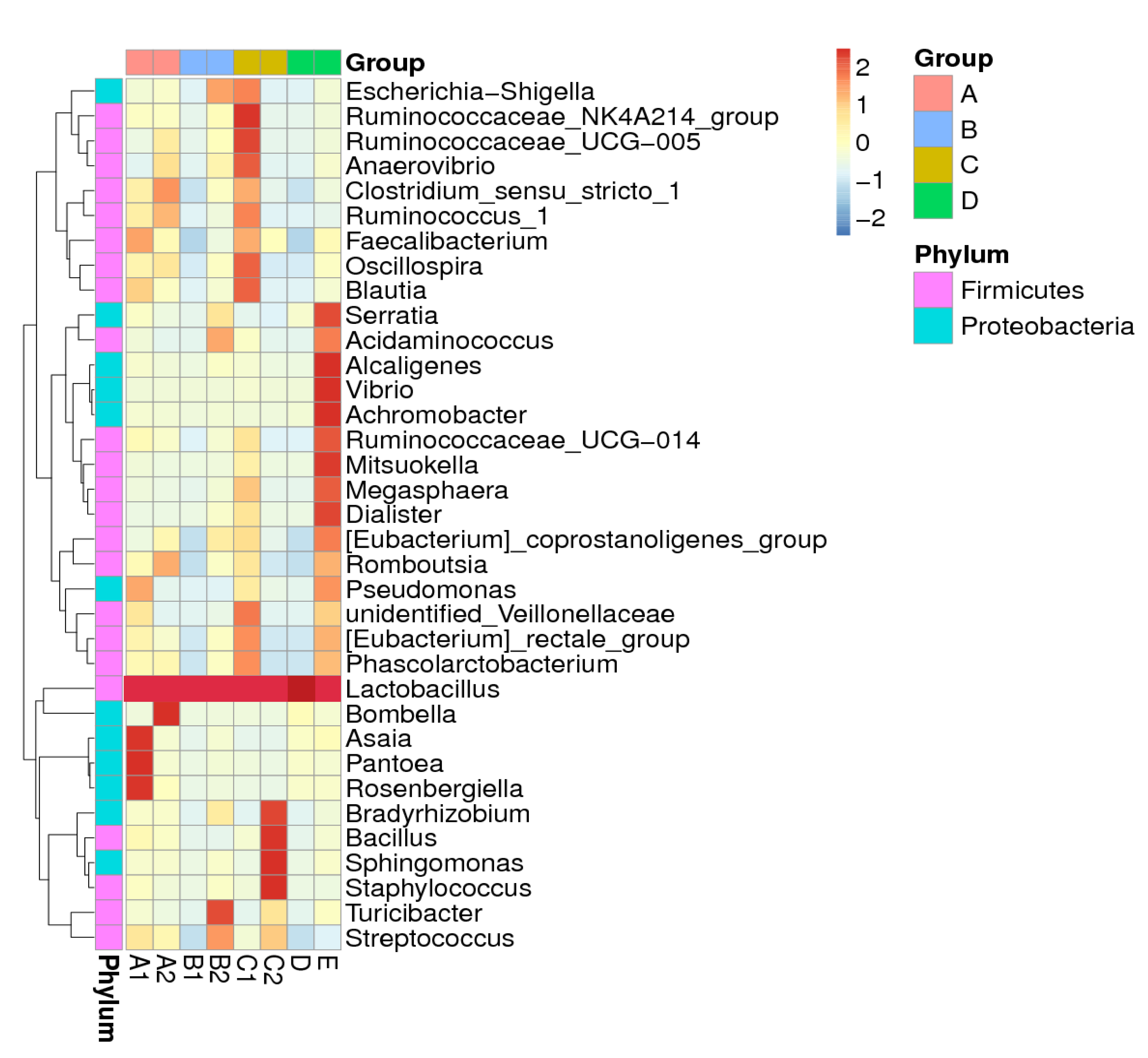
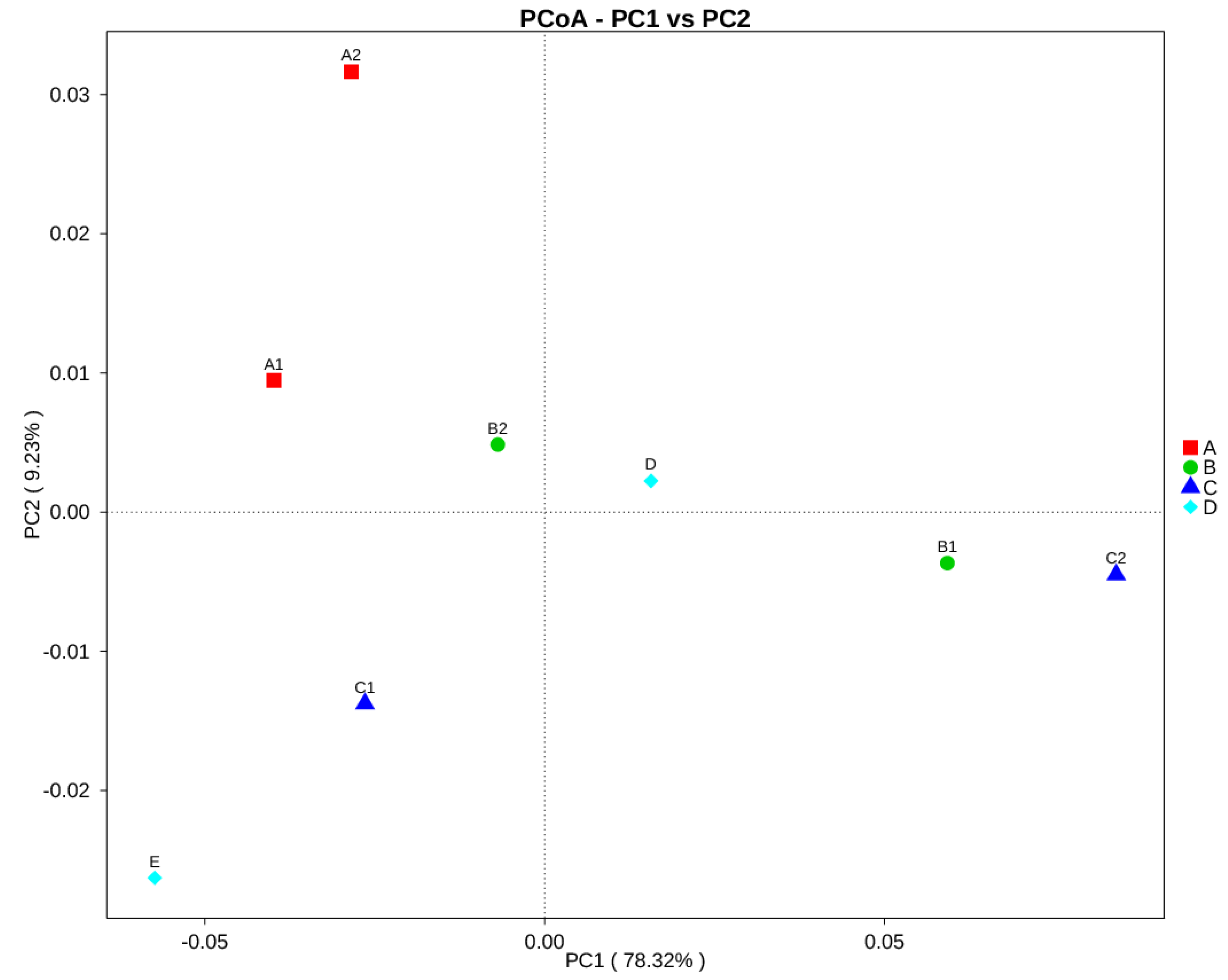
3. Results
3.1. Antimicrobial Activity
3.2. Sequencing Information and Bacterial Diversity of Stingless Bee Honey
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Vit, P.; Pedro, S.R.; Roubik, D. Pot-Honey: A Legacy of Stingless Bees; Springer: New York, NY, USA, 2013. [Google Scholar]
- Roubik, D.W. Stingless bee nesting biology. Apidologie 2006, 37, 124–143. [Google Scholar] [CrossRef] [Green Version]
- Amin, Z.; Aina, F.; Sabri, S.; Mohammad, S.M.; Ismail, M.; Chan, K.W.; Ismail, N.; Norhaizan, M.E.; Zawawi, N. Therapeutic properties of stingless bee honey in comparison with European bee honey. Adv. Pharm. Sci. 2018, 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.V.; Krishnan, K.T.; Salleh, N.; Gan, S.H. Biological and therapeutic effects of honey produced by honey bees and stingless bees: A comparative review. Rev. Bras. Farm. 2016, 26, 657–664. [Google Scholar] [CrossRef] [Green Version]
- Biesmeijer, J.C.; Slaa, E.J. Information flow and organization of stingless bee foraging. Apidologie 2004, 35, 143–157. [Google Scholar] [CrossRef] [Green Version]
- Souza, B.A.; Roubik, D.W.; Barth, O.M.; Heard, T.A.; Enríquez, E.; Carvalho, C.; Villas-Bôas, J.; Marchini, L.; Locatelli, J.; Persano-Oddo, L. Composition of stingless bee honey: Setting quality standards. Interciencia 2006, 31, 867–875. [Google Scholar]
- Tanzi, M.G.; Gabay, M.P. Association between honey consumption and infant botulism. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2002, 22, 1479–1483. [Google Scholar] [CrossRef]
- Reyes-González, A.; Camou-Guerrero, A.; Gómez-Arreola, S. From extraction to meliponiculture: A case study of the management of stingless bees in the West-central region of Mexico. In Beekeeping and Bee Conservation; Chambó, E., Ed.; InTechOpen: Rijeka, Croatia, 2016; pp. 201–223. [Google Scholar]
- Lim, S.-M.; Im, D.-S. Screening and characterization of probiotic lactic acid bacteria isolated from Korean fermented foods. J. Microbiol. Biotechnol. 2009, 19, 178–186. [Google Scholar] [CrossRef] [Green Version]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef]
- Lee, H.; Churey, J.J.; Worobo, R.W. Antimicrobial activity of bacterial isolates from different floral sources of honey. Int. J. Food Microbiol. 2008, 126, 240–244. [Google Scholar] [CrossRef]
- Casalone, E.; Cavalieri, D.; Daly, G.; Vitali, F.; Perito, B. Propolis hosts a diverse microbial community. World J. Microbiol. Biotechnol. 2020, 36, 50. [Google Scholar] [CrossRef]
- Silva, M.S.; Rabadzhiev, Y.; Eller, M.R.; Iliev, I.; Ivanova, I.; Santana, W.C. Microorganisms in Honey. In Honey Analysis; de Toledo, V.A.A., Ed.; InTechOpen: Rijeka, Croatia, 2017; pp. 232–245. [Google Scholar]
- Sabree, Z.L.; Hansen, A.K.; Moran, N.A. Independent studies using deep sequencing resolve the same set of core bacterial species dominating gut communities of honey bees. PLoS ONE 2012, 7, e41250. [Google Scholar] [CrossRef] [Green Version]
- Alnaimat, S.; Wainwright, M.; Al’Abri, K. Antibacterial potential of honey from different origins: A comparsion with manuka honey. J. Microbiol. Biotechnol. Food Sci. 2012, 1, 1328–1338. [Google Scholar]
- Ewnetu, Y.; Lemma, W.; Birhane, N. Antibacterial effects of Apis mellifera and stingless bees honeys on susceptible and resistant strains of Escherichia coli, Staphylococcus aureus and Klebsiella pneumoniae in Gondar, Northwest Ethiopia. BMC Complement. Altern. Med. 2013, 13, 269. [Google Scholar] [CrossRef] [Green Version]
- Israili, Z.H. Antimicrobial properties of honey. Am. J. Ther. 2014, 21, 304–323. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, D. Characterization of Lactobacillus isolated from dairy samples for probiotic properties. Anaerobe 2015, 33, 117–123. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Cullings, K. Simplified Doyle and Doyle extraction procedure. Mol. Ecol. 1992, 1, 233–240. [Google Scholar] [CrossRef]
- Smith, D.P.; Peay, K.G. Sequence depth, not PCR replication, improves ecological inference from next generation DNA sequencing. PLoS ONE 2014, 9, e90234. [Google Scholar] [CrossRef] [Green Version]
- Menezes, C.; Vollet-Neto, A.; Contrera, F.A.F.L.; Venturieri, G.C.; Imperatriz-Fonseca, V.L. The Role of Useful Microorganisms to Stingless Bees and Stingless Beekeeping; Springer: New York, NY, USA, 2013; pp. 153–171. [Google Scholar]
- Lee, I.C.; Tomita, S.; Kleerebezem, M.; Bron, P.A. The quest for probiotic effector molecules—Unraveling strain specificity at the molecular level. Pharmacol. Res. 2013, 69, 61–74. [Google Scholar] [CrossRef]
- Maldonado-Barragán, A.; Caballero-Guerrero, B.; Lucena-Padrós, H.; Ruiz-Barba, J.L. Induction of bacteriocin production by coculture is widespread among plantaricin-producing Lactobacillus plantarum strains with different regulatory operons. Food Microbiol. 2013, 33, 40–47. [Google Scholar] [CrossRef]
- Collins, F.W.; Mesa-Pereira, B.; O’Connor, P.M.; Rea, M.C.; Hill, C.; Ross, R.P. Reincarnation of bacteriocins from the Lactobacillus pangenomic graveyard. Front. Microbiol. 2018, 9, 1298. [Google Scholar] [CrossRef]
- Ranjan, R.; Rani, A.; Metwally, A.; McGee, H.S.; Perkins, D.L. Analysis of the microbiome: Advantages of whole genome shotgun versus 16S amplicon sequencing. Biochem. Biophys. Res. Commun. 2016, 469, 967–977. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.; Jacobus, N.; Deneke, C.; Gorbach, S. Antimicrobial substance from a human Lactobacillus strain. Antimicrob. Agents Chemother. 1987, 31, 1231–1233. [Google Scholar] [CrossRef] [Green Version]
- Chuttong, B.; Chanbang, Y.; Sringarm, K.; Burgett, M. Physicochemical profiles of stingless bee (Apidae: Meliponini) honey from South east Asia (Thailand). Food Chem. 2016, 192, 149–155. [Google Scholar] [CrossRef]
- Bakar, M.A.; Sanusi, S.B.; Bakar, F.A.; Cong, O.J.; Mian, Z. Physicochemical and antioxidant potential of raw unprocessed honey from Malaysian stingless bees. Pak. J. Nutr. 2017, 16, 888–894. [Google Scholar] [CrossRef] [Green Version]
- Gilliam, M.; Buchmann, S.L.; Lorenz, B.J.; Roubik, D.W. Microbiology of the larval provisions of the stingless bee, Trigona hypogea, an obligate necrophage. Biotropica 1985, 71, 28–31. [Google Scholar] [CrossRef]
- Gilliam, M.; Roubik, D.W.; Lorenz, B.J. Microorganisms associated with pollen, honey, and brood provisions in the nest of a stingless bee, Melipona fasciata. Apidologie 1990, 21, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Zulkhairi, A.F.; Sabri, S.; Ismail, M.; Chan, K.W.; Ismail, N.; Mohd, E.N.; Mohd, L.M.; Zawawi, N. Probiotic Properties of Bacillus Strains Isolated from Stingless Bee (Heterotrigona itama) Honey Collected across Malaysia. Int. J. Environ. Res. Public Health 2019, 17, 278. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, T.C.; Alsterfjord, M.; Nilson, B.; Butler, È.; Vásquez, A. Lactobacillus apinorum sp. nov., Lactobacillus mellifer sp. nov., Lactobacillus mellis sp. nov., Lactobacillus melliventris sp. nov., Lactobacillus kimbladii sp. nov., Lactobacillus helsingborgensis sp. nov. and Lactobacillus kullabergensis sp. nov., isolated from the honey stomach of the honeybee Apis mellifera. Int. J. Syst. Evol. Microbiol. 2014, 64, 3109–3119. [Google Scholar]
- Tajabadi, N.; Mardan, M.; Manap, M.Y.A.; Shuhaimi, M.; Meimandipour, A.; Nateghi, L. Detection and identification of Lactobacillus bacteria found in the honey stomach of the giant honeybee Apis dorsata. Apidologie 2011, 42, 642. [Google Scholar] [CrossRef]
- Killer, J.; Kopečný, J.; Mrázek, J.; Rada, V.; Dubná, S.; Marounek, M. Bifidobacteria in the digestive tract of bumblebees. Anaerobe 2010, 16, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Avila, S.; Beux, M.R.; Ribani, R.H.; Zambiazi, R.C. Stingless bee honey: Quality parameters, bioactive compounds, health-promotion properties and modification detection strategies. Trends Food Sci. Technol. 2018, 81, 37–50. [Google Scholar] [CrossRef]
- G-Alegría, E.; López, I.; Ruiz, J.I.; Sáenz, J.; Fernández, E.; Zarazaga, M.; Dizy, M.; Torres, C.; Ruiz-Larrea, F. High tolerance of wild Lactobacillus plantarum and Oenococcus oeni strains to lyophilisation and stress environmental conditions of acid pH and ethanol. FEMS Microbiol. Lett. 2004, 230, 53–61. [Google Scholar] [CrossRef]
- Basari, N.; Ramli, S.; Mohd Khairi, N. Food reward and distance influence the foraging pattern of stingless bee, Heterotrigona itama. Insects 2018, 9, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olofsson, T.C.; Vásquez, A. Detection and identification of a novel lactic acid bacterial flora within the honey stomach of the honeybee Apis mellifera. Curr. Microbiol. 2008, 57, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Horton, M.A.; Oliver, R.; Newton, I.L. No apparent correlation between honey bee forager gut microbiota and honey production. PeerJ 2015, 3, e1329. [Google Scholar] [CrossRef] [Green Version]
- Moran, N.A. Genomics of the honey bee microbiome. Curr. Opin. Insect Sci. 2015, 10, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Vasquez, A.; Olofsson, T.C. The lactic acid bacteria involved in the production of bee pollen and bee bread. J. Apic. Res. 2009, 48, 189–195. [Google Scholar] [CrossRef]
- Russell, C.; Walker, T. Lactobacillus malefermentans n. sp., Isolated from beer. Microbiology 1953, 8, 160–162. [Google Scholar]
- Oberg, C.J.; Oberg, T.S.; Culumber, M.D.; Ortakci, F.; Broadbent, J.R.; McMahon, D.J. Lactobacillus wasatchensis sp. nov., a non-starter lactic acid bacteria isolated from aged cheddar cheese. Int. J. Syst. Evol. Microbiol. 2016, 66, 158–164. [Google Scholar] [CrossRef]
- Ortakci, F.; Broadbent, J.R.; Oberg, C.J.; McMahon, D.J. Late blowing of Cheddar cheese induced by accelerated ripening and ribose and galactose supplementation in presence of a novel obligatory heterofermentative nonstarter Lactobacillus wasatchensis. J. Dairy Sci. 2015, 98, 7460–7472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isobe, H.; Nishiyama, A.; Takano, T.; Higuchi, W.; Nakagawa, S.; Taneike, I.; Fukushima, Y.; Yamamoto, T. Reduction of overall Helicobacter pylori colonization levels in the stomach of Mongolian gerbil by Lactobacillus johnsonii La1 (LC1) and its in vitro activities against H. pylori motility and adherence. Biosci. Biotechnol. Biochem. 2012, 76, 850–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davoren, M.J.; Liu, J.; Castellanos, J.; Rodríguez-Malavé, N.I.; Schiestl, R.H. A novel probiotic, Lactobacillus johnsonii 456, resists acid and can persist in the human gut beyond the initial ingestion period. Gut Microbes 2019, 10, 458–480. [Google Scholar] [CrossRef] [PubMed]
- Nath, A.H. Development and Quality Evaluation of Probiotic Honey Beverage. Ph.D. Thesis, College of Horticulture, Vellayani, India, 2015. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Stingless Bee Honey Samples of Different Species & Concentration | Inhibition Zone Measurement Against Different Bacteria (mm) | |||||
|---|---|---|---|---|---|---|
| pH | S. aureus | A. faecalis | S. marcescens | B. subtilis | E. coli | |
| H. fimbriata | 2.64 ± 0.17 | |||||
| 50% | 28 ± 0.58 | 21 ± 1.00 | 18 ± 1.00 | 9 ± 3.72 | 12 ± 0.58 | |
| 25% | 19 ± 0.58 | 15 ± 0.58 | 12 ± 0.58 | 0 ± 2.40 | 11 ± 0.00 | |
| 12.5% | 16 ± 0.58 | 0 ± 0.00 | 11 ± 0.00 | 0 ± 8.59 | 9 ± 0.58 | |
| T. melanoleuca | 2.89 ± 0.20 | |||||
| 50% | 22 ± 0.58 | 21 ± 1.00 | 14 ± 0.58 | 9 ± 0.82 | 9 ± 1.00 | |
| 25% | 18 ± 0.58 | 0 ± 0.00 | 11 ± 1.53 | 11 ± 9.88 | 0 ± 0.00 | |
| 12.5% | 0 ± 0.00 | 0 ± 0.00 | 10 ± 1.53 | 0 ± 0.00 | 0 ± 0.00 | |
| H. itama | 2.57 ± 0.19 | |||||
| 50% | 13 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 12 ± 0.00 | 0 ± 0.00 | |
| 25% | 11 ± 0.58 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| 12.5% | 10 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| T. apicalis | 2.20 ± 0.21 | |||||
| 50% | 13 ± 0.58 | 0 ± 0.00 | 0 ± 0.00 | 12 ± 0.00 | 0 ± 0.00 | |
| 25% | 9 ± 1.53 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| 12.5% | 8 ± 0.58 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| L. terminata | 2.36 ± 0.20 | |||||
| 50% | 11 ± 0.58 | 0 ± 0.00 | 0 ± 0.00 | 12 ± 1.53 | 0 ± 0.00 | |
| 25% | 10 ± 0.58 | 0 ± 0.00 | 0 ± 0.00 | 9 ± 0.58 | 0 ± 0.00 | |
| 12.5% | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| G. thoracica | 1.95 ± 0.13 | |||||
| 50% | 11 ± 1.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| 25% | 9 ± 1.53 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| 12.5% | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| T. binghami | 2.19 ± 0.19 | |||||
| 50% | 10 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| 25% | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| 12.5% | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| H. erythrogastra | 1.82 ± 0.l2 | |||||
| 50% | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 9 ± 1.00 | 0 ± 0.00 | |
| 25% | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| 12.5% | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | |
| Sample Name | Number of Bacterial OTU | Shannon Index (H) | Simpson Index (D) |
|---|---|---|---|
| A2 | 158 | 2.18 | 0.53 |
| C1 | 205 | 2.10 | 0.47 |
| A1 | 131 | 2.00 | 0.47 |
| E | 149 | 1.96 | 0.42 |
| C2 | 51 | 1.60 | 0.56 |
| B2 | 106 | 1.18 | 0.25 |
| B1 | 20 | 1.76 | 0.68 |
| D | 28 | 0.85 | 0.22 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosli, F.N.; Hazemi, M.H.F.; Akbar, M.A.; Basir, S.; Kassim, H.; Bunawan, H. Stingless Bee Honey: Evaluating Its Antibacterial Activity and Bacterial Diversity. Insects 2020, 11, 500. https://doi.org/10.3390/insects11080500
Rosli FN, Hazemi MHF, Akbar MA, Basir S, Kassim H, Bunawan H. Stingless Bee Honey: Evaluating Its Antibacterial Activity and Bacterial Diversity. Insects. 2020; 11(8):500. https://doi.org/10.3390/insects11080500
Chicago/Turabian StyleRosli, Farah Nadiah, Mohd Hafiz Fikri Hazemi, Muhamad Afiq Akbar, Syazwani Basir, Hakimi Kassim, and Hamidun Bunawan. 2020. "Stingless Bee Honey: Evaluating Its Antibacterial Activity and Bacterial Diversity" Insects 11, no. 8: 500. https://doi.org/10.3390/insects11080500