A Multiplex PCR Based on Mitochondrial COI Sequences for Identification of Members of the Anopheles barbirostris Complex (Diptera: Culicidae) in Thailand and Other Countries in the Region

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquitoes
2.2. DNA Extraction, Amplification and Sequencing of the COI Gene
2.3. Sequencing Alignment and Phylogenetic Analysis
2.4. Primer Design and Allele-Specific PCR
2.5. Multiplex PCR and Validation
3. Results
3.1. Phylogenetic Analysis
3.2. Primer Design and Multiplex PCR
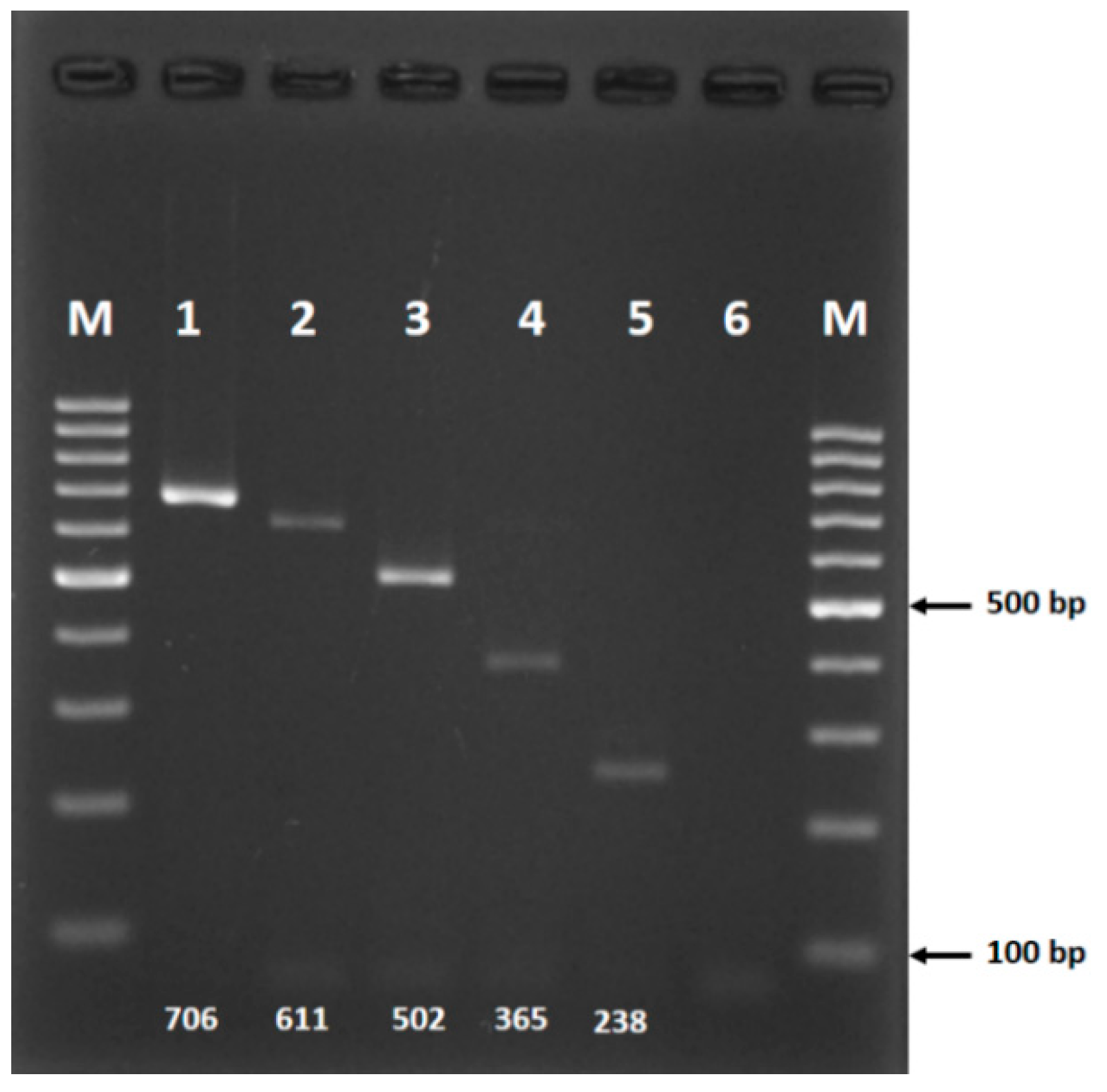
3.3. Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van der Wulp, F.M. Note XXXVIII. On exotic Diptera. Notes Leyden Mus. 1884, 6, 248–256. [Google Scholar]
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Chareonviriyaphap, T.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Elyazar, I.R.; Kabaria, C.W.; Harbach, R.E.; et al. The dominant Anopheles vectors of human malaria in the Asia-Pacific region: Occurrence data, distribution maps and bionomic precis. Parasit. Vectors 2011, 4, 89. [Google Scholar] [CrossRef] [Green Version]
- Satoto, T.B.T. Cryptic Species within Anopheles barbirostris van der Wulp, 1884, Inferred from Nuclear and Mitochondrial Gene Sequence Variation. Ph.D. Thesis, University of Liverpool, Liverpool, UK, 2001. [Google Scholar]
- Taai, K.; Harbach, R.E. Systematics of the Anopheles barbirostris species complex (Diptera: Culicidae: Anophelinae) in Thailand. Zool. J. Linn. Soc. 2015, 174, 244–264. [Google Scholar] [CrossRef] [Green Version]
- Saeung, A.; Baimai, V.; Otsuka, Y.; Rattanarithikul, R.; Somboon, P.; Junkum, A.; Tuetun, B.; Takaoka, H.; Choochote, W. Molecular and cytogenetic evidence of three sibling species of the Anopheles barbirostris Form A (Diptera: Culicidae) in Thailand. Parasitol. Res. 2008, 102, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Esquivel, C.; Donnelly, M.J.; Harbach, R.E.; Townson, H. A molecular phylogeny of mosquitoes in the Anopheles barbirostris Subgroup reveals cryptic species: Implications for identification of disease vectors. Mol. Phylogenet. Evol. 2009, 50, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Townson, H.; Dyer, N.; McAlister, E.; Satoto, T.B.T.; Bangs, M.J.; Harbach, R.E. Systematics of Anopheles barbirostris van der Wulp and a sibling species of the Barbirostris Complex (Diptera: Culicidae) in eastern Java, Indonesia. Syst. Entomol. 2013, 38, 180–191. [Google Scholar] [CrossRef]
- Elyazar, I.R.; Sinka, M.E.; Gething, P.W.; Tarmidzi, S.N.; Surya, A.; Kusriastuti, R.; Baird, J.K.; Hay, S.I.; Bangs, M.J. The distribution and bionomics of Anopheles malaria vector mosquitoes in Indonesia. Adv. Parasitol. 2013, 83, 173–266. [Google Scholar]
- Davidson, J.R.; Wahid, I.; Sudirman, R.; Makuru, V.; Hasan, H.; Arfah, A.M.; Nur, N.; Hidayat, M.Y.; Hendershot, A.L.; Xiao, H.; et al. Comparative field evaluation of kelambu traps, barrier screens and barrier screens with eaves for longitudinal surveillance of adult Anopheles mosquitoes in Sulawesi, Indonesia. Parasit. Vectors 2019, 12, 399. [Google Scholar] [CrossRef] [Green Version]
- Thongsahuan, S.; Baimai, V.; Junkum, A.; Saeung, A.; Min, G.-S.; Joshi, D.; Park, M.-H.; Somboon, P.; Suwonkerd, W.; Tippawangkosol, P. Susceptibility of Anopheles campestris-like and Anopheles barbirostris species complexes to Plasmodium falciparum and Plasmodium vivax in Thailand. Mem. Inst. Oswaldo Cruz 2011, 106, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Amerasinghe, P.H.; Amerasinghe, F.P.; Konradsen, F.; Fonseka, K.T.; Wirtz, R.A. Malaria vectors in a traditional dry zone village in Sri Lanka. Am. J. Trop. Med. Hyg. 1999, 60, 421–429. [Google Scholar] [CrossRef]
- Gajapathy, K.; Jude, P.J.; Goodacre, S.L.; Peiris, L.B.; Ramasamy, R.; Surendran, S.N. Molecular characterization of the malaria vector Anopheles barbirostris van der Wulp in Sri Lanka. Parasit. Vectors 2014, 7, 348. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, D.R.; Rajavel, A.R.; Mohapatra, P.K.; Jambulingam, P.; Mahanta, J.; Prakash, A. Faunal richness and the checklist of Indian mosquitoes (Diptera: Culicidae). Check List 2014, 10, 1342–1358. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, J.; Ma, Y. Molecular characterization of cryptic species of Anopheles barbirostris van der Wulp in China. Parasit. Vectors 2014, 7, 592. [Google Scholar] [CrossRef]
- Reid, J.A. The Anopheles barbirostris group (Diptera, Culicidae). Bull. Entomol. Res. 1962, 53, 1–57. [Google Scholar] [CrossRef]
- Reid, J.A. Anopheline mosquitoes of Malaya and Bornea. Stud. Inst. Med. Res. Malaya 1968, 31, 1–520. [Google Scholar]
- Harrison, B.A.; Klein, J.M. A revised list of the Anopheles of Cambodia. Mosq. Syst. 1975, 7, 9–12. [Google Scholar]
- Harrison, B.A.; Scanlon, J.E. Medical entomology studies–II. The subgenus Anopheles in Thailand (Diptera: Culicidae). Contri. Am. Entomol. Inst. (Ann. Arbor) 1975, 12, iv + 1–307. [Google Scholar]
- Ngo, C.T.; Dubois, G.; Sinou, V.; Parzy, D.; Le, H.Q.; Harbach, R.E.; Manguin, S. Diversity of Anopheles mosquitoes in Binh Phuoc and Dak Nong Provinces of Vietnam and their relation to disease. Parasit. Vectors 2014, 7, 316. [Google Scholar] [CrossRef] [Green Version]
- Baimai, V.; Rattanarithikul, R.; Kijchalao, U. Metaphase karyotypes of Anopheles of Thailand and Southeast Asia: IV. The Barbirostris and Umbrosus Species Groups, subgenus Anopheles (Diptera: Culicidae). J. Am. Mosq. Control. Assoc. 1995, 11, 323–328. [Google Scholar]
- Rattanarithikul, R.; Harrison, B.A.; Harbach, R.E.; Panthusiri, P.; Coleman, R.E. Illustrated keys to the mosquitoes of Thailand. IV. Anopheles. Southeast. Asian J. Trop. Med. Public Health 2006, 37 (Suppl. 2), 1–128. [Google Scholar]
- Limrat, D.; Rojruthai, B.; Apiwathnasorn, C.; Samung, Y.; Prommongkol, S. Anopheles barbirostris/campestris as a probable vector of malaria in Aranyaprathet, Sa Kaeo Province. Southeast. Asian J. Trop. Med. Public Health 2001, 32, 739–744. [Google Scholar] [PubMed]
- Apiwathnasorn, C.; Prommongkol, S.; Samung, Y.; Limrat, D.; Rojruthai, B. Potential for Anopheles campestris (Diptera: Culicidae) to transmit malaria parasites in Pa Rai subdistrict (Aranyaprathet, Sa Kaeo Province), Thailand. J. Med. Entomol. 2002, 39, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Saeung, A.; Otsuka, Y.; Baimai, V.; Somboon, P.; Pitasawat, B.; Tuetun, B.; Junkum, A.; Takaoka, H.; Choochote, W. Cytogenetic and molecular evidence for two species in the Anopheles barbirostris complex (Diptera: Culicidae) in Thailand. Parasitol. Res. 2007, 101, 1337–1344. [Google Scholar] [CrossRef] [PubMed]
- Brosseau, L.; Udom, C.; Sukkanon, C.; Chareonviriyaphap, T.; Bangs, M.J.; Saeung, A.; Manguin, S. A multiplex PCR assay for the identification of five species of the Anopheles barbirostris complex in Thailand. Parasit. Vectors 2019, 12, 223. [Google Scholar] [CrossRef] [PubMed]
- Suwannamit, S.; Baimai, V.; Otsuka, Y.; Saeung, A.; Thongsahuan, S.; Tuetun, B.; Apiwathnasorn, C.; Jariyapan, N.; Somboon, P.; Takaoka, H.; et al. Cytogenetic and molecular evidence for an additional new species within the taxon Anopheles barbirostris (Diptera: Culicidae) in Thailand. Parasitol. Res. 2009, 104, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Thongsahuan, S.; Baimai, V.; Otsuka, Y.; Saeung, A.; Tuetun, B.; Jariyapan, N.; Suwannamit, S.; Somboon, P.; Jitpakdi, A.; Takaoka, H. Karyotypic variation and geographic distribution of Anopheles campestris-like (Diptera: Culicidae) in Thailand. Mem. Inst. Oswaldo Cruz 2009, 104, 558–566. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Okimoto, R.; Dodgson, J.B. Improved PCR amplification of multiple specific alleles (PAMSA) using internally mismatched primers. Biotechniques 1996, 21, 20–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germer, S.; Higuchi, R. Single-tube genotyping without oligonucleotide probes. Genome Res. 1999, 9, 72–78. [Google Scholar]
- Gunathilaka, N.; Karunaraj, P. Identification of sibling species status of Anopheles culicifacies breeding in polluted water bodies in Trincomalee district of Sri Lanka. Malar. J. 2015, 14, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namgay, R.; Pemo, D.; Wangdi, T.; Phanitchakun, T.; Harbach, R.E.; Somboon, P. Molecular and morphological evidence for sibling species within Anopheles (Anopheles) lindesayi Giles (Diptera: Culicidae) in Bhutan. Acta Trop. 2020, 207, 105455. [Google Scholar] [CrossRef] [PubMed]
- Sum, J.S.; Lee, W.C.; Amir, A.; Braima, K.A.; Jeffery, J.; Abdul-Aziz, N.M.; Fong, M.Y.; Lau, Y.L. Phylogenetic study of six species of Anopheles mosquitoes in Peninsular Malaysia based on inter-transcribed spacer region 2 (ITS2) of ribosomal DNA. Parasit. Vectors 2014, 7, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]




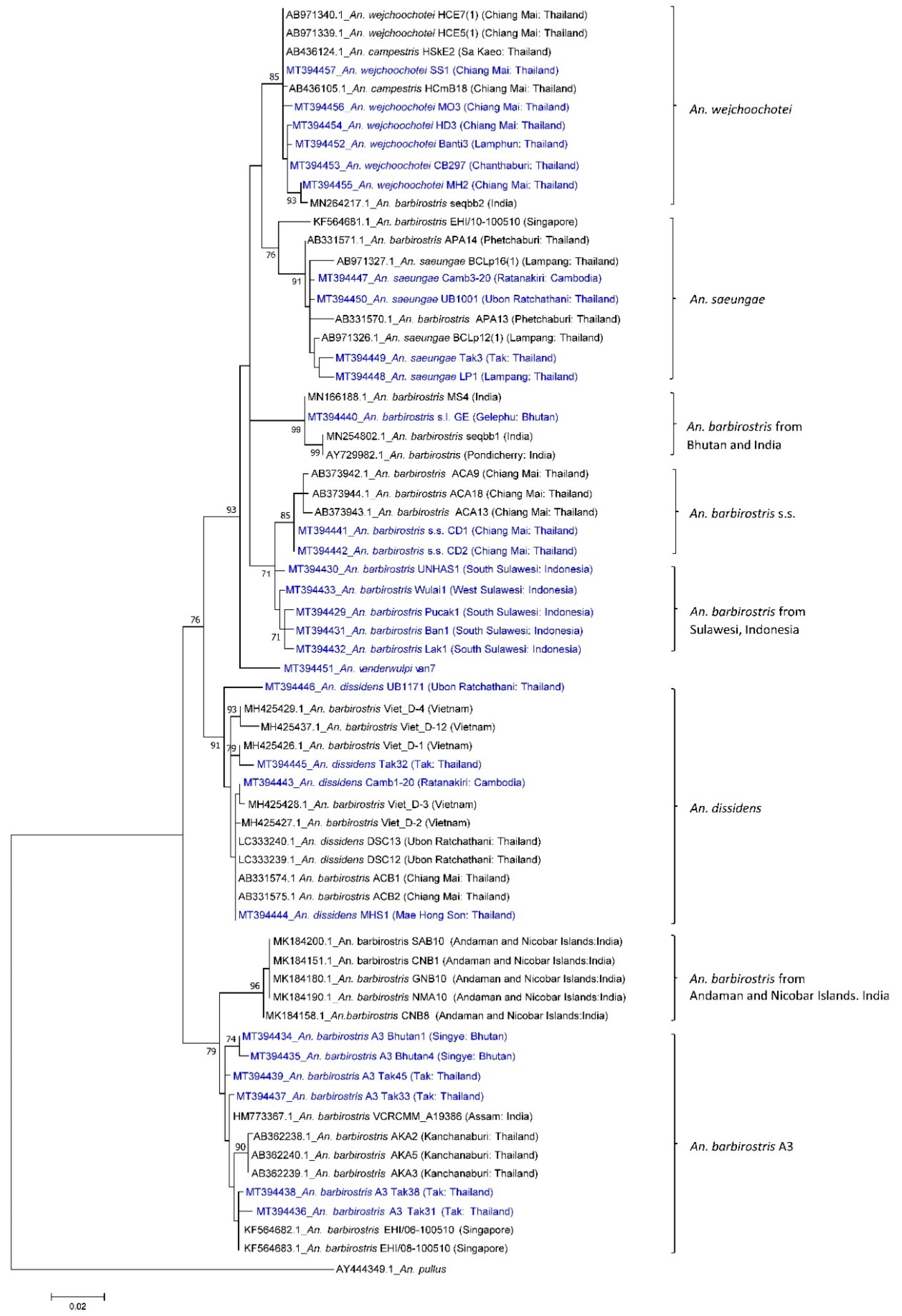{kind=link}
{kind=link}
{kind=link}
| Country | Location | Location Code | Coordinates | Multiplex PCR /COI Sequences (No. of Specimens) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| An. barbirostris s.s. | An. dissidens | An. saeungae | An. wejchoochotei | An. barbirostris A3 | Unknown | Total | ||||
| Thailand | Mae On, Chiang Mai | MO | 18°44′45″ N 99°13′27″ E | 7/7 | 7/7 | |||||
| Hang Dong, Chiang Mai | HD | 18°43′45″ N 98°54′22″ E | 3/3 | 3/3 | ||||||
| San Sai, Chiang Mai | SS | 18°59′09″ N 99°05′11″ E | 6/6 | 6/6 | ||||||
| Chiang Dao, Chiang Mai | CD | 19°23′27″ N 98°55′51″ E | 2/2 | 2/2 | ||||||
| Mueang Chiang Mai, Chiang Mai | MH | 18°45′21″ N 98°56′22″ E | 2/2 | 2/2 | ||||||
| Ko Kha, Lampang | LP | 18°07′44″ N 99°20′43″ E | 2/2 | 2/2 | ||||||
| Ban Thi, Lamphun | Banti | 18°39′16″ N 99°10′04″ E | 8/8 | 8/8 | ||||||
| Mae Sariang, Mae Hong Son | MS | 18°09′24″ N 97°56′15″ E | 3/3 | 3/3 | ||||||
| Tha Song Yang, Tak | Tak | 17°27′05″ N 98°10′40″ E | 3/3 | 1/1 | 9/9 | 13/13 | ||||
| Makham, Chanthaburi | CB | 12°40′17″ N 102°11′51″ E | 4/4 | 4/4 | ||||||
| Na Chaluai, Ubon Ratchathani | UB | 14°32′54″ N 105°14′33″ E | 38/38 | 3/3 | 41/41 | |||||
| Cambodia | Ratanakiri | Camb | 13°51′26″ N 107°06′04″ E | 5/5 | 1/1 | 6/6 | ||||
| Bhutan | Singye, Sarpang | Bhutan | 26°50′44″ N 90°12′26″ E | 2/2 | 2/2 | |||||
| Gelephu, Sarpang | GE | 26°54′47″ N 90°30′07″ E | 0/1 | 0/1 | ||||||
| Indonesia | Bantaeng, South Sulawesi | Ban | 05°26′58″ S 119°54′15″ E | 0/1 | 0/1 | |||||
| Makassar, South Sulawesi | Lak | 05°07′20″ S 119°28′05″ E | 0/5 | 0/5 | ||||||
| UNHAS | 05°07′54″ S 119°29′02″ E | 0/2 | 0/2 | |||||||
| Maros, South Sulawesi | Pucak | 05°09′29″ S 119°42′01″ E | 0/2 | 0/2 | ||||||
| Wulai, West Sulawesi | Wulai | 01°02′42″ S 119°31′27″ E | 0/1 | 0/1 | ||||||
| Total | 2/2 | 49/49 | 7/7 | 30/30 | 11/11 | 0/12 | 99/111 | |||
| Species | Primer Name | Primer Sequence (5′–3′) | Product Size (bp) |
|---|---|---|---|
| An. barbirostris s.s. | BARA4_F | [long GC tail] a AATAGTAGGAACTTCTTTATGA | 706 |
| An. dissidens | BARA1_F | ATTACTACTGTTATTAATATAGGA | 238 |
| An. saeungae | BARA2_F | TTAGGTCACCCAGGAGCA | 611 |
| An. wejchoochotei | BARWEJ_F | GATTTGGAAACTGATTACTG | 502 |
| An. barbirostris A3 | BARA3_F | CGGAACTGGATGAACTGTA | 365 |
| Universal reverse primer | HCO 2198_R | TAAACTTCAGGGTGACCAAAAAATCA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilai, P.; Namgay, R.; Made Ali, R.S.; Saingamsook, J.; Saeung, A.; Junkum, A.; Walton, C.; Harbach, R.E.; Somboon, P. A Multiplex PCR Based on Mitochondrial COI Sequences for Identification of Members of the Anopheles barbirostris Complex (Diptera: Culicidae) in Thailand and Other Countries in the Region. Insects 2020, 11, 409. https://doi.org/10.3390/insects11070409
Wilai P, Namgay R, Made Ali RS, Saingamsook J, Saeung A, Junkum A, Walton C, Harbach RE, Somboon P. A Multiplex PCR Based on Mitochondrial COI Sequences for Identification of Members of the Anopheles barbirostris Complex (Diptera: Culicidae) in Thailand and Other Countries in the Region. Insects. 2020; 11(7):409. https://doi.org/10.3390/insects11070409
Chicago/Turabian StyleWilai, Parinya, Rinzin Namgay, Rusdiyah Sudirman Made Ali, Jassada Saingamsook, Atiporn Saeung, Anuluck Junkum, Catherine Walton, Ralph E Harbach, and Pradya Somboon. 2020. "A Multiplex PCR Based on Mitochondrial COI Sequences for Identification of Members of the Anopheles barbirostris Complex (Diptera: Culicidae) in Thailand and Other Countries in the Region" Insects 11, no. 7: 409. https://doi.org/10.3390/insects11070409