
MicroRNA-155 and Disease-Related Immunohistochemical Parameters in Cutaneous Melanoma

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Silico Data Analysis
2.1.1. Identification of Differentially Expressed Melanoma-Related miRNAs
2.1.2. Evidence-Based Melanoma miRNA Regulon
2.2. In-House Clinical Validation
2.2.1. Study Population
2.2.2. Clinical and Pathological Assessment
2.2.3. RNA Extraction and Real-Time Reverse Transcriptase-Quantitative Polymerase Chain Reaction (RT-qPCR)
2.2.4. Statistical Analysis
2.3. Literature Screening and Review for the Role of miR-155 in Melanoma
3. Results
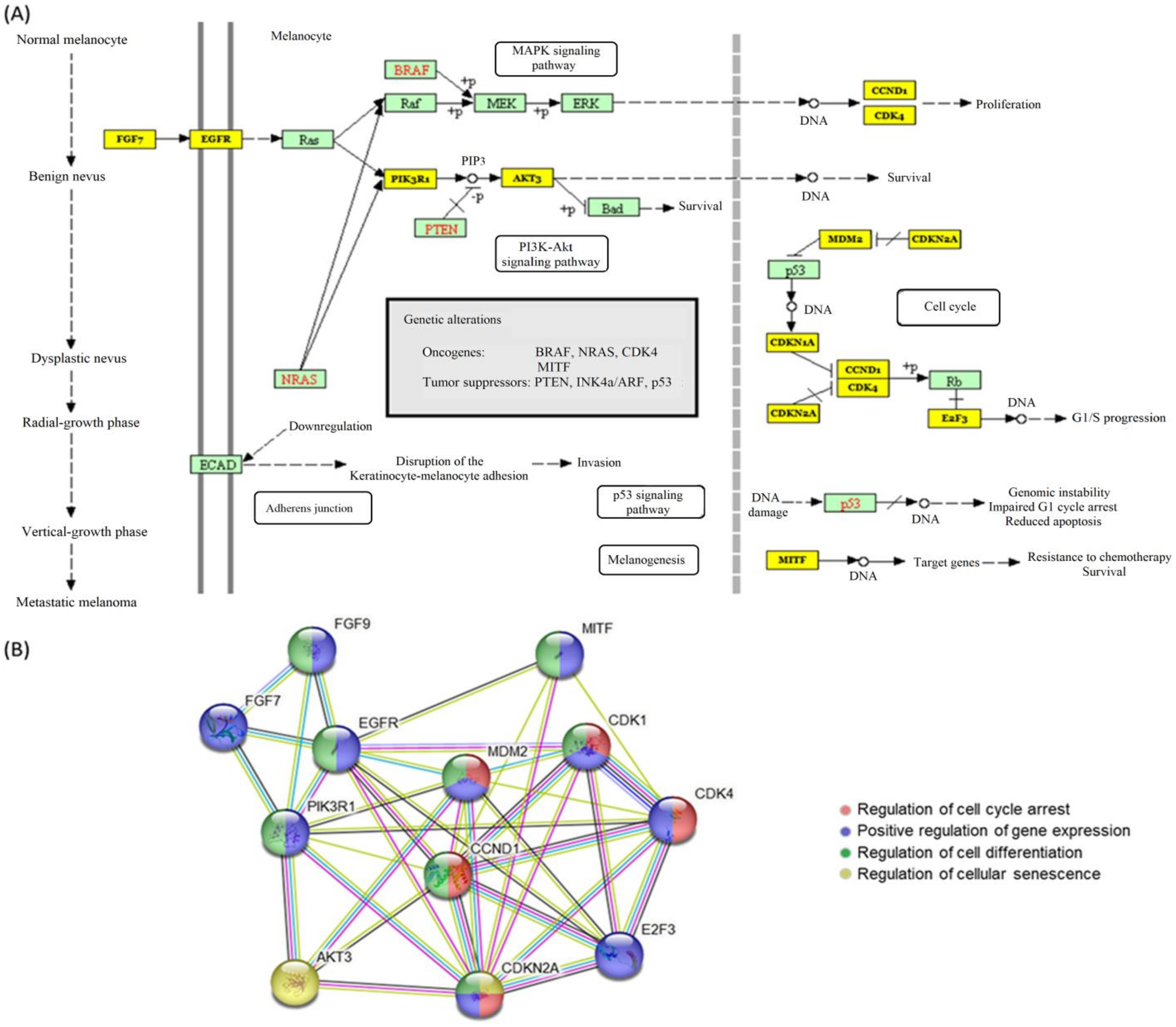
3.1. Database Screening for Deregulation of miR-155 in Melanoma
3.1.1. Deregulated miR-155 in Melanoma
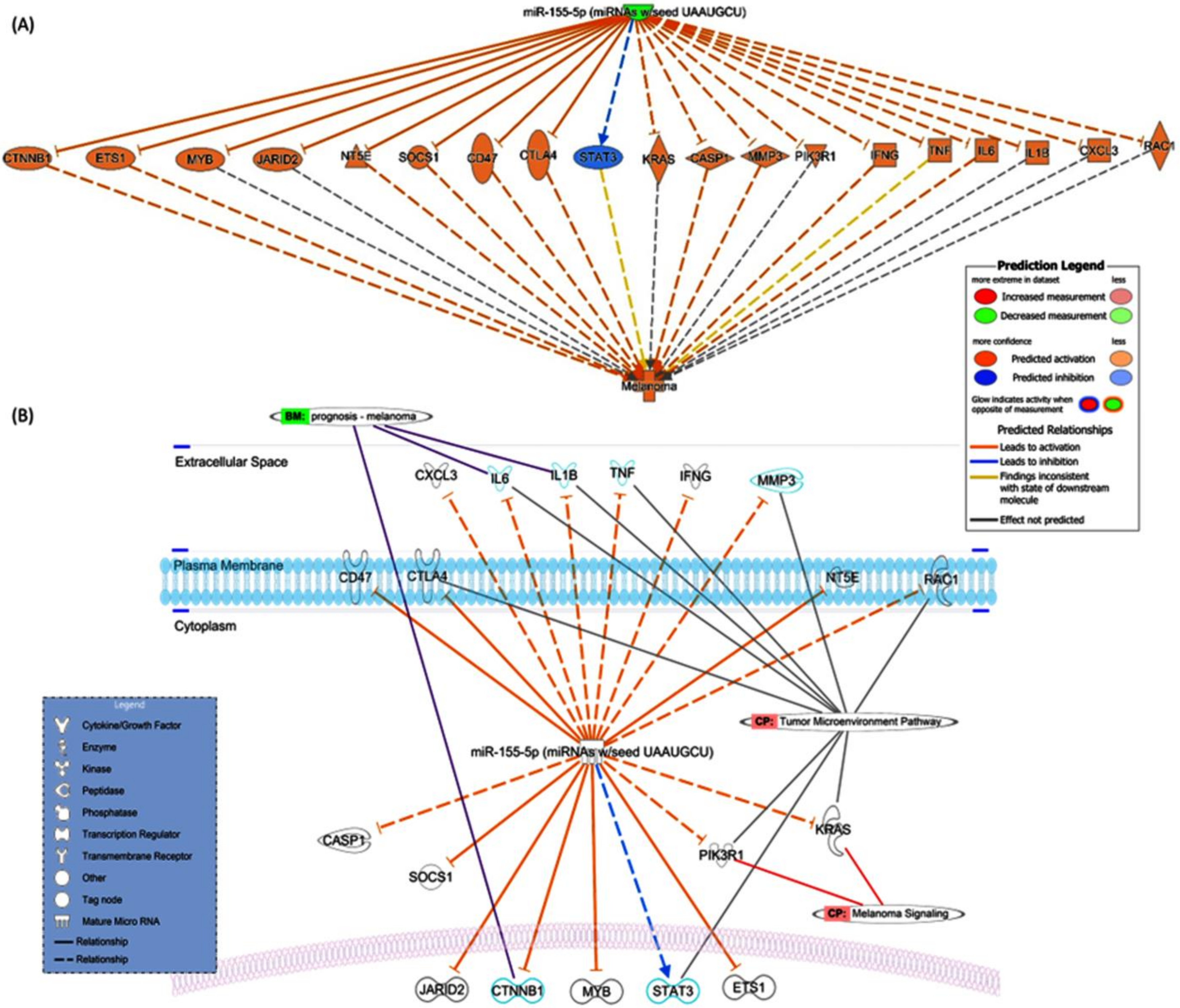
3.1.2. Molecular Targets of miR-155 in Melanoma
3.2. Validation in Melanoma Cohorts
3.2.1. Baseline Characteristics of In-House Patients
3.2.2. Comparison between Indolent and Recurrent Cohorts
3.2.3. Comparison between Survivors and Deceased Groups
3.2.4. Expression Pattern of miR-155 in Cancer Samples
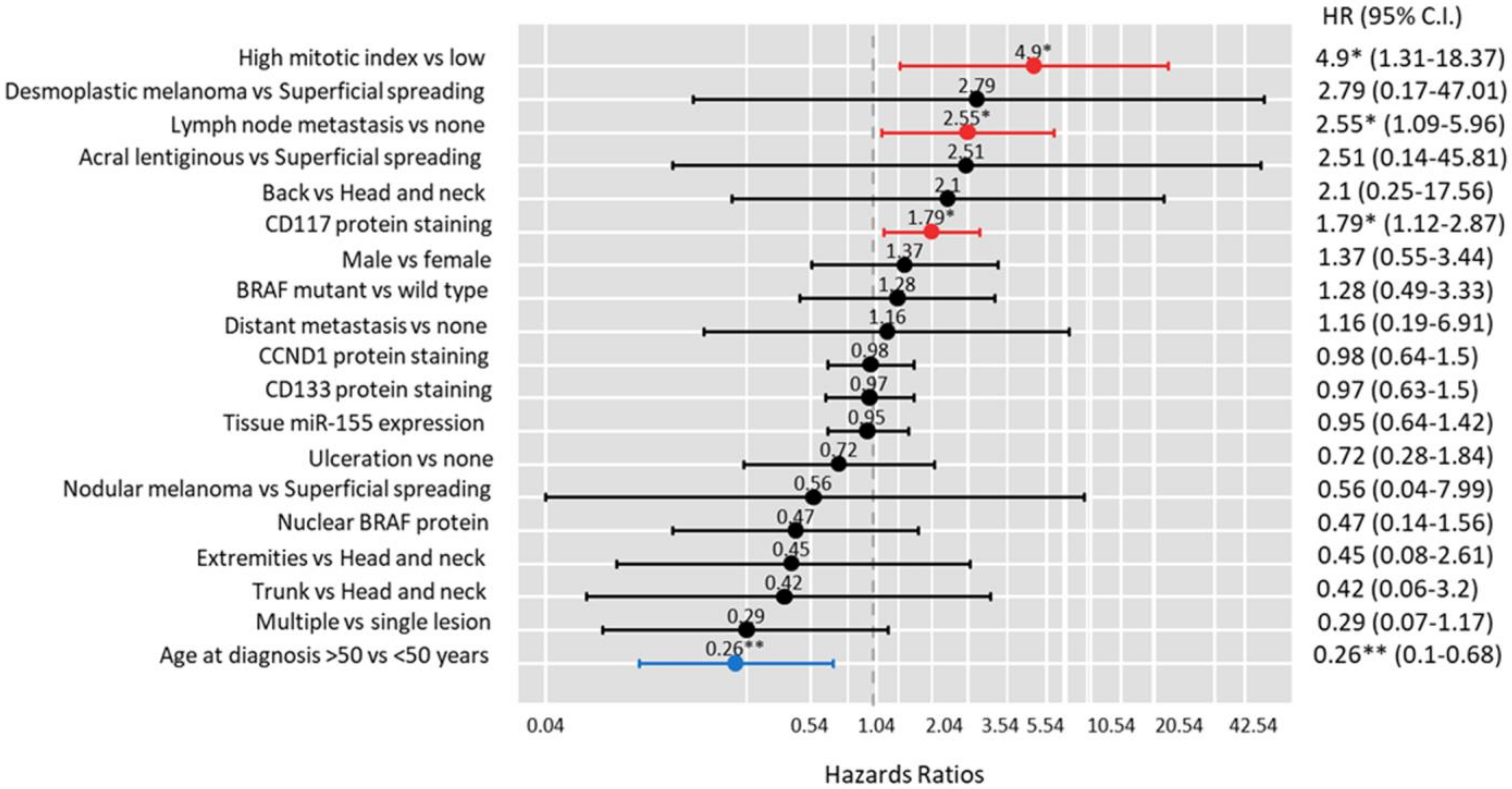
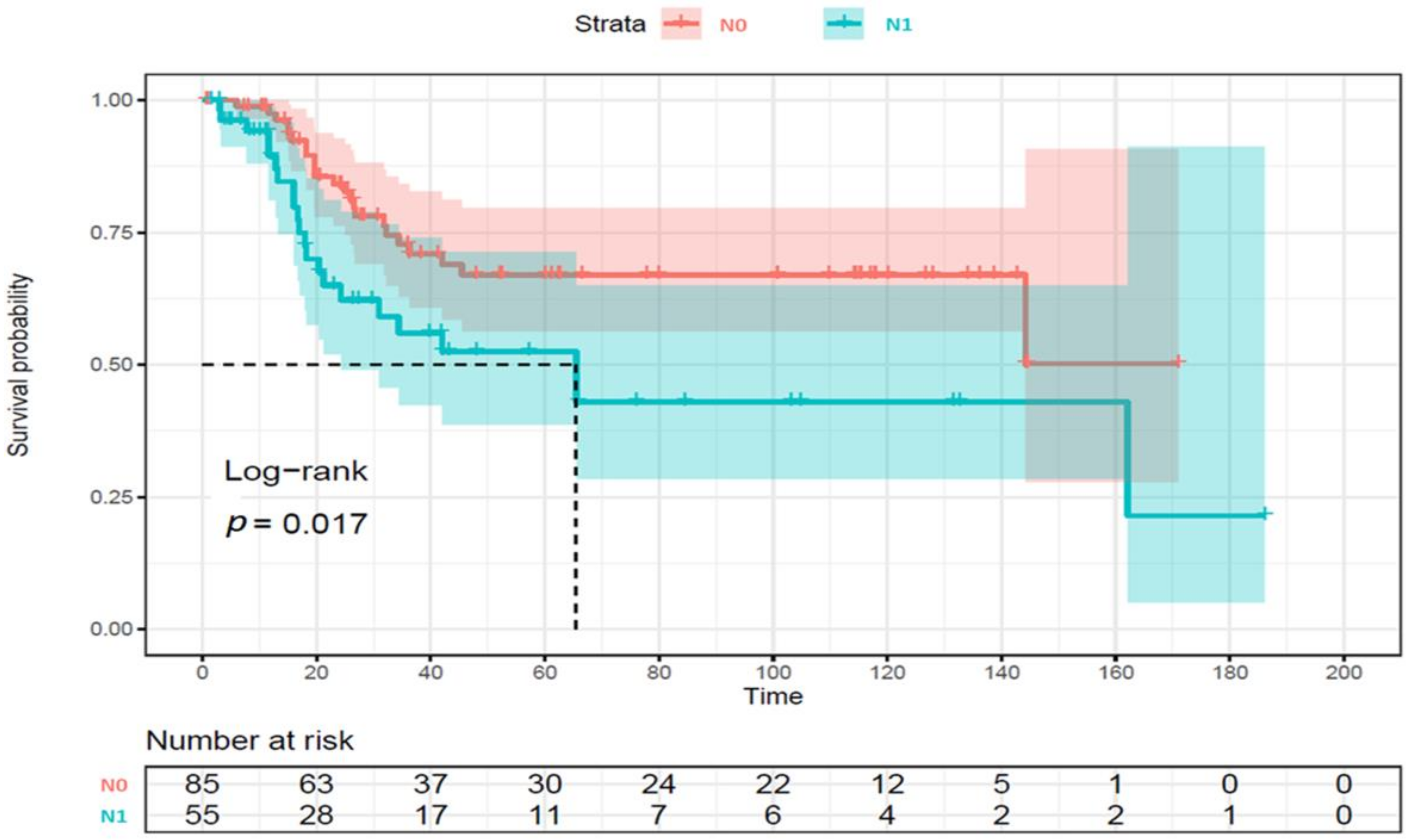
3.2.5. Survival Analysis
4. Discussion
4.1. Diagnostic Role of miR-155 in Human Subjects
4.2. Prognostic Role of miR-155 in Human Subjects
4.3. Expression Levels of miR-155 In Vitro and In Vivo
4.4. Potential Role of miR-155 in Melanoma Genesis
4.5. Role of miR-155 in the Tumor Microenvironment of Melanoma Samples
4.5.1. Role of miR-155 in Inflammation
4.5.2. miR-155 and T Cells
4.5.3. miR-155 and Cancer-Associated Fibroblasts (CAFs)
4.5.4. miR-155 and Macrophages
4.5.5. miR-155 and Myeloid-Derived Suppressor Cells (MDSCs)
4.5.6. miR-155 and Natural Killer Cells (NK Cells)
4.5.7. miR-155 and Tumor-Associated Mesenchymal Stem Cells (MSC)
4.6. Role of miR-155 in Predicting Treatment Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arnold, M.; Singh, D.; Laversanne, M.; Vignat, J.; Vaccarella, S.; Meheus, F.; Cust, A.E.; de Vries, E.; Whiteman, D.C.; Bray, F. Global Burden of Cutaneous Melanoma in 2020 and Projections to 2040. JAMA Dermatol. 2022, 158, 495–503. [Google Scholar] [CrossRef]
- Berman-Rosa, M.; Logan, J.; Ghazawi, F.M.; Le, M.; Conte, S.; Netchiporouk, E.; Mukovozov, I.M.; Cyr, J.; Mourad, A.; Miller, W.H.; et al. Analysis of Geographic and Environmental Factors and Their Association with Cutaneous Melanoma Incidence in Canada. Dermatology 2022, 238, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Conforti, C.; Zalaudek, I. Epidemiology and Risk Factors of Melanoma: A Review. Dermatol. Pract. Concept. 2021, 11, e2021161S. [Google Scholar] [CrossRef] [PubMed]
- Bellenghi, M.; Puglisi, R.; Pontecorvi, G.; De Feo, A.; Carè, A.; Mattia, G. Sex and Gender Disparities in Melanoma. Cancers 2020, 12, 1819. [Google Scholar] [CrossRef]
- Ward-Peterson, M.; Acuña, J.M.; Alkhalifah, M.K.; Nasiri, A.M.; Al-Akeel, E.S.; Alkhaldi, T.M.; Dawari, S.A.; Aldaham, S.A. Association Between Race/Ethnicity and Survival of Melanoma Patients in the United States Over 3 Decades: A Secondary Analysis of SEER Data. Medicine 2016, 95, e3315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, X.; Chen, D.; Yu, J. Radiotherapy combined with immunotherapy: The dawn of cancer treatment. Signal Transduct. Target. Ther. 2022, 7, 258. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.A.; Zhong, J.; Rosenbaum, B.E.; Utter, K.; Moran, U.; Darvishian, F.; Polsky, D.; Berman, R.S.; Shapiro, R.L.; Pavlick, A.C.; et al. Impact of initial stage on metastatic melanoma survival. Melanoma Res. 2019, 29, 281–288. [Google Scholar] [CrossRef]
- Revythis, A.; Shah, S.; Kutka, M.; Moschetta, M.; Ozturk, M.A.; Pappas-Gogos, G.; Ioannidou, E.; Sheriff, M.; Rassy, E.; Boussios, S. Unraveling the Wide Spectrum of Melanoma Biomarkers. Diagnostics 2021, 11, 1341. [Google Scholar] [CrossRef]
- Cantisani, C.; Ambrosio, L.; Cucchi, C.; Meznerics, F.A.; Kiss, N.; Bánvölgyi, A.; Rega, F.; Grignaffini, F.; Barbuto, F.; Frezza, F.; et al. Melanoma Detection by Non-Specialists: An Untapped Potential for Triage? Diagnostics 2022, 12, 2821. [Google Scholar] [CrossRef]
- Lan, H.; Lu, H.; Wang, X.; Jin, H. MicroRNAs as potential biomarkers in cancer: Opportunities and challenges. BioMed Res. Int. 2015, 2015, 125094. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating microRNAs as potential cancer biomarkers: The advantage and disadvantage. Clin. Epigenetics 2018, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.; Wang, H.; Yao, X.; Zhang, D.; Xie, Y.; Cui, R.; Zhang, X. Circulating MicroRNAs in Cancer: Potential and Challenge. Front. Genet. 2019, 10, 626. [Google Scholar] [CrossRef] [Green Version]
- Anglicheau, D.; Muthukumar, T.; Suthanthiran, M. MicroRNAs: Small RNAs with big effects. Transplantation 2010, 90, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Medina, P.P.; Slack, F.J. microRNAs and cancer: An overview. Cell Cycle 2008, 7, 2485–2492. [Google Scholar] [CrossRef] [Green Version]
- Eichmüller, S.B.; Osen, W.; Mandelboim, O.; Seliger, B. Immune Modulatory microRNAs Involved in Tumor Attack and Tumor Immune Escape. J. Natl. Cancer Ins. 2017, 109, djx034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otmani, K.; Lewalle, P. Tumor Suppressor miRNA in Cancer Cells and the Tumor Microenvironment: Mechanism of Deregulation and Clinical Implications. Front. Oncol. 2021, 11, 708765. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. microRNAs as oncogenes and tumor suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Qiu, C.; Zhang, H.; Wang, J.; Cui, Q.; Yin, Y. Human microRNA oncogenes and tumor suppressors show significantly different biological patterns: From functions to targets. PLoS ONE 2010, 5, e13067. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.I.; Katsura, A.; Matsuyama, H.; Miyazono, K. MicroRNA regulons in tumor microenvironment. Oncogene 2015, 34, 3085–3094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambari, R.; Brognara, E.; Spandidos, D.A.; Fabbri, E. Targeting oncomiRNAs and mimicking tumor suppressor miRNAs: Νew trends in the development of miRNA therapeutic strategies in oncology (Review). Int. J. Oncol. 2016, 49, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Gracias, D.T.; Stelekati, E.; Hope, J.L.; Boesteanu, A.C.; Doering, T.A.; Norton, J.; Mueller, Y.M.; Fraietta, J.A.; Wherry, E.J.; Turner, M.; et al. The microRNA miR-155 controls CD8(+) T cell responses by regulating interferon signaling. Nat. Immunol. 2013, 14, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.D.; Feng, S.Y.; Huang, A.F. Role of miR-155 in inflammatory autoimmune diseases: A comprehensive review. Inflamm. Res. 2022, 71, 1501–1517. [Google Scholar] [CrossRef]
- Cai, Z.K.; Chen, Q.; Chen, Y.B.; Gu, M.; Zheng, D.C.; Zhou, J.; Wang, Z. microRNA-155 promotes the proliferation of prostate cancer cells by targeting annexin 7. Mol. Med. Rep. 2015, 11, 533–538. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Zhang, H.W.; Lu, M.H.; He, X.H.; Li, Y.; Gu, H.; Liu, M.F.; Wang, E.D. MicroRNA-155 functions as an OncomiR in breast cancer by targeting the suppressor of cytokine signaling 1 gene. Cancer Res. 2010, 70, 3119–3127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar] [CrossRef] [Green Version]
- Kalkusova, K.; Taborska, P.; Stakheev, D.; Smrz, D. The Role of miR-155 in Antitumor Immunity. Cancers 2022, 14, 5414. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Opyrchal, M.; Lu, X. Harnessing tumorous flaws for immune supremacy: Is miRNA-155 the weak link in breast cancer progression? J. Clin. Investig. 2022, 132, 3010. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Xu, W.; Gong, F.; Chi, B.; Chen, J.; Zhou, L. MicroRNA-155 regulates the proliferation, cell cycle, apoptosis and migration of colon cancer cells and targets CBL. Exp. Ther. Med. 2017, 14, 4053–4060. [Google Scholar] [CrossRef]
- Kapral, M.; Wawszczyk, J.; Węglarz, L. Regulation of MicroRNA-155 and Its Related Genes Expression by Inositol Hexaphosphate in Colon Cancer Cells. Molecules 2019, 24, 4153. [Google Scholar] [CrossRef] [Green Version]
- Tili, E.; Croce, C.M.; Michaille, J.J. miR-155: On the crosstalk between inflammation and cancer. Int. Rev. Immunol. 2009, 28, 264–284. [Google Scholar] [CrossRef]
- Trotta, R.; Chen, L.; Ciarlariello, D.; Josyula, S.; Mao, C.; Costinean, S.; Yu, L.; Butchar, J.P.; Tridandapani, S.; Croce, C.M.; et al. miR-155 regulates IFN-γ production in natural killer cells. Blood 2012, 119, 3478–3485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashima, R. Physiological roles of miR-155. Immunology 2015, 145, 323–333. [Google Scholar] [CrossRef]
- Ekiz, H.A.; Huffaker, T.B.; Grossmann, A.H.; Stephens, W.Z.; Williams, M.A.; Round, J.L.; O’Connell, R.M. MicroRNA-155 coordinates the immunological landscape within murine melanoma and correlates with immunity in human cancers. JCI Insight 2019, 4, 6543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimaldi, A.M.; Nuzzo, S.; Condorelli, G.; Salvatore, M.; Incoronato, M. Prognostic and Clinicopathological Significance of MiR-155 in Breast Cancer: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 5834. [Google Scholar] [CrossRef]
- Xu, T.P.; Zhu, C.H.; Zhang, J.; Xia, R.; Wu, F.L.; Han, L.; Shen, H.; Liu, L.X.; Shu, Y.Q. MicroRNA-155 expression has prognostic value in patients with non-small cell lung cancer and digestive system carcinomas. Asian Pac. J. Cancer Prev. 2013, 14, 7085–7090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orosz, E.; Kiss, I.; Gyöngyi, Z.; Varjas, T. Expression of Circulating. In Vivo 2018, 32, 1333–1337. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, Y.; Guo, Q.; Diao, Y.; Liu, H.; Song, G.; Wang, W.; Zhang, Z.; Yin, H.; Li, L. Prognostic role of microRNA-155 in patients with leukemia: A meta-analysis. Clin. Chim. Acta 2018, 483, 6–13. [Google Scholar] [CrossRef]
- Wang, F.; Zhou, J.; Zhang, Y.; Wang, Y.; Cheng, L.; Bai, Y.; Ma, H. The Value of MicroRNA-155 as a Prognostic Factor for Survival in Non-Small Cell Lung Cancer: A Meta-Analysis. PLoS ONE 2015, 10, e0136889. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Xu, L.J.; Qin, J.J.; Zhang, L.; Zhuang, G.H. MicroRNA-155 inversely correlates with esophageal cancer progression through regulating tumor-associated macrophage FGF2 expression. Biochem. Biophys. Res. Commun. 2018, 503, 452–458. [Google Scholar] [CrossRef]
- Li, S.; Zhang, T.; Zhou, X.; Du, Z.; Chen, F.; Luo, J.; Liu, Q. The tumor suppressor role of miR-155-5p in gastric cancer. Oncol. Lett. 2018, 16, 2709–2714. [Google Scholar] [CrossRef] [Green Version]
- Levati, L.; Alvino, E.; Pagani, E.; Arcelli, D.; Caporaso, P.; Bondanza, S.; Di Leva, G.; Ferracin, M.; Volinia, S.; Bonmassar, E.; et al. Altered expression of selected microRNAs in melanoma: Antiproliferative and proapoptotic activity of miRNA-155. Int. J. Oncol. 2009, 35, 393–400. [Google Scholar]
- Philippidou, D.; Schmitt, M.; Moser, D.; Margue, C.; Nazarov, P.V.; Muller, A.; Vallar, L.; Nashan, D.; Behrmann, I.; Kreis, S. Signatures of microRNAs and selected microRNA target genes in human melanoma. Cancer Res. 2010, 70, 4163–4173. [Google Scholar] [CrossRef] [Green Version]
- Grignol, V.; Fairchild, E.T.; Zimmerer, J.M.; Lesinski, G.B.; Walker, M.J.; Magro, C.M.; Kacher, J.E.; Karpa, V.I.; Clark, J.; Nuovo, G.; et al. miR-21 and miR-155 are associated with mitotic activity and lesion depth of borderline melanocytic lesions. Br. J. Cancer. 2011, 105, 1023–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hufbauer, M.; Lazić, D.; Reinartz, M.; Akgül, B.; Pfister, H.; Weissenborn, S.J. Skin tumor formation in human papillomavirus 8 transgenic mice is associated with a deregulation of oncogenic miRNAs and their tumor suppressive targets. J. Dermatol. Sci. 2011, 64, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Kunz, M. MicroRNAs in melanoma biology. Adv. Exp. Med. Biol. 2013, 774, 103–120. [Google Scholar] [CrossRef]
- Joshi, P.; Kooshki, M.; Aldrich, W.; Varghai, D.; Zborowski, M.; Singh, A.D.; Triozzi, P.L. Expression of natural killer cell regulatory microRNA by uveal melanoma cancer stem cells. Clin. Exp. Metastasis 2016, 33, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Latchana, N.; Ganju, A.; Howard, J.H.; Carson, W.E. MicroRNA dysregulation in melanoma. Surg. Oncol. 2016, 25, 184–189. [Google Scholar] [CrossRef]
- Latchana, N.; Del Campo, S.E.; Grignol, V.P.; Clark, J.R.; Albert, S.P.; Zhang, J.; Wei, L.; Aldrink, J.H.; Nicol, K.K.; Ranalli, M.A.; et al. Classification of Indeterminate Melanocytic Lesions by MicroRNA Profiling. Ann. Surg. Oncol. 2017, 24, 347–354. [Google Scholar] [CrossRef]
- Yang, Z.; Ren, F.; Liu, C.; He, S.; Sun, G.; Gao, Q.; Yao, L.; Zhang, Y.; Miao, R.; Cao, Y.; et al. dbDEMC: A database of differentially expressed miRNAs in human cancers. BMC Genom. 2010, 11, S5. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Wang, Y.; Ling, Y.; Zhou, C.; Wang, H.; Teschendorff, A.E.; Zhao, Y.; Zhao, H.; He, Y.; Zhang, G.; et al. dbDEMC 3.0: Functional Exploration of Differentially Expressed miRNAs in Cancers of Human and Model Organisms. Genom. Proteom. Bioinform. 2022, 20, 446–454. [Google Scholar] [CrossRef]
- Scolyer, R.A.; Long, G.V.; Thompson, J.F. Evolving concepts in melanoma classification and their relevance to multidisciplinary melanoma patient care. Mol. Oncol. 2011, 5, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Ward, W.H.; Lambreton, F.; Goel, N.; Jian, Q.Y.; Farma, J.M. Clinical Presentation and Staging of Melanoma; Codon Publications: Brisbane, Australia, 2017; p. 79. [Google Scholar]
- Delgado, A.; Guddati, A.K. Clinical endpoints in oncology–A primer. Am. J. Cancer Res. 2021, 11, 1121–1131. [Google Scholar]
- Howe, K. Extraction of miRNAs from Formalin-Fixed Paraffin-Embedded (FFPE) Tissues. Methods Mol. Biol. 2017, 1509, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Toraih, E.A.; Fawzy, M.S.; Ning, B.; Zerfaoui, M.; Errami, Y.; Ruiz, E.M.; Hussein, M.H.; Haidari, M.; Bratton, M.; Tortelote, G.G.; et al. A miRNA-Based Prognostic Model to Trace Thyroid Cancer Recurrence. Cancers 2022, 14, 4128. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Toraih, E.A.; Fawzy, M.S.; Hussein, M.H.; El-Labban, M.M.; Ruiz, E.M.L.; Attia, A.A.; Halat, S.; Moroz, K.; Errami, Y.; Zerfaoui, M.; et al. MicroRNA-Based Risk Score for Predicting Tumor Progression Following Radioactive Iodine Ablation in Well-Differentiated Thyroid Cancer Patients: A Propensity-Score Matched Analysis. Cancers 2021, 13, 4649. [Google Scholar] [CrossRef]
- Park, S.; Eom, K.; Kim, J.; Bang, H.; Wang, H.Y.; Ahn, S.; Kim, G.; Jang, H.; Kim, S.; Lee, D.; et al. MiR-9, miR-21, and miR-155 as potential biomarkers for HPV positive and negative cervical cancer. BMC Cancer 2017, 17, 658. [Google Scholar] [CrossRef] [Green Version]
- Toraih, E.A.; Ruiz, E.; Ning, B.; Tortelote, G.G.; Hilliard, S.; Moroz, K.; Hu, T.; Fawzy, M.S.; Kandil, E. Chromatin-Accessible miRNA Regulons Driving Thyroid Tumorigenesis and Progression. J. Am. Coll. Surg. 2023, 236, 732–750. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yao, G.; Wang, J.; Yang, M.; Zhang, H.; Li, W. ncRPheno: A comprehensive database platform for identification and validation of disease related noncoding RNAs. RNA Biol. 2020, 17, 943–955. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef]
- Peng, J.; Liu, H.; Liu, C. MiR-155 Promotes Uveal Melanoma Cell Proliferation and Invasion by Regulating NDFIP1 Expression. Technol. Cancer Res. Treat. 2017, 16, 1160–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thyagarajan, A.; Tsai, K.Y.; Sahu, R.P. MicroRNA heterogeneity in melanoma progression. Semin. Cancer Biol. 2019, 59, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Poniewierska-Baran, A.; Słuczanowska-Głąbowska, S.; Małkowska, P.; Sierawska, O.; Zadroga, Ł.; Pawlik, A.; Niedźwiedzka-Rystwej, P. Role of miRNA in Melanoma Development and Progression. Int. J. Mol. Sci. 2022, 24, 201. [Google Scholar] [CrossRef]
- Korfiati, A.; Grafanaki, K.; Kyriakopoulos, G.C.; Skeparnias, I.; Georgiou, S.; Sakellaropoulos, G.; Stathopoulos, C. Revisiting miRNA Association with Melanoma Recurrence and Metastasis from a Machine Learning Point of View. Int. J. Mol. Sci. 2022, 23, 1299. [Google Scholar] [CrossRef] [PubMed]
- Poniewierska-Baran, A.; Zadroga, Ł.; Danilyan, E.; Małkowska, P.; Niedźwiedzka-Rystwej, P.; Pawlik, A. MicroRNA as a Diagnostic Tool, Therapeutic Target and Potential Biomarker in Cutaneous Malignant Melanoma Detection—Narrative Review. Int. J. Mol. Sci. 2023, 24, 5386. [Google Scholar] [CrossRef]
- Huber, V.; Vallacchi, V.; Fleming, V.; Hu, X.; Cova, A.; Dugo, M.; Shahaj, E.; Sulsenti, R.; Vergani, E.; Filipazzi, P.; et al. Tumor-derived microRNAs induce myeloid suppressor cells and predict immunotherapy resistance in melanoma. J. Clin. Investig. 2018, 128, 5505–5516. [Google Scholar] [CrossRef] [Green Version]
- Shu, S.; Yang, Y.; Allen, C.L.; Maguire, O.; Minderman, H.; Sen, A.; Ciesielski, M.J.; Collins, K.A.; Bush, P.J.; Singh, P.; et al. Metabolic reprogramming of stromal fibroblasts by melanoma exosome microRNA favours a pre-metastatic microenvironment. Sci. Rep. 2018, 8, 12905. [Google Scholar] [CrossRef] [Green Version]
- Daveri, E.; Vergani, E.; Shahaj, E.; Bergamaschi, L.; La Magra, S.; Dosi, M.; Castelli, C.; Rodolfo, M.; Rivoltini, L.; Vallacchi, V.; et al. microRNAs Shape Myeloid Cell-Mediated Resistance to Cancer Immunotherapy. Front. Immunol. 2020, 11, 1214. [Google Scholar] [CrossRef]
- Lunavat, T.R.; Cheng, L.; Kim, D.K.; Bhadury, J.; Jang, S.C.; Lässer, C.; Sharples, R.A.; López, M.D.; Nilsson, J.; Gho, Y.S.; et al. Small RNA deep sequencing discriminates subsets of extracellular vesicles released by melanoma cells–Evidence of unique microRNA cargos. RNA Biol. 2015, 12, 810–823. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Usatorre, A.; Sempere, L.F.; Carmona, S.J.; Carretero-Iglesia, L.; Monnot, G.; Speiser, D.E.; Rufer, N.; Donda, A.; Zehn, D.; Jandus, C.; et al. MicroRNA-155 Expression Is Enhanced by T-cell Receptor Stimulation Strength and Correlates with Improved Tumor Control in Melanoma. Cancer Immunol. Res. 2019, 7, 1013–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arts, N.; Cané, S.; Hennequart, M.; Lamy, J.; Bommer, G.; Van den Eynde, B.; De Plaen, E. microRNA-155, induced by interleukin-1ß, represses the expression of microphthalmia-associated transcription factor (MITF-M) in melanoma cells. PLoS ONE 2015, 10, e0122517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug. Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Levati, L.; Pagani, E.; Romani, S.; Castiglia, D.; Piccinni, E.; Covaciu, C.; Caporaso, P.; Bondanza, S.; Antonetti, F.R.; Bonmassar, E.; et al. MicroRNA-155 targets the SKI gene in human melanoma cell lines. Pigment Cell Melanoma Res. 2011, 24, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Gholipour, M.; Taheri, M. MicroRNA Signature in Melanoma: Biomarkers and Therapeutic Targets. Front. Oncol. 2021, 11, 608987. [Google Scholar] [CrossRef]
- Gencia, I.; Baderca, F.; Avram, S.; Gogulescu, A.; Marcu, A.; Seclaman, E.; Marian, C.; Solovan, C. A preliminary study of microRNA expression in different types of primary melanoma. Bosn. J. Basic. Med. Sci. 2020, 20, 197–208. [Google Scholar] [CrossRef]
- Bouamar, H.; Jiang, D.; Wang, L.; Lin, A.P.; Ortega, M.; Aguiar, R.C. MicroRNA 155 control of p53 activity is context dependent and mediated by Aicda and Socs1. Mol. Cell. Biol. 2015, 35, 1329–1340. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.T.; Luo, Y.H.; Li, A.L.; Tsai, J.C.; Wu, K.L.; Chung, P.J.; Ma, N. miRNA as a Modulator of Immunotherapy and Immune Response in Melanoma. Biomolecules 2021, 11, 1648. [Google Scholar] [CrossRef]
- Romano, G.; Kwong, L.N. miRNAs, Melanoma and Microenvironment: An Intricate Network. Int. J. Mol. Sci. 2017, 18, 2354. [Google Scholar] [CrossRef] [Green Version]
- Thyagarajan, A.; Shaban, A.; Sahu, R.P. MicroRNA-Directed Cancer Therapies: Implications in Melanoma Intervention. J. Pharmacol. Exp. Ther. 2018, 364, 1–12. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.S. The Interplay among miRNAs, Major Cytokines, and Cancer-Related Inflammation. Mol. Ther. Nucleic Acids 2020, 20, 606–620. [Google Scholar] [CrossRef] [PubMed]
- Mann, M.; Mehta, A.; Zhao, J.L.; Lee, K.; Marinov, G.K.; Garcia-Flores, Y.; Lu, L.F.; Rudensky, A.Y.; Baltimore, D. An NF-κB-microRNA regulatory network tunes macrophage inflammatory responses. Nat. Commun. 2017, 8, 851. [Google Scholar] [CrossRef] [Green Version]
- Tittarelli, A.; Navarrete, M.; Lizana, M.; Hofmann-Vega, F.; Salazar-Onfray, F. Hypoxic Melanoma Cells Deliver microRNAs to Dendritic Cells and Cytotoxic T Lymphocytes through Connexin-43 Channels. Int. J. Mol. Sci. 2020, 21, 7567. [Google Scholar] [CrossRef]
- Huffaker, T.B.; Lee, S.H.; Tang, W.W.; Wallace, J.A.; Alexander, M.; Runtsch, M.C.; Larsen, D.K.; Thompson, J.; Ramstead, A.G.; Voth, W.P.; et al. Antitumor immunity is defective in T cell-specific microRNA-155-deficient mice and is rescued by immune checkpoint blockade. J. Biol. Chem. 2017, 292, 18530–18541. [Google Scholar] [CrossRef] [Green Version]
- Dudda, J.C.; Salaun, B.; Ji, Y.; Palmer, D.C.; Monnot, G.C.; Merck, E.; Boudousquie, C.; Utzschneider, D.T.; Escobar, T.M.; Perret, R.; et al. MicroRNA-155 is required for effector CD8+ T cell responses to virus infection and cancer. Immunity 2013, 38, 742–753. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Nunez, R.T.; Louafi, F.; Sanchez-Elsner, T. The interleukin 13 (IL-13) pathway in human macrophages is modulated by microRNA-155 via direct targeting of interleukin 13 receptor alpha1 (IL13Ralpha1). J. Biol. Chem. 2011, 286, 1786–1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Yan, T.; Huang, C.; Xu, Z.; Wang, L.; Jiang, E.; Wang, H.; Chen, Y.; Liu, K.; Shao, Z.; et al. Melanoma cell-secreted exosomal miR-155-5p induce proangiogenic switch of cancer-associated fibroblasts via SOCS1/JAK2/STAT3 signaling pathway. J. Exp. Clin. Cancer Res. 2018, 37, 242. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, S.A.; Dudley, M.E. Adoptive cell therapy for the treatment of patients with metastatic melanoma. Curr. Opin. Immunol. 2009, 21, 233–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazari-Jahantigh, M.; Wei, Y.; Noels, H.; Akhtar, S.; Zhou, Z.; Koenen, R.R.; Heyll, K.; Gremse, F.; Kiessling, F.; Grommes, J.; et al. MicroRNA-155 promotes atherosclerosis by repressing Bcl6 in macrophages. J. Clin. Investig. 2012, 122, 4190–4202. [Google Scholar] [CrossRef] [Green Version]
- Fridman, W.H.; Pagès, F.; Sautès-Fridman, C.; Galon, J. The immune contexture in human tumours: Impact on clinical outcome. Nat. Rev. Cancer 2012, 12, 298–306. [Google Scholar] [CrossRef]
- Dong, L.; Tian, X.; Zhao, Y.; Tu, H.; Wong, A.; Yang, Y. The Roles of MiRNAs (MicroRNAs) in Melanoma Immunotherapy. Int. J. Mol. Sci. 2022, 23, 14775. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Vigorito, E.; Clare, S.; Warren, M.V.; Couttet, P.; Soond, D.R.; van Dongen, S.; Grocock, R.J.; Das, P.P.; Miska, E.A.; et al. Requirement of bic/microRNA-155 for normal immune function. Science 2007, 316, 608–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsin, J.P.; Lu, Y.; Loeb, G.B.; Leslie, C.S.; Rudensky, A.Y. The effect of cellular context on miR-155-mediated gene regulation in four major immune cell types. Nat. Immunol. 2018, 19, 1137–1145. [Google Scholar] [CrossRef]
- Ji, Y.; Wrzesinski, C.; Yu, Z.; Hu, J.; Gautam, S.; Hawk, N.V.; Telford, W.G.; Palmer, D.C.; Franco, Z.; Sukumar, M.; et al. miR-155 augments CD8+ T-cell antitumor activity in lymphoreplete hosts by enhancing responsiveness to homeostatic γc cytokines. Proc. Natl. Acad. Sci. USA 2015, 112, 476–481. [Google Scholar] [CrossRef] [Green Version]
- DiSano, J.A.; Huffnagle, I.; Gowda, R.; Spiegelman, V.S.; Robertson, G.P.; Pameijer, C.R. Loss of miR-155 upregulates WEE1 in metastatic melanoma. Melanoma Res. 2019, 29, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Song, J.B.; Chen, H.X.; Wang, Q.Q.; Meng, L.X.; Li, Y. MiR-155 inhibits proliferation, invasion and migration of melanoma via targeting CBL. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 9525–9534. [Google Scholar] [CrossRef]
- Chen, S.; Wang, L.; Fan, J.; Ye, C.; Dominguez, D.; Zhang, Y.; Curiel, T.J.; Fang, D.; Kuzel, T.M.; Zhang, B. Host miR155 promotes tumor growth through a myeloid-derived suppressor cell-dependent mechanism. Cancer Res. 2015, 75, 519–531. [Google Scholar] [CrossRef] [Green Version]
- Zonari, E.; Pucci, F.; Saini, M.; Mazzieri, R.; Politi, L.S.; Gentner, B.; Naldini, L. A role for miR-155 in enabling tumor-infiltrating innate immune cells to mount effective antitumor responses in mice. Blood 2013, 122, 243–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Zhang, J.; Diao, W.; Wang, D.; Wei, Y.; Zhang, C.Y.; Zen, K. MicroRNA-155 and MicroRNA-21 promote the expansion of functional myeloid-derived suppressor cells. J. Immunol. 2014, 192, 1034–1043. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Wang, M.; Yang, F.; Tian, Y.; Cai, J.; Yang, H.; Fu, H.; Mao, F.; Zhu, W.; Qian, H.; et al. miR-155-5p inhibition promotes the transition of bone marrow mesenchymal stem cells to gastric cancer tissue derived MSC-like cells via NF-κB p65 activation. Oncotarget 2016, 7, 16567–16580. [Google Scholar] [CrossRef] [Green Version]
- Monti, M.; Lunardini, S.; Magli, I.A.; Campi, R.; Primiceri, G.; Berardinelli, F.; Amparore, D.; Terracciano, D.; Lucarelli, G.; Schips, L.; et al. Micro-RNAs Predict Response to Systemic Treatments in Metastatic Renal Cell Carcinoma Patients: Results from a Systematic Review of the Literature. Biomedicines 2022, 10, 1287. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ma, T.; Huang, C.; Hu, T.; Li, J. The pivotal role of microRNA-155 in the control of cancer. J. Cell Physiol. 2014, 229, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, C.; De Summa, S.; Pinto, R.; Danza, K.; Tommasi, S. miRNAs as Key Players in the Management of Cutaneous Melanoma. Cells 2020, 9, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Level | Value | Characteristic | Level | Value |
|---|---|---|---|---|---|
| Demographics | Mitotic rate | <5 mitosis/hpf | 106 (75.7) | ||
| Age at diagnosis, y | Mean ± SD | 61.9 ± 14.9 | ≥5 mitosis/hpf | 34 (24.3) | |
| Sex | Male | 76 (54.3) | Breslow depth | Median (IQR) | 7.0 (3.0–12) |
| BMI, Kg/m2 | Mean ± SD | Clark level | I | 23 (16.4) | |
| Clinical data | II | 28 (20) | |||
| Origin | De novo | 140 (100) | III | 13 (9.3) | |
| Anatomic location | Head and neck | 12 (8.6) | IV | 39 (27.9) | |
| Extremities | 64 (45.7) | V | 37 (26.4) | ||
| Back | 9 (6.4) | Molecular and IHC-related data | |||
| Trunk | 55 (39.3) | CD133 staining * | Negative | 28 (20) | |
| Multiple lesions at presentation | Negative | 123 (87.9) | Weak intensity | 25 (17.9) | |
| Positive | 17 (12.1) | Moderate | 37 (26.4) | ||
| Subtype | Superficial spreading | 3 (2.1) | Strong intensity | 50 (35.7) | |
| Acral lentiginous | 25 (17.9) | CD117 staining * | Negative | 49 (35) | |
| Nodular | 106 (75.7) | Weak intensity | 46 (32.9) | ||
| Desmoplastic | 6 (4.3) | Moderate | 21 (15) | ||
| Ulceration | Negative | 47 (33.6) | Strong intensity | 24 (17.1) | |
| Positive | 93 (66.4) | CCND1 staining * | Negative | 24 (17.1) | |
| Clinical stage | I | 3 (2.1) | Weak intensity | 40 (28.6) | |
| IIA | 3 (2.1) | Moderate | 21 (15) | ||
| IIB | 21 (15.0) | Strong intensity | 55 (39.3) | ||
| IIC | 52 (37.1) | BRAFV600E * | Wild type | 62 (44.3) | |
| IIIA | 4 (2.9) | Mutant | 78 (55.7) | ||
| IIIB | 20 (14.3) | BRAF Protein * | Overexpression | 94 (67.1) | |
| IIIC | 20 (14.3) | miR-155-5p | Median (IQR) | −0.2 (−1.4, −0.7) | |
| IIID | 2 (1.4) | Treatment | |||
| IV | 15 (10.7) | Surgical margin | Free | 100 (71.4) | |
| Pathological data | Residue | 40 (28.6) | |||
| T stage | T1 | 1 (0.7) | Adjuvant therapy | Negative | 112 (80) |
| T2a | 3 (2.1) | Positive | 28 (20) | ||
| T2b | 4 (2.9) | Type of adjuvant therapy | CVD | 7 (5.0) | |
| T3a | 7 (5) | Interferon | 14 (10.0) | ||
| T3b | 8 (5.7) | DTC | 5 (3.6) | ||
| T4a | 31 (22.1) | Dacarbazine | 1 (0.7) | ||
| T4b | 86 (61.4) | Sunitinib | 1 (0.7) | ||
| N stage | N0 | 85 (60.7) | Radiotherapy | 1 (0.7) | |
| N1a | 9 (6.4) | Follow-up | |||
| N1b | 9 (6.4) | Relapse | Negative | 108 (77.1) | |
| N2a | 5 (3.6) | Positive | 32 (22.9) | ||
| N2b | 15 (10.7) | Recurrence or progression | Negative | 77 (55.0) | |
| N3 | 17 (12.1) | Positive | 63 (45.0) | ||
| M stage | M0 | 125 (89.3) | Died | Negative | 95 (67.9) |
| M1 | 15 (10.7) | Positive | 45 (32.1) |
| Variables | Levels | Remitted (n = 77) | Recurrence (n = 63) | p-Value |
|---|---|---|---|---|
| Demographics | ||||
| Age at diagnosis, years | Mean ± SD | 62.1 ± 15.0 | 61.6 ± 15.0 | 0.88 |
| Sex | Male | 40 (51.9) | 36 (57.1) | 0.61 |
| Body mass index, Kg/m2 | Mean ± SD | 26.5 ± 4.76 | 26.8 ± 6.14 | 0.82 |
| Diabetes mellitus | Positive | 13 (16.9) | 7 (11.1) | 0.47 |
| Hypertension | Positive | 14 (18.2) | 10 (15.9) | 0.83 |
| Hepatitis C | Positive | 10 (13) | 6 (9.5) | 0.60 |
| Clinical data | ||||
| Anatomic location | Head and neck | 5 (6.5) | 7 (11.1) | 0.61 |
| Extremities | 35 (45.5) | 29 (46) | ||
| Back | 4 (5.2) | 5 (7.9) | ||
| Trunk | 33 (42.9) | 22 (34.9) | ||
| Multiple lesions | Positive | 10 (13) | 7 (11.1) | 0.80 |
| Subtype | Superficial spreading | 2 (2.6) | 1 (1.6) | 0.39 |
| Acral lentiginous | 10 (13) | 15 (23.8) | ||
| Nodular melanoma | 62 (80.5) | 44 (69.8) | ||
| Desmoplastic | 3 (3.9) | 3 (4.8) | ||
| Ulceration | Positive | 49 (63.6) | 44 (69.8) | 0.48 |
| Clinical stage | I | 3 (3.9) | 0 (0) | 0.009 |
| II | 46 (59.7) | 30 (47.6) | ||
| III | 17 (22.1) | 29 (46) | ||
| IV | 11 (14.3) | 4 (6.3) | ||
| Pathological data | ||||
| Tumor size | T1 | 1 (1.3) | 0 (0) | 0.40 |
| T2 | 5 (6.5) | 2 (3.2) | ||
| T3 | 6 (7.8) | 9 (14.3) | ||
| T4 | 65 (84.4) | 52 (82.5) | ||
| Lymph node metastasis | Positive | 25 (32.5) | 30 (47.6) | 0.08 |
| Distant metastasis | Positive | 11 (14.3) | 4 (6.3) | 0.17 |
| Mitotic rate | ≥5 mitosis/hpf | 17 (22.1) | 17 (27) | 0.56 |
| Breslow depth | Median (IQR) | 7.0 (2.5–11.5) | 8.0 (4.0–13) | 0.026 |
| Clark level | I | 11 (14.3) | 12 (19) | 0.59 |
| II | 15 (19.5) | 13 (20.6) | ||
| III | 9 (11.7) | 4 (6.3) | ||
| IV | 24 (31.2) | 15 (23.8) | ||
| V | 18 (23.4) | 19 (30.2) | ||
| Treatment | ||||
| Surgical margin | Free | 68 (88.3) | 32 (50.8) | <0.001 |
| Residue | 9 (11.7) | 31 (49.2) | ||
| Adjuvant therapy | Positive | 11 (14.3) | 17 (27) | 0.09 |
| Follow-up | ||||
| Died | Positive | 0 (0.0) | 45 (71.4) | <0.001 |
| Variables | Levels | Survived (n = 95) | Died (n = 45) | p-Value |
|---|---|---|---|---|
| Demographics | ||||
| Age at diagnosis, years | Mean ± SD | 61.7 ± 14.8 | 62.2 ± 15.4 | 0.85 |
| Sex | Male | 48 (50.5) | 28 (62.2) | 0.21 |
| BMI, Kg/m2 | Mean ± SD | 27.1 ± 5.17 | 25.5 ± 5.7 | 0.17 |
| Diabetes mellitus | Positive | 13 (13.7) | 7 (15.6) | 0.79 |
| Hypertension | Positive | 17 (17.9) | 7 (15.6) | 0.81 |
| Hepatitis C | Positive | 10 (10.5) | 6 (13.3) | 0.77 |
| Clinical data | ||||
| Anatomic location | Head and neck | 6 (6.3) | 6 (13.3) | 0.23 |
| Extremities | 42 (44.2) | 22 (48.9) | ||
| Back | 5 (5.3) | 4 (8.9) | ||
| Trunk | 42 (44.2) | 13 (28.9) | ||
| Multiple lesions | Positive | 10 (10.5) | 7 (15.6) | 0.41 |
| Subtype | Superficial spreading | 2 (2.1) | 1 (2.2) | 0.07 |
| Acral lentiginous | 12 (12.6) | 13 (28.9) | ||
| Nodular melanoma | 78 (82.1) | 28 (62.2) | ||
| Desmoplastic | 3 (3.2) | 3 (6.7) | ||
| Ulceration | Positive | 61 (64.2) | 32 (71.1) | 0.45 |
| Clinical stage | I | 3 (3.2) | 0 (0) | 0.14 |
| II | 54 (56.8) | 22 (48.9) | ||
| III | 26 (27.4) | 20 (44.4) | ||
| IV | 12 (12.6) | 3 (6.7) | ||
| Pathological data | ||||
| Tumor size | T1 | 1 (1.1) | 0 (0) | 0.39 |
| T2 | 6 (6.3) | 1 (2.2) | ||
| T3 | 8 (8.4) | 7 (15.6) | ||
| T4 | 80 (84.2) | 37 (82.2) | ||
| LNM | Positive | 33 (34.7) | 22 (48.9) | 0.14 |
| Distant metastasis | Positive | 12 (12.6) | 3 (6.7) | 0.38 |
| Mitotic rate | ≥5 mitosis/hpf | 21 (22.1) | 13 (28.9) | 0.40 |
| Breslow depth | Median (IQR) | 7.0 (3.0–11.0) | 8.0 (4.0–14.5) | 0.26 |
| Clark level | I | 12 (12.6) | 11 (24.4) | 0.31 |
| II | 18 (18.9) | 10 (22.2) | ||
| III | 11 (11.6) | 2 (4.4) | ||
| IV | 28 (29.5) | 11 (24.4) | ||
| V | 26 (27.4) | 11 (24.4) | ||
| Treatment | ||||
| Surgical margin | Free | 80 (84.2) | 20 (44.4) | <0.001 |
| Residue | 15 (15.8) | 25 (55.6) | ||
| Adjuvant therapy | Positive | 13 (13.7) | 15 (33.3) | 0.012 |
| Follow-up | ||||
| Relapse | Positive | 11 (11.6) | 21 (46.7) | <0.001 |
| Recurrence | Positive | 18 (18.9) | 45 (100) | <0.001 |
| Variables | Number | Values | p-Value | |
|---|---|---|---|---|
| Demographics | ||||
| Age at diagnosis, years | <50 years | 30 | 0.24 (−1.30–1.04) | 0.26 |
| ≥50 years | 110 | −0.26 (−1.46–0.68) | ||
| Sex | Female | 64 | −0.16 (−1.45–0.97) | 0.48 |
| Male | 76 | −0.21 (−1.34–0.73) | ||
| Diabetes mellitus | Negative | 120 | −0.09 (−1.38–0.84) | |
| Positive | 20 | −0.97 (−1.57–0.10) | ||
| Hypertension | Negative | 116 | −0.17 (−1.36–0.78) | |
| Positive | 24 | −0.97 (−1.50–0.64) | ||
| Hepatitis C | Negative | 124 | −0.17 (−1.37–0.77) | |
| Positive | 16 | −0.83 (−2.08–1.03) | ||
| Clinical data | ||||
| Anatomic location | Head and neck | 12 | −0.17 (−1.50–1.14) | 0.73 |
| Extremities | 64 | −0.23 (−1.33–0.78) | ||
| Back | 9 | −0.25 (−1.57–1.54) | ||
| Trunk | 55 | 0.09 (−1.45–0.68) | ||
| Multiple lesions | Negative | 123 | −0.25 (−1.48–0.77) | 0.19 |
| Positive | 17 | 0.65 (−0.79–1.06) | ||
| Subtype | Superficial spreading | 3 | 2.84 (1.07–0.00) | 0.005 |
| Acral lentiginous | 25 | 0.27 (−1.44–0.92) | ||
| Nodular melanoma | 106 | −0.41 (−1.45–0.66) | ||
| Desmoplastic | 6 | 1.04 (−0.41–1.82) | ||
| Ulceration | Negative | 47 | 0.19 (−1.42–0.78) | 0.20 |
| Positive | 93 | −0.29 (−1.42–0.77) | ||
| Clinical stage | I | 3 | −0.82 (−0.83–0.00) | 0.69 |
| II | 76 | 0.05 (−1.51–0.87) | ||
| III | 46 | −0.52 (−1.37–0.61) | ||
| IV | 15 | 0.42 (−1.24–0.78) | ||
| Pathological data | ||||
| Tumor size | T1 | 1 | −0.83 (−1.28–0.19) | 0.65 |
| T2 | 7 | −0.29 (−1.26–0.68) | ||
| T3 | 15 | −0.13 (−1.50–0.80) | ||
| T4 | 117 | −0.83 (−1.12–−0.03) | ||
| LNM | Negative | 85 | 0.10 (−1.51–0.91) | 0.41 |
| Positive | 55 | −0.29 (−1.29–0.67) | ||
| Distant metastasis | Negative | 125 | −0.25 (−1.47–0.77) | 0.46 |
| Positive | 15 | 0.42 (−1.24–0.78) | ||
| Mitotic rate | <5 mitosis/hpf | 106 | −0.57 (−1.52–0.63) | 0.010 |
| ≥5 mitosis/hpf | 34 | 0.66 (−0.41–1.15) | ||
| Clark level | I | 23 | 0.51 (−1.51–1.76) | 0.18 |
| II | 28 | 0.21 (−1.22–0.83) | ||
| III | 13 | −1.31 (−2.12–1.40) | ||
| IV | 39 | −0.12 (−1.48–0.48) | ||
| V | 37 | −0.29 (−1.42–0.64) | ||
| Treatment | ||||
| Surgical margin | Free | 100 | −0.23 (−1.38–0.77) | 0.95 |
| Residue | 40 | −0.15 (−1.51–0.81) | ||
| Molecular markers | ||||
| CD133 | Negative | 28 | −0.52 (−1.65–0.68) | 0.37 |
| Weak | 25 | −0.83 (−1.38–0.46) | ||
| Moderate | 37 | −0.12 (−1.39–0.94) | ||
| Intense | 50 | 0.18 (−1.31–0.80) | ||
| CD117 | Negative | 49 | −0.29 (−1.52–0.73) | 0.52 |
| Weak | 46 | 0.20 (−1.41–1.15) | ||
| Moderate | 21 | 0.16 (−1.38–0.57) | ||
| Intense | 24 | −0.59 (−1.48–0.33) | ||
| CCND1 | Negative | 24 | −0.52 (−1.65–0.68) | 0.87 |
| Weak | 40 | −0.83 (−1.38–0.46) | ||
| Moderate | 21 | −0.12 (−1.39–0.94) | ||
| Intense | 55 | 0.18 (−1.31–0.80) | ||
| BRAFV600E | Negative | 62 | −0.09 (−1.30–0.83) | 0.32 |
| Positive | 78 | −0.26 (−1.49–0.70) | ||
| Nuclear BRAF | Negative | 97 | −0.29 (−1.44–0.77) | 0.55 |
| Positive | 42 | −0.01 (−1.41–0.81) | ||
| Follow-up | ||||
| Relapse | Negative | 108 | −0.18 (−1.44–0.80) | 0.88 |
| Positive | 32 | −0.21 (−1.38–0.68) | ||
| Recurrence | Negative | 77 | −0.13 (−1.45–0.89) | 0.89 |
| Positive | 63 | −0.26 (−1.36–0.68) | ||
| Survival | Survived | 95 | −0.16 (−1.45–0.89) | 0.98 |
| Deceased | 45 | −0.25 (−1.32–0.63) | ||
| Risk Factors | Recurrence | Overall Survival | ||
|---|---|---|---|---|
| HR (95% CI) | p-Value | HR (95% CI) | p-Value | |
| Age at diagnosis >50 vs. <50 years | 0.90 (0.48–1.72) | 0.76 | 0.87 (0.38–1.99) | 0.75 |
| Male vs. female | 1.31 (0.72–2.38) | 0.38 | 1.35 (0.66–2.79) | 0.41 |
| Extremities vs. Head and neck | 0.48 (0.19–1.24) | 0.13 | 0.49 (0.16–1.45) | 0.20 |
| Back vs. Head and neck | 1.11 (0.28–4.46) | 0.88 | 1.38 (0.29–6.57) | 0.69 |
| Trunk vs. Head and neck | 0.46 (0.15–1.44) | 0.18 | 0.48 (0.13–1.75) | 0.26 |
| Acral lentiginous vs. Superficial spreading | 1.59 (0.14–18.47) | 0.71 | 1.73 (0.13–23.44) | 0.68 |
| Nodular melanoma vs. Superficial spreading | 1.27 (0.13–12.68) | 0.84 | 0.55 (0.05–6.09) | 0.63 |
| Desmoplastic vs. Superficial spreading | 0.82 (0.06–10.41) | 0.88 | 0.91 (0.07–12.71) | 0.95 |
| Multiple vs. single lesion | 0.81 (0.30–2.21) | 0.68 | 0.66 (0.22–2.00) | 0.46 |
| Ulceration vs. none | 1.23 (0.65–2.32) | 0.53 | 1.02 (0.46–2.28) | 0.95 |
| Lymph node metastasis vs. none | 2.29 (1.31–4.03) | 0.004 | 2.10 (1.08–4.09) | 0.029 |
| Distant metastasis vs. none | 1.48 (0.43–5.05) | 0.53 | 0.71 (0.17–3.00) | 0.64 |
| High mitotic index vs. low | 2.15 (0.96–4.80) | 0.06 | 1.69 (0.61–4.67) | 0.31 |
| Tissue miR-155 expression | 1.04 (0.83–1.31) | 0.74 | 1.03 (0.79–1.34) | 0.81 |
| CD133 protein staining | 0.89 (0.68–1.16) | 0.39 | 0.92 (0.67–1.26) | 0.60 |
| CD117 protein staining | 1.25 (0.94–1.65) | 0.12 | 1.31 (0.92–1.87) | 0.13 |
| CCND1 protein staining | 1.01 (0.79–1.30) | 0.93 | 1.02 (0.75–1.40) | 0.88 |
| BRAF mutant vs. wild type | 1.14 (0.61–2.13) | 0.68 | 1.19 (0.57–2.48) | 0.65 |
| Nuclear BRAF protein | 0.62 (0.29–1.31) | 0.21 | 0.87 (0.37–2.03) | 0.74 |
| Direction | Downstream Aberration | Function | Experimental Design | References |
|---|---|---|---|---|
| ↑ miR-155 | Targets: STAT5, SHIP1, SOCS1, and PTPN2 ↓ Akt and Stat5 signaling ↓ | Enhancing CD8+ T-cell antitumor responses | In melanoma cell lines and murine models | [33,72,74,84,85,86] |
| ↑ miR-155 | Glycolysis metabolism ↑ Oxidative phosphorylation ↓ | Formation of CAFs | Only in melanoma cell lines | [87] |
| ↑ miR-155 | Direct target: SOCS1 ↓ JAK2/STAT3 signaling ↑ Proangiogenic factors: VEGFa, FGF2, and MMP9 ↑ | Formation and the proangiogenic switch of CAFs | In melanoma cell lines and murine models | [88] |
| ↑ miR-155 | IFN-inducible genes ↑ | Promoting the activation of M2 macrophages | In melanoma cell lines and murine models | [87,89,90] |
| ↑ miR-155 | STAT3 downstream signaling pathways ↓ | Polarization and T-cell-inhibiting functions of MDSCs | In melanoma cell lines and murine models | [68] |
| ↑ miR-155 | IFN-c and granzyme B production and NKG2D expression ↑ | Decreasing the sensitivity of melanoma cells to NK cells’ cytolysis | Only in melanoma cell lines | [31] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fawzy, M.S.; Ibrahiem, A.T.; Bayomy, N.A.; Makhdoom, A.K.; Alanazi, K.S.; Alanazi, A.M.; Mukhlef, A.M.; Toraih, E.A. MicroRNA-155 and Disease-Related Immunohistochemical Parameters in Cutaneous Melanoma. Diagnostics 2023, 13, 1205. https://doi.org/10.3390/diagnostics13061205
Fawzy MS, Ibrahiem AT, Bayomy NA, Makhdoom AK, Alanazi KS, Alanazi AM, Mukhlef AM, Toraih EA. MicroRNA-155 and Disease-Related Immunohistochemical Parameters in Cutaneous Melanoma. Diagnostics. 2023; 13(6):1205. https://doi.org/10.3390/diagnostics13061205
Chicago/Turabian StyleFawzy, Manal S., Afaf T. Ibrahiem, Naglaa A. Bayomy, Amin K. Makhdoom, Khalid S. Alanazi, Abdulaziz M. Alanazi, Abdulaziz M. Mukhlef, and Eman A. Toraih. 2023. "MicroRNA-155 and Disease-Related Immunohistochemical Parameters in Cutaneous Melanoma" Diagnostics 13, no. 6: 1205. https://doi.org/10.3390/diagnostics13061205