
Involvement of APOBEC3A/B Deletion in Mouse Mammary Tumor Virus (MMTV)-like Positive Human Breast Cancer

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects and Data
2.2. Detection of MMTV-Like env Gene and APOBEC3A/B Genotyping
2.3. Statistical Analyses
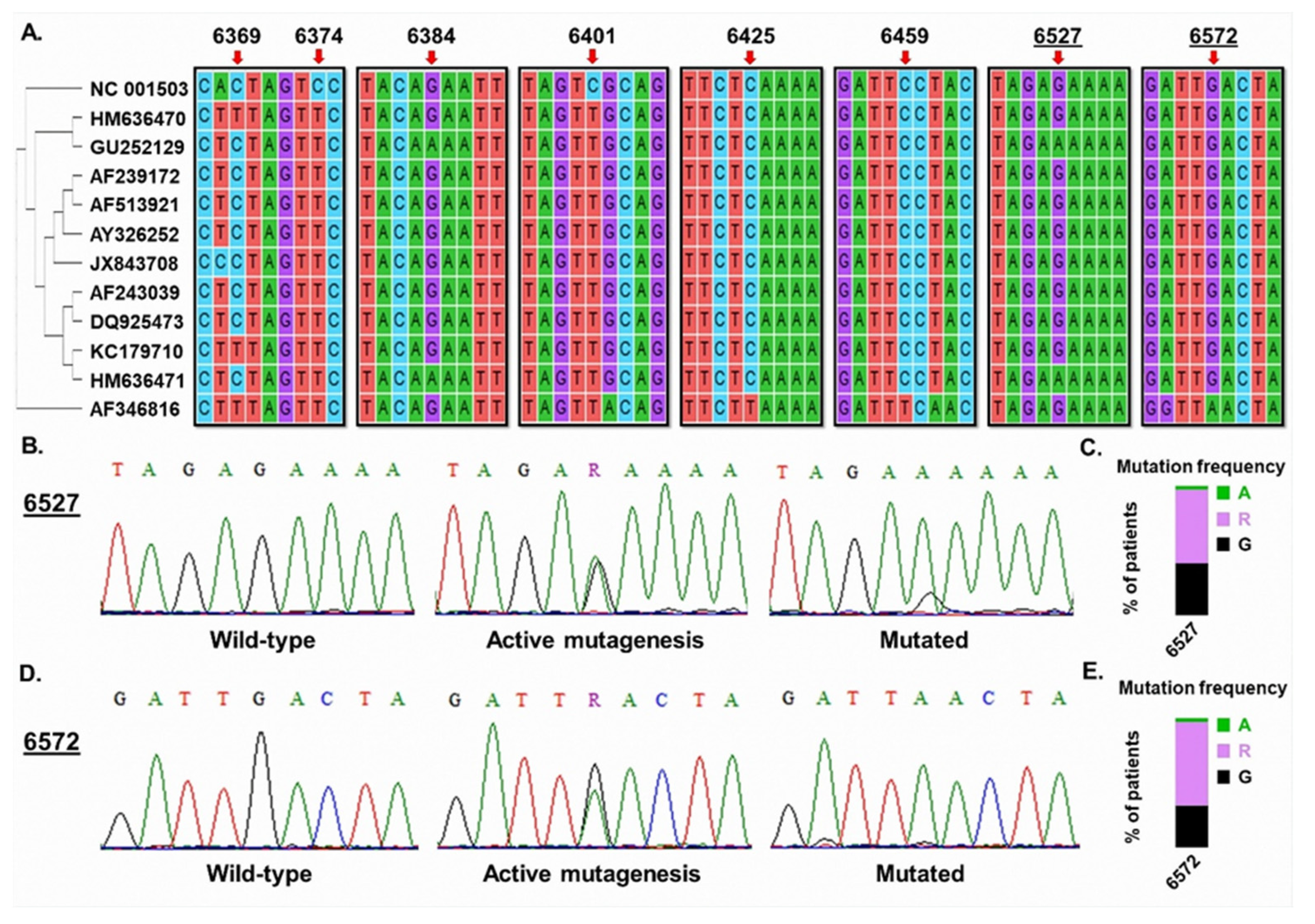
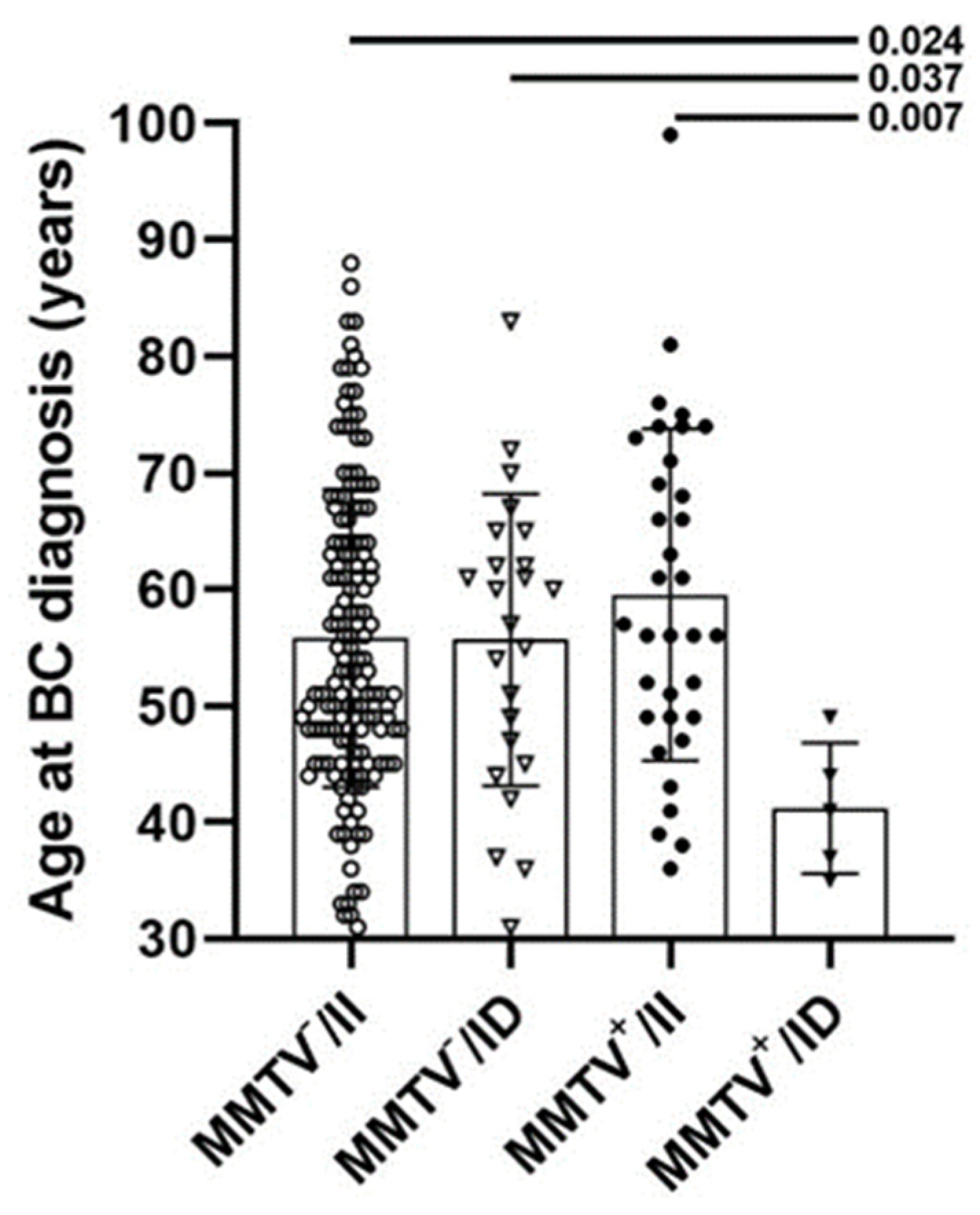
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- INCA. Estimativa 2023: Incidência de Câncer No Brasil; Instituto Nacional do Câncer: Rio de Janeiro, RJ, Brazil, 2022. [Google Scholar]
- Cirqueira, M.B.; Moreira, M.A.R.; Soares, L.R.; Freitas-junior, R. Subtipos Moleculares do Câncer de Mama. FEMINA 2011, 30, 499–503. [Google Scholar]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sørlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, J.; Åsvold, B.O.; Opdahl, S.; Tretli, S.; Vatten, L.J. Reproductive factors and the risk of breast cancer in old age: A Norwegian cohort study. Breast Cancer Res. Treat. 2013, 139, 237–243. [Google Scholar] [CrossRef]
- Brewer, H.R.; Jones, M.E.; Schoemaker, M.J.; Ashworth, A.; Swerdlow, A.J. Family history and risk of breast cancer: An analysis accounting for family structure. Breast Cancer Res. Treat. 2017, 165, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Makarem, N.; Chandran, U.; Bandera, E.V.; Parekh, N. Dietary fat in breast cancer survival. Annu. Rev. Nutr. 2013, 33, 319–348. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.; Wang, M.; Anderson, K.; Baglietto, L.; Bergkvist, L.; Bernstein, L.; van den Brandt, P.A.; Brinton, L.; Buring, J.E.; Eliassen, A.H.; et al. Alcohol consumption and breast cancer risk by estrogen receptor status: In a pooled analysis of 20 studies. Int. J. Epidemiol. 2016, 45, 916–928. [Google Scholar] [CrossRef] [Green Version]
- Moore, P.S.; Chang, Y. Why do viruses cause cancer? Highlights of the first century of human tumour virology. Nat. Rev. Cancer 2010, 10, 878–889. [Google Scholar] [CrossRef] [Green Version]
- Gaglia, M.M.; Munger, K. More than just oncogenes: Mechanisms of tumorigenesis by human viruses. Curr. Opin. Virol. 2018, 32, 48–59. [Google Scholar] [CrossRef]
- Bittner, J.J. Some possible effects of nursing on the mammary gland tumor incidence in mice. Science 1936, 84, 162. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wang, Y.; Melana, S.M.; Pelisson, I.; Najfeld, V.; Holland, J.F.; Pogo, B.G. Identification of a proviral structure in human breast cancer. Cancer Res. 2001, 61, 1754–1759. [Google Scholar] [PubMed]
- Amarante, M.K.; de Sousa Pereira, N.; Vitiello, G.A.F.; Watanabe, M.A.E. Involvement of a mouse mammary tumor virus (MMTV) homologue in human breast cancer: Evidence for, against and possible causes of controversies. Microb. Pathog. 2019, 130, 283–294. [Google Scholar] [CrossRef]
- Lawson, J.S.; Glenn, W.K. Evidence for a causal role by mouse mammary tumour-like virus in human breast cancer. NPJ Breast Cancer 2019, 5, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitiello, G.A.F.; Ferreira, W.A.S.; Cordeiro de Lima, V.C.; Medina, T.d.S. Antiviral Responses in Cancer: Boosting Antitumor Immunity through Activation of Interferon Pathway in the Tumor Microenvironment. Front. Immunol. 2021, 12, 782852. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W. Interferon-Stimulated Genes: What Do They All Do? Annu. Rev. Virol. 2019, 6, 567–584. [Google Scholar] [CrossRef]
- Alexandrov, L.B.; Kim, J.; Haradhvala, N.J.; Huang, M.N.; Tian Ng, A.W.; Wu, Y.; Boot, A.; Covington, K.R.; Gordenin, D.A.; Bergstrom, E.N.; et al. The repertoire of mutational signatures in human cancer. Nature 2020, 578, 94–101. [Google Scholar] [CrossRef] [Green Version]
- Henderson, S.; Chakravarthy, A.; Su, X.; Boshoff, C.; Fenton, T.R. APOBEC-mediated cytosine deamination links PIK3CA helical domain mutations to human papillomavirus-driven tumor development. Cell Rep. 2014, 7, 1833–1841. [Google Scholar] [CrossRef] [Green Version]
- Roberts, S.A.; Lawrence, M.S.; Klimczak, L.J.; Grimm, S.A.; Fargo, D.; Stojanov, P.; Kiezun, A.; Kryukov, G.V.; Carter, S.L.; Saksena, G.; et al. An APOBEC cytidine deaminase mutagenesis pattern is widespread in human cancers. Nat. Genet. 2013, 45, 970–976. [Google Scholar] [CrossRef]
- Burns, M.B.; Temiz, N.A.; Harris, R.S. Evidence for APOBEC3B mutagenesis in multiple human cancers. Nat. Genet. 2013, 45, 977–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortez, L.M.; Brown, A.L.; Dennis, M.A.; Collins, C.D.; Brown, A.J.; Mitchell, D.; Mertz, T.M.; Roberts, S.A. APOBEC3A is a prominent cytidine deaminase in breast cancer. PLoS Genet. 2019, 15, e1008545. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.; Roberts, S.A.; Klimczak, L.J.; Sterling, J.F.; Saini, N.; Malc, E.P.; Kim, J.; Kwiatkowski, D.J.; Fargo, D.C.; Mieczkowski, P.A.; et al. An APOBEC3A hypermutation signature is distinguishable from the signature of background mutagenesis by APOBEC3B in human cancers. Nat. Genet. 2015, 47, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Petljak, M.; Dananberg, A.; Chu, K.; Bergstrom, E.N.; Striepen, J.; von Morgen, P.; Chen, Y.; Shah, H.; Sale, J.E.; Alexandrov, L.B.; et al. Mechanisms of APOBEC3 mutagenesis in human cancer cells. Nature 2022, 607, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Nik-Zainal, S.; Wedge, D.C.; Alexandrov, L.B.; Petljak, M.; Butler, A.P.; Bolli, N.; Davies, H.R.; Knappskog, S.; Martin, S.; Papaemmanuil, E.; et al. Association of a germline copy number polymorphism of APOBEC3A and APOBEC3B with burden of putative APOBEC-dependent mutations in breast cancer. Nat. Genet. 2014, 46, 487–491. [Google Scholar] [CrossRef] [Green Version]
- Hashemi, M.; Moazeni-Roodi, A.; Taheri, M. Association of APOBEC3 deletion with cancer risk: A meta-analysis of 26 225 cases and 37 201 controls. Asia-Pac. J. Clin. Oncol. 2019, 15, 275–287. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Qi, Q.; He, Q.; Sun, M.; Wang, S.; Zhou, G.; Sun, Y. APOBEC3 deletion increases the risk of breast cancer: A meta-analysis. Oncotarget 2016, 7, 74979–74986. [Google Scholar] [CrossRef] [Green Version]
- Middlebrooks, C.D.; Banday, A.R.; Matsuda, K.; Udquim, K.I.; Onabajo, O.O.; Paquin, A.; Figueroa, J.D.; Zhu, B.; Koutros, S.; Kubo, M.; et al. Association of germline variants in the APOBEC3 region with cancer risk and enrichment with APOBEC-signature mutations in tumors. Nat. Genet. 2016, 48, 1330–1338. [Google Scholar] [CrossRef]
- Xuan, D.; Li, G.; Cai, Q.; Deming-Halverson, S.; Shrubsole, M.J.; Shu, X.-O.; Kelley, M.C.; Zheng, W.; Long, J. APOBEC3 deletion polymorphism is associated with breast cancer risk among women of European ancestry. Carcinogenesis 2013, 34, 2240–2243. [Google Scholar] [CrossRef] [Green Version]
- Gansmo, L.B.; Romundstad, P.; Hveem, K.; Vatten, L.; Nik-Zainal, S.; Lønning, P.E.; Knappskog, S. APOBEC3A/B deletion polymorphism and cancer risk. Carcinogenesis 2017, 39, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Gansmo, L.B.; Sofiyeva, N.; Bjørnslett, M.; Romundstad, P.; Hveem, K.; Vatten, L.; Dørum, A.; Lønning, P.E.; Knappskog, S. Impact of the APOBEC3A/B deletion polymorphism on risk of ovarian cancer. Sci. Rep. 2021, 11, 23463. [Google Scholar] [CrossRef]
- McCann, J.L.; Klein, M.M.; Leland, E.M.; Law, E.K.; Brown, W.L.; Salamango, D.J.; Harris, R.S. The DNA deaminase APOBEC3B interacts with the cell-cycle protein CDK4 and disrupts CDK4-mediated nuclear import of Cyclin D1. J. Biol. Chem. 2019, 294, 12099–12111. [Google Scholar] [CrossRef] [PubMed]
- Bakhoum, S.F. APOBEC3A drives STING-dependent metastasis. Nat. Cancer 2021, 2, 1293–1295. [Google Scholar] [CrossRef] [PubMed]
- Wörmann, S.M.; Zhang, A.; Thege, F.I.; Cowan, R.W.; Rupani, D.N.; Wang, R.; Manning, S.L.; Gates, C.; Wu, W.; Levin-Klein, R.; et al. APOBEC3A drives deaminase domain-independent chromosomal instability to promote pancreatic cancer metastasis. Nat. Cancer 2021, 2, 1338–1356. [Google Scholar] [CrossRef]
- Kondo, S.; Wakae, K.; Wakisaka, N.; Nakanishi, Y.; Ishikawa, K.; Komori, T.; Moriyama-Kita, M.; Endo, K.; Murono, S.; Wang, Z.; et al. APOBEC3A associates with human papillomavirus genome integration in oropharyngeal cancers. Oncogene 2017, 36, 1687–1697. [Google Scholar] [CrossRef] [PubMed]
- Vartanian, J.-P.; Guétard, D.; Henry, M.; Wain-Hobson, S. Evidence for Editing of Human Papillomavirus DNA by APOBEC3 in Benign and Precancerous Lesions. Science 2008, 320, 230–233. [Google Scholar] [CrossRef] [Green Version]
- Faden, D.L.; Kuhs, K.A.L.; Lin, M.; Langenbucher, A.; Pinheiro, M.; Yeager, M.; Cullen, M.; Boland, J.F.; Steinberg, M.K.; Bass, S.E.; et al. APOBEC Mutagenesis Is Concordant between Tumor and Viral Genomes in HPV-Positive Head and Neck Squamous Cell Carcinoma. Viruses 2021, 13, 1666. [Google Scholar] [CrossRef]
- Lawson, J.S.; Glenn, W.K. Mouse Mammary Tumour Virus (MMTV) in Human Breast Cancer-The Value of Bradford Hill Criteria. Viruses 2022, 14, 721. [Google Scholar] [CrossRef]
- de Sousa Pereira, N.; Akelinghton Freire Vitiello, G.; Karina Banin-Hirata, B.; Scantamburlo Alves Fernandes, G.; José Sparça Salles, M.; Karine Amarante, M.; Angelica Ehara Watanabe, M. Mouse Mammary Tumor Virus (MMTV)-Like env Sequence in Brazilian Breast Cancer Samples: Implications in Clinicopathological Parameters in Molecular Subtypes. Int. J. Environ. Res. Public Health 2020, 17, 9496. [Google Scholar] [CrossRef]
- Vitiello, G.A.F.; de Sousa Pereira, N.; Amarante, M.K.; Banin-Hirata, B.K.; Campos, C.Z.; de Oliveira, K.B.; Losi-Guembarovski, R.; Watanabe, M.A.E. Germline APOBEC3B deletion influences clinicopathological parameters in luminal-A breast cancer: Evidences from a southern Brazilian cohort. J. Cancer Res. Clin. Oncol. 2020, 146, 1523–1532. [Google Scholar] [CrossRef]
- Hammond, M.E.; Hayes, D.F.; Dowsett, M.; Allred, D.C.; Hagerty, K.L.; Badve, S.; Fitzgibbons, P.L.; Francis, G.; Goldstein, N.S.; Hayes, M.; et al. American Society of Clinical Oncology/College Of American Pathologists guideline recommendations for immunohistochemical testing of estrogen and progesterone receptors in breast cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2010, 28, 2784–2795. [Google Scholar] [CrossRef] [Green Version]
- Wolff, A.C.; Hammond, M.E.H.; Allison, K.H.; Harvey, B.E.; Mangu, P.B.; Bartlett, J.M.S.; Bilous, M.; Ellis, I.O.; Fitzgibbons, P.; Hanna, W.; et al. Human Epidermal Growth Factor Receptor 2 Testing in Breast Cancer: American Society of Clinical Oncology/College of American Pathologists Clinical Practice Guideline Focused Update. Arch. Pathol. Lab. Med. 2018, 142, 1364–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brierley, J.D.; Gospodarowicz, M.K.; Wittekind, C. TNM Classification of Malignant Tumours; Wiley: Hoboken, NJ, USA, 2017. [Google Scholar]
- Goldhirsch, A.; Ingle, J.N.; Gelber, R.D.; Coates, A.S.; Thürlimann, B.; Senn, H.J. Thresholds for therapies: Highlights of the St Gallen International Expert Consensus on the primary therapy of early breast cancer 2009. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2009, 20, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Cobo, M.; Melana, S.M.; Holland, J.F.; Pogo, B.G. Transcription profile of a human breast cancer cell line expressing MMTV-like sequences. Infect. Agents Cancer 2006, 1, 7. [Google Scholar] [CrossRef] [Green Version]
- Stavrou, S.; Ross, S.R. APOBEC3 Proteins in Viral Immunity. J. Immunol. 2015, 195, 4565–4570. [Google Scholar] [CrossRef] [Green Version]
- Swanton, C.; McGranahan, N.; Starrett, G.J.; Harris, R.S. APOBEC Enzymes: Mutagenic Fuel for Cancer Evolution and Heterogeneity. Cancer Discov. 2015, 5, 704–712. [Google Scholar] [CrossRef] [Green Version]
- Vieira, V.C.; Leonard, B.; White, E.A.; Starrett, G.J.; Temiz, N.A.; Lorenz, L.D.; Lee, D.; Soares, M.A.; Lambert, P.F.; Howley, P.M.; et al. Human papillomavirus E6 triggers upregulation of the antiviral and cancer genomic DNA deaminase APOBEC3B. mBio 2014, 5, e02234-14. [Google Scholar] [CrossRef] [Green Version]
- Bobrovnitchaia, I.; Valieris, R.; Drummond, R.D.; Lima, J.P.; Freitas, H.C.; Bartelli, T.F.; de Amorim, M.G.; Nunes, D.N.; Dias-Neto, E.; da Silva, I.T. APOBEC-mediated DNA alterations: A possible new mechanism of carcinogenesis in EBV-positive gastric cancer. Int. J. Cancer 2020, 146, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Okeoma, C.M.; Lovšin, N.; Peterlin, B.M.; Ross, S.R. APOBEC3 inhibits mouse mammary tumour virus replication in vivo. Nature 2007, 445, 927–930. [Google Scholar] [CrossRef] [PubMed]
- Okeoma, C.M.; Huegel, A.L.; Lingappa, J.; Feldman, M.D.; Ross, S.R. APOBEC3 proteins expressed in mammary epithelial cells are packaged into retroviruses and can restrict transmission of milk-borne virions. Cell Host Microbe 2010, 8, 534–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacMillan, A.L.; Kohli, R.M.; Ross, S.R. APOBEC3 inhibition of mouse mammary tumor virus infection: The role of cytidine deamination versus inhibition of reverse transcription. J. Virol. 2013, 87, 4808–4817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagen, B.; Kraase, M.; Indikova, I.; Indik, S. A high rate of polymerization during synthesis of mouse mammary tumor virus DNA alleviates hypermutation by APOBEC3 proteins. PLoS Pathog. 2019, 15, e1007533. [Google Scholar] [CrossRef] [Green Version]
- Konstantoulas, C.J.; Lamp, B.; Rumenapf, T.H.; Indik, S. Single amino acid substitution (G42E) in the receptor binding domain of mouse mammary tumour virus envelope protein facilitates infection of non-murine cells in a transferrin receptor 1-independent manner. Retrovirology 2015, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Klonowska, K.; Kluzniak, W.; Rusak, B.; Jakubowska, A.; Ratajska, M.; Krawczynska, N.; Vasilevska, D.; Czubak, K.; Wojciechowska, M.; Cybulski, C.; et al. The 30 kb deletion in the APOBEC3 cluster decreases APOBEC3A and APOBEC3B expression and creates a transcriptionally active hybrid gene but does not associate with breast cancer in the European population. Oncotarget 2017, 8, 76357–76374. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.; Marathe, S.D.; Nain, S.; Nema, V.; Ghate, M.V.; Gangakhedkar, R.R. APOBEC3B deletion impacts on susceptibility to acquire HIV-1 and its advancement among individuals in western India. APMIS 2016, 124, 881–887. [Google Scholar] [CrossRef] [PubMed]
- An, P.; Johnson, R.; Phair, J.; Kirk, G.D.; Yu, X.-F.; Donfield, S.; Buchbinder, S.; Goedert, J.J.; Winkler, C.A. APOBEC3B Deletion and Risk of HIV-1 Acquisition. J. Infect. Dis. 2009, 200, 1054–1058. [Google Scholar] [CrossRef] [Green Version]
- Revathidevi, S.; Manikandan, M.; Rao, A.K.; Vinothkumar, V.; Arunkumar, G.; Rajkumar, K.S.; Ramani, R.; Rajaraman, R.; Ajay, C.; Munirajan, A.K. Analysis of APOBEC3A/3B germline deletion polymorphism in breast, cervical and oral cancers from South India and its impact on miRNA regulation. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2016, 37, 11983–11990. [Google Scholar] [CrossRef]
- Zhang, T.; Cai, J.; Chang, J.; Yu, D.; Wu, C.; Yan, T.; Zhai, K.; Bi, X.; Zhao, H.; Xu, J.; et al. Evidence of associations of APOBEC3B gene deletion with susceptibility to persistent HBV infection and hepatocellular carcinoma. Hum. Mol. Genet. 2012, 22, 1262–1269. [Google Scholar] [CrossRef] [Green Version]
- Cescon, D.W.; Haibe-Kains, B.; Mak, T.W. APOBEC3B expression in breast cancer reflects cellular proliferation, while a deletion polymorphism is associated with immune activation. Proc. Natl. Acad. Sci. USA 2015, 112, 2841–2846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udquim, K.I.; Zettelmeyer, C.; Banday, A.R.; Lin, S.H.; Prokunina-Olsson, L. APOBEC3B expression in breast cancer cell lines and tumors depends on the estrogen receptor status. Carcinogenesis 2020, 41, 1030–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Periyasamy, M.; Patel, H.; Lai, C.F.; Nguyen, V.T.M.; Nevedomskaya, E.; Harrod, A.; Russell, R.; Remenyi, J.; Ochocka, A.M.; Thomas, R.S.; et al. APOBEC3B-Mediated Cytidine Deamination Is Required for Estrogen Receptor Action in Breast Cancer. Cell Rep. 2015, 13, 108–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bos, M.K.; Smid, M.; Sleijfer, S.; Martens, J.W.M. Apolipoprotein B mRNA-Editing Catalytic Polypeptide-Like–Induced Protein Changes in Estrogen Receptor–Positive, Human Epidermal Growth Factor Receptor 2–Negative Breast Cancer Throughout Disease Progression. JCO Precis. Oncol. 2022, 6, e2100190. [Google Scholar] [CrossRef]
- Sammons, S.; Raskina, K.; Danziger, N.; Alder, L.; Schrock, A.B.; Venstrom, J.M.; Knutson, K.L.; Thompson, E.A.; McGregor, K.; Sokol, E.; et al. APOBEC Mutational Signatures in Hormone Receptor–Positive Human Epidermal Growth Factor Receptor 2–Negative Breast Cancers Are Associated With Poor Outcomes on CDK4/6 Inhibitors and Endocrine Therapy. JCO Precis. Oncol. 2022, 6, e2200149. [Google Scholar] [CrossRef] [PubMed]
- André, F.; Ciruelos, E.; Rubovszky, G.; Campone, M.; Loibl, S.; Rugo, H.S.; Iwata, H.; Conte, P.; Mayer, I.A.; Kaufman, B.; et al. Alpelisib for PIK3CA-Mutated, Hormone Receptor-Positive Advanced Breast Cancer. N. Engl. J. Med. 2019, 380, 1929–1940. [Google Scholar] [CrossRef]
- Rezaei, M.; Hashemi, M.; Hashemi, S.M.; Mashhadi, M.A.; Taheri, M. APOBEC3 Deletion is Associated with Breast Cancer Risk in a Sample of Southeast Iranian Population. Int. J. Mol. Cell. Med. 2015, 4, 103–108. [Google Scholar]



{kind=link}
{kind=link}
| APOBEC 3A/B Genotypes | MMTV-like env | |||
|---|---|---|---|---|
| Negative | Positive | χ2 p-Value | Odds Ratio (CI 95%) | |
| WT/WT [n (%)] | 145 (85.8) | 35 (87.5) | 1.00 | 0.87 (0.31–2.46) |
| WT/Del [n (%)] | 24 (14.2) | 5 (12.5) | ||
| Parameter | MMTV-like Positive (n = 40) | MMTV-like Negative (n = 169) | ||||
|---|---|---|---|---|---|---|
| APOBEC3A/B | WT/WT | WT/Del | Tau (p) | WT/WT | WT/Del | Tau (p) |
| Age (years) | ||||||
| Mean (SD) | 60 (14) | 41 (6) | −0.382 (0.009 *) | 56 (13) | 56 (13) | 0.007 (0.906) |
| Median (IQR) | 57 (22) | 41 (7) | 53 (17) | 59 (18) | ||
| <40 [n (%)] | 3 (8.8) | 2 (40.0) | 13 (8.9) | 3 (12.5) | ||
| 40–49 [n (%)] | 7 (20.6) | 3 (60.0) | 41 (28.3) | 5 (20.8) | ||
| 50–59 [n (%)] | 8 (23.5) | 0 (0.0) | 36 (24.8) | 4 (16.6) | ||
| 60–69 [n (%)] | 7 (20.6) | 0 (0.0) | 33 (22.7) | 9 (37.5) | ||
| 70–79 [n (%)] | 7 (20.6) | 0 (0.0) | 16 (11.1) | 2 (8.3) | ||
| ≥80 [n (%)] | 2 (5.9) | 0 (0.0) | 6 (4.2) | 1 (4.3) | ||
| Missed [n (%)] | 1 | 0 | ||||
| Tumor size (cm) | ||||||
| Mean (SD) | 2.9 (2.4) | 3.7 (1.7) | 0.192 (0.186) | 2.8 (1.9) | 2.6 (1.7) | −0.006 (0.918) |
| Median (IQR) | 2.0 (0.7) | 3.5 (1.5) | 2.2 (2.0) | 2.4 (1.4) | ||
| 0–1.5 [n (%)] | 6 (17.1) | 1 (20.0) | 41 (28.7) | 6 (25.0) | ||
| 1.51–3.0 [n (%)] | 21 (60.0) | 1 (20.0) | 62 (43.3) | 14 (58.4) | ||
| >3.0 [n (%)] | 8 (22.9) | 3 (60.0) | 40 (28.0) | 4 (16.6) | ||
| Missed | 1 | 2 | ||||
| Histopathological grade [n (%)] | ||||||
| I | 5 (15.1) | 0 (0.0) | 0.091 (0.504) | 22 (16.1) | 1 (4.8) | 0.043 (0.534) |
| II | 11 (33.4) | 2 (40.0) | 57 (41.6) | 11 (52.4) | ||
| III | 17 (51.5) | 3 (60.0) | 58 (42.3) | 9 (42.8) | ||
| Missed | 2 | 11 | ||||
| Tumor stage [n (%)] | ||||||
| 0 | 2 (7.7) | 0 (0.0) | 0.097 (0.394) | 6 (5.3) | 2 (10.0) | −0.151 (0.044 *) |
| I | 8 (30.8) | 0 (0.0) | 22 (19.3) | 5 (25.0) | ||
| II | 7 (26.9) | 3 (75.0) | 46 (40.4) | 11 (55.0) | ||
| III | 7 (26.9) | 1 (25.0) | 34 (29.7) | 1 (5.0) | ||
| IV | 2 (7.7) | 0 (0.0) | 6 (5.3) | 1 (5.0) | ||
| Missed | 10 | 38 | ||||
| Lymph node metastasis [n (%)] | ||||||
| Positive | 16 (47.1) | 2 (40.0) | −0.047 (0.765) | 68 (49.3) | 7 (30.4) | −0.132 (0.085) |
| Negative | 18 (52.9) | 3 (60.0) | 70 (50.7) | 16 (69.6) | ||
| Missed | 1 | 8 | ||||
| Estrogen receptor [n (%)] | ||||||
| Positive | 27 (79.4) | 5 (100.0) | 0.179 (0.041 *) | 108 (78.3) | 20 (86.9) | 0.075 (0.276) |
| Negative | 7 (20.6) | 0 (0.0) | 30 (21.7) | 3 (13.1) | ||
| Missed | 1 | 8 | ||||
| Progesterone receptor [n (%)] | ||||||
| Positive | 19 (55.9) | 2 (40.0) | −0.107 (0.511) | 74 (53.6) | 13 (56.5) | 0.020 (0.796) |
| Negative | 15 (44.1) | 3 (60.0) | 64 (46.4) | 10 (43.5) | ||
| Missed | 1 | 8 | ||||
| HER2 [n (%)] | ||||||
| Positive | 10 (30.3) | 1 (20.0) | −0.077 (0.605) | 24 (17.9) | 2 (8.7) | −0.088 (0.182) |
| Negative | 23 (69.7) | 4 (80.0) | 110 (82.1) | 21 (91.3) | ||
| Missed | 2 | 12 | ||||
| Ki67 [n (%)] | ||||||
| Low | 9 (29.0) | 0 (0.0) | 0.198 (0.160) | 36 (28.8) | 2 (10.5) | 0.023 (0.713) |
| Intermediate | 13 (42.0) | 2 (50.0) | 48 (38.4) | 13 (68.4) | ||
| High | 9 (29.0) | 2 (50.0) | 41 (32.8) | 4 (21.1) | ||
| Missed | 5 | 25 | ||||
| p53 [n (%)] | ||||||
| Positive | 11 (36.7) | 3 (75.0) | 0.251 (0.173) | 45 (33.8) | 5 (23.8) | −0.073 (0.331) |
| Negative | 19 (63.3) | 1 (25.0) | 88 (66.2) | 16 (76.2) | ||
| Missed | 6 | 15 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Sousa Pereira, N.; Vitiello, G.A.F.; Amarante, M.K. Involvement of APOBEC3A/B Deletion in Mouse Mammary Tumor Virus (MMTV)-like Positive Human Breast Cancer. Diagnostics 2023, 13, 1196. https://doi.org/10.3390/diagnostics13061196
de Sousa Pereira N, Vitiello GAF, Amarante MK. Involvement of APOBEC3A/B Deletion in Mouse Mammary Tumor Virus (MMTV)-like Positive Human Breast Cancer. Diagnostics. 2023; 13(6):1196. https://doi.org/10.3390/diagnostics13061196
Chicago/Turabian Stylede Sousa Pereira, Nathália, Glauco Akelinghton Freire Vitiello, and Marla Karine Amarante. 2023. "Involvement of APOBEC3A/B Deletion in Mouse Mammary Tumor Virus (MMTV)-like Positive Human Breast Cancer" Diagnostics 13, no. 6: 1196. https://doi.org/10.3390/diagnostics13061196