
Role of Cyanobacterial Exopolysaccharides in Phototrophic Biofilms and in Complex Microbial Mats

Abstract
:1. Introduction
1.1. Cyanobacterial EPSs: Chemical and Physical Characteristics


1.2. Factors Affecting EPS Synthesis in Cyanobacteria
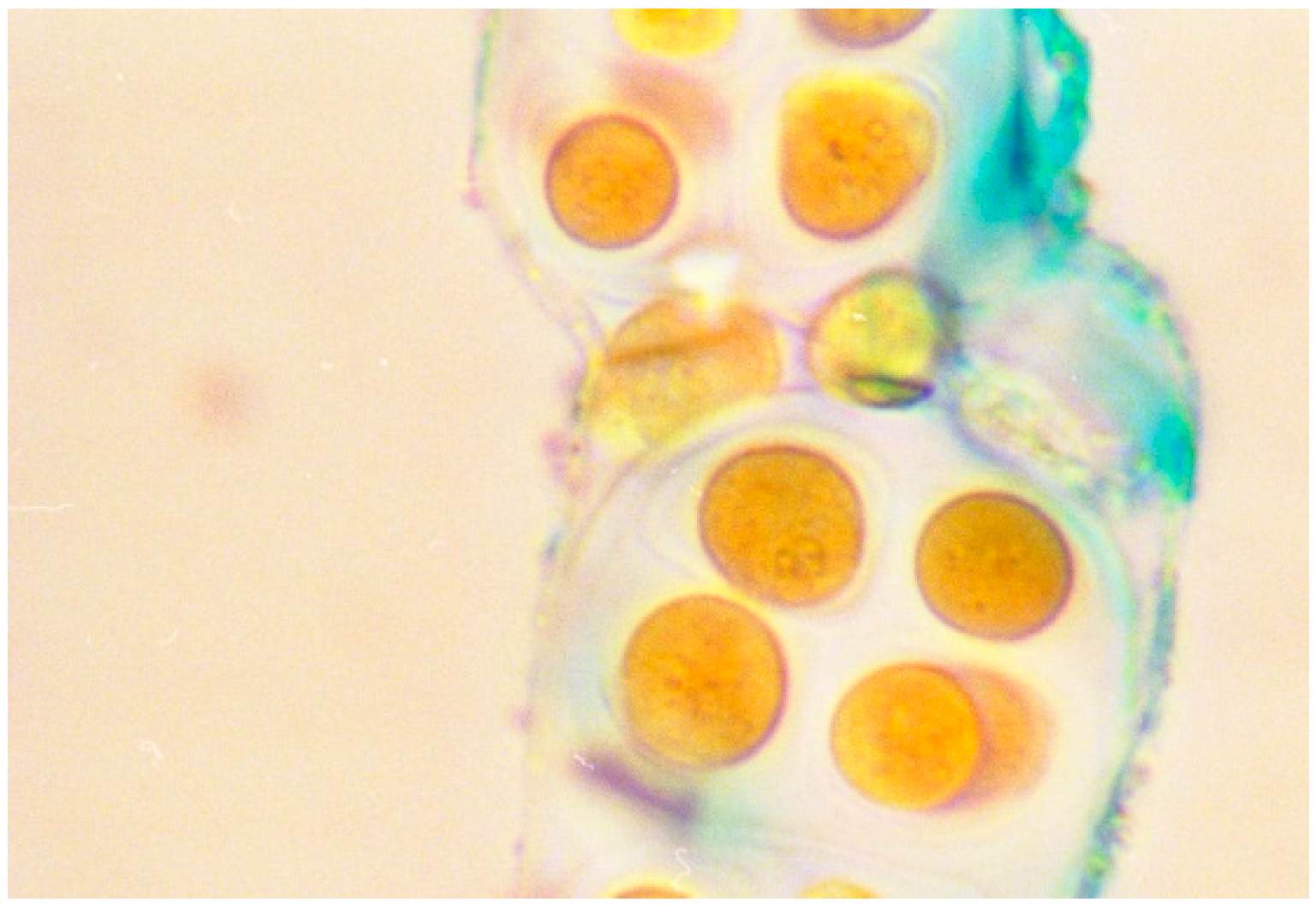
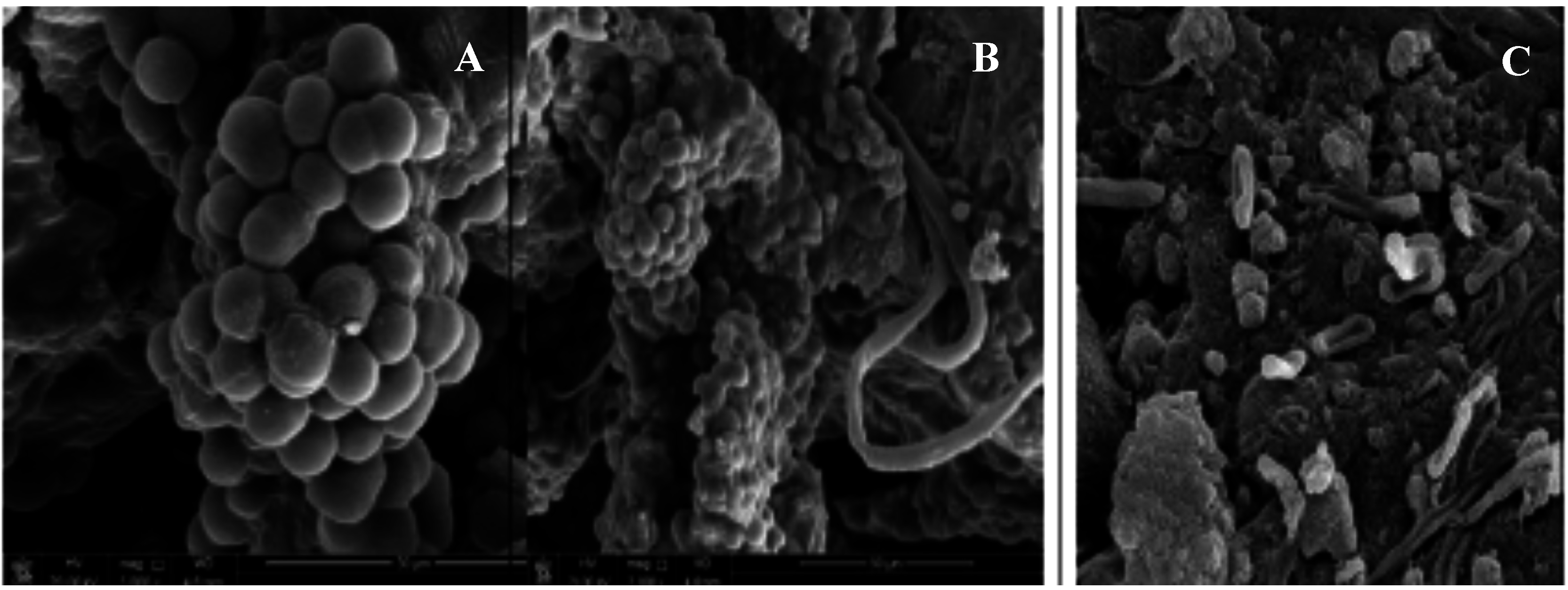
2. Role of Cyanobacterial EPSs in Microbial Mats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Role | Notes | Section |
|---|---|---|
| Adhesion | The adhesion is determined by the hydrophobic characteristics of the EPSs | 2.1 |
| Structure | The extracellular polymeric matrix (EPM) constituted by EPS gives structure to the soil pore system and determines soil particle organization | 2.2 |
| Protection against a-biotic stresses | EPSs counteract water stress, UV stress, physical and chemical stresses | 2.3 |
| Bioweathering processes | EPSs promote mineral concentration and induce pore cracking by drying/swelling cycles | 2.4 |
| Gliding motility | EPSs excretion is involved in cell propulsion | 2.5 |
| Nutrient repositories | EPSs contribute to the concentration of nutrients; EPS constitute a C source for heterotrophic microorganisms | 2.6 |

2.1. Role of EPSs in Cyanobacterial Adhesion to Solid Surfaces and Particles
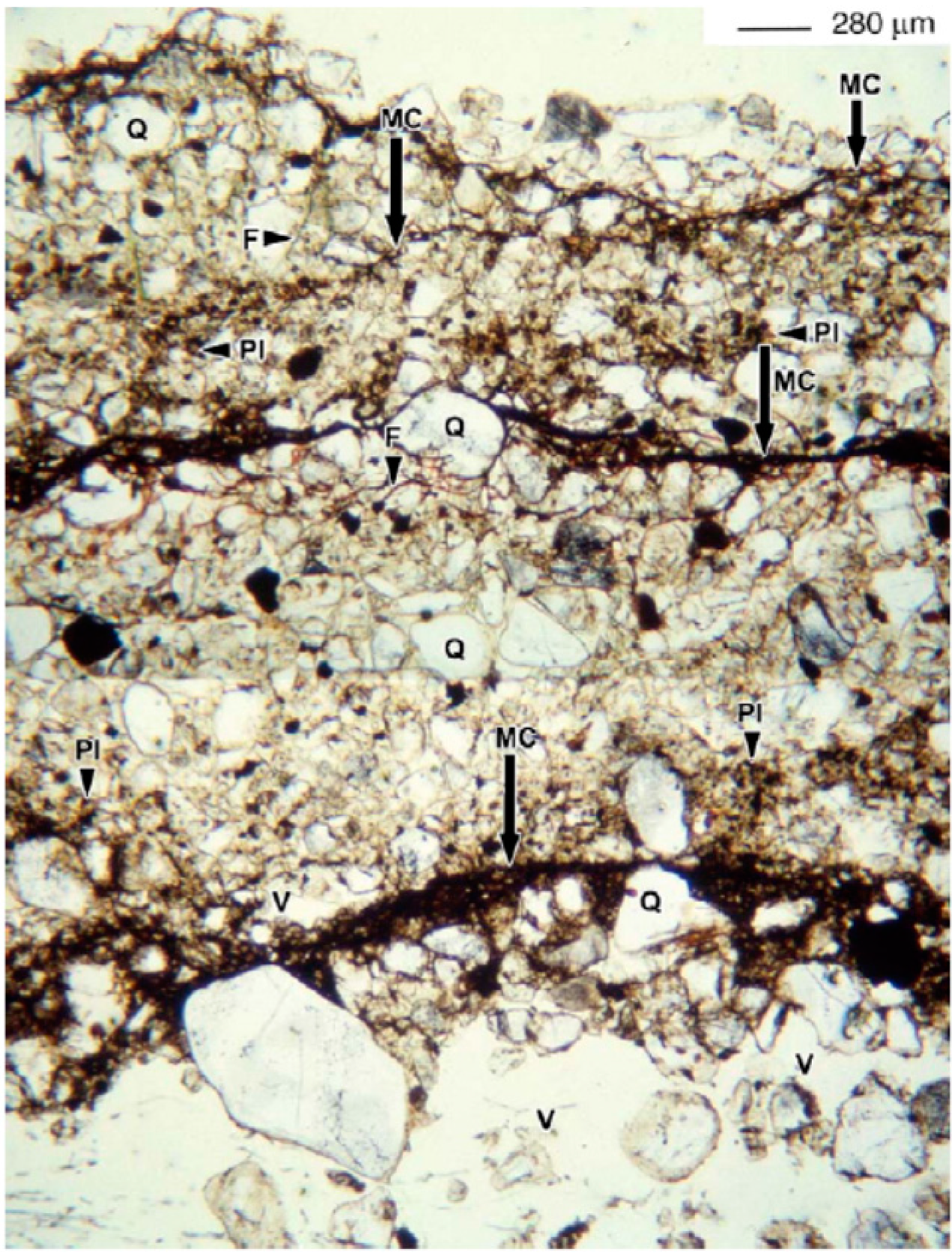
2.2. The Role of EPSs in Biological Soil Crusts



2.3. The Role of EPSs in Tolerance to Water Stress and UV Radiation

2.4. Role of EPSs in Bio-Weathering Processes

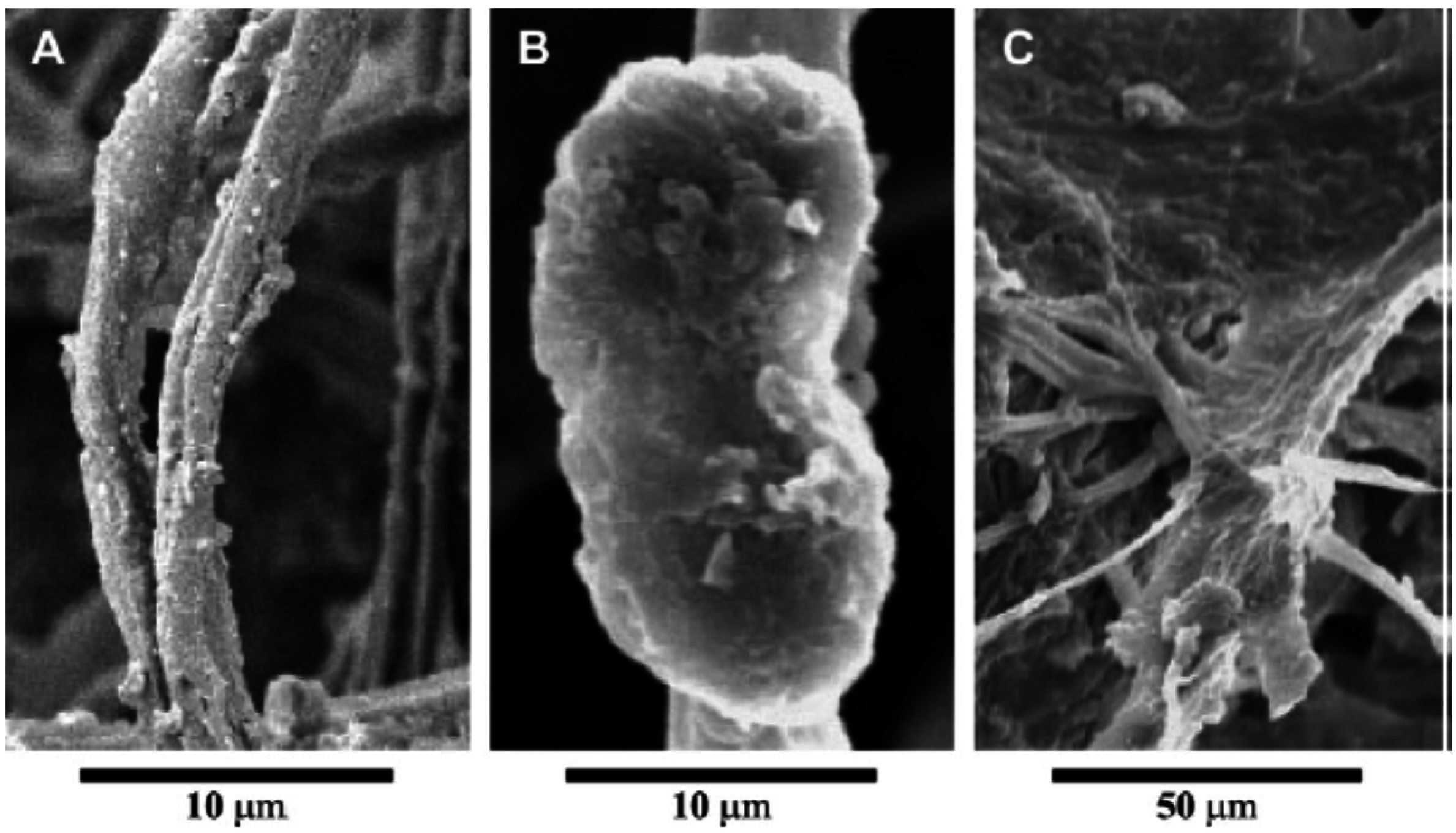
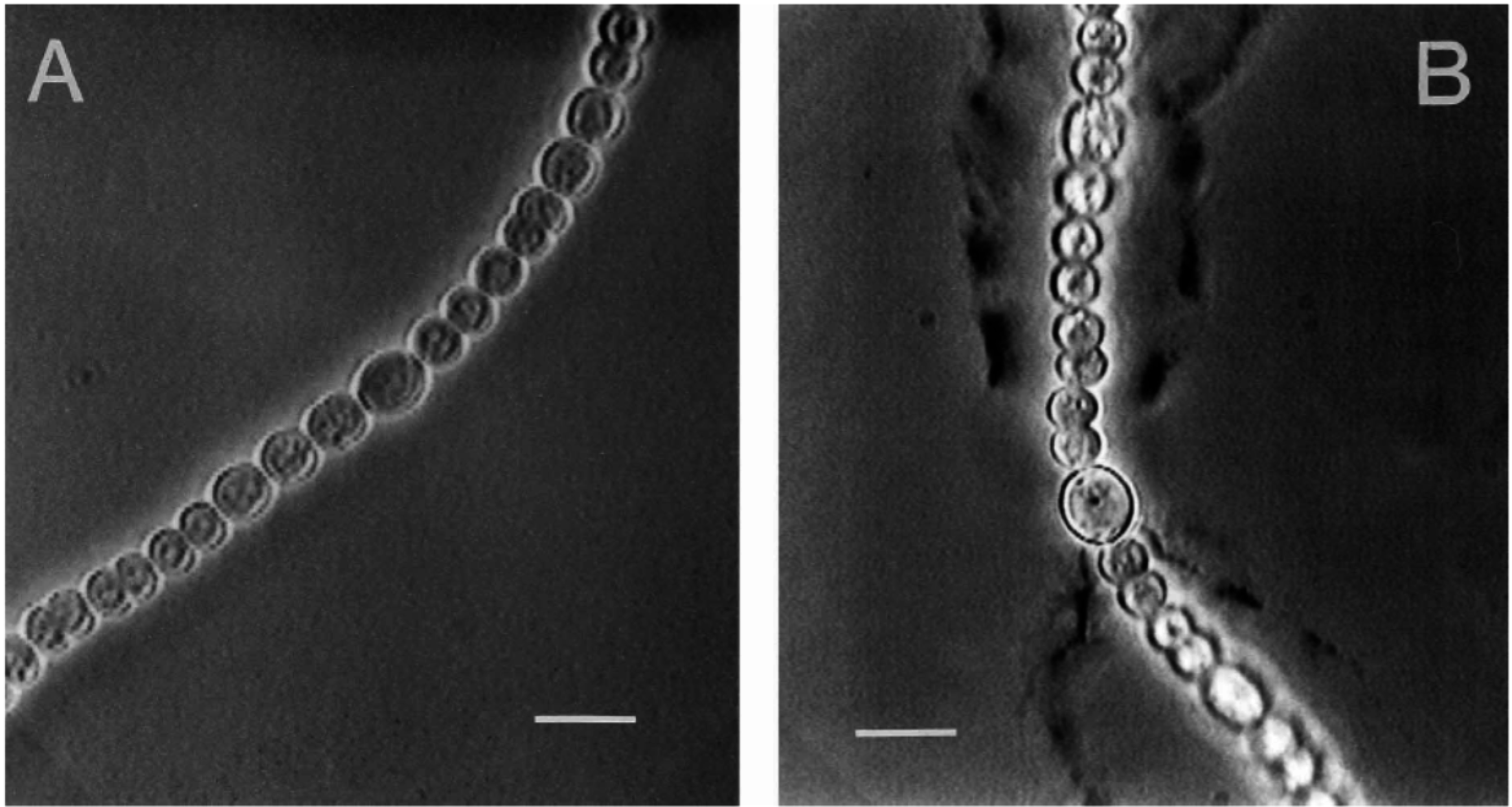
2.5. Role of EPSs in Cyanobacterial Gliding Motility
2.6. Cyanobacterial EPSs as a C-Source
3. Conclusions
Acknowledgments
Conflicts of Interest
References
- Fogg, G.E. The comparative physiology and biochemistry of the blue-green algae. Bacteriol. Rev. 1956, 20, 148–165. [Google Scholar] [PubMed]
- Montoya, H. Algal and cyanobacterial saline biofilms of the Grande Coastal Lagoon, Lima, Peru. Nat. Resour. Environ. Issues 2009, 15, 127–134. [Google Scholar]
- Pereira, S.; Zille, A.; Micheletti, E.; Moradas-Ferreira, P.; de Philippis, R.; Tamagnini, P. Complexity of cyanobacterial exopolysaccharides: Composition, structures, inducing factors and putative genes involved in their biosynthesis and assembly. FEMS Microbiol. Rev. 2009, 33, 917–941. [Google Scholar] [CrossRef] [PubMed]
- De Philippis, R.; Vincenzini, M. Exocellular polysaccharides from cyanobacteria and their possible applications. FEMS Microbiol. Rev. 1998, 22, 151–175. [Google Scholar]
- Hill, D.R.; Keenan, T.W.; Helm, R.F.; Potts, M.; Crowe, L.M.; Crowe, J.H. Extracellular polysaccharide of Nostoc commune (Cyanobacteria) inhibits fusion of membrane vesicles during desiccation. J. Appl. Phycol. 1997, 9, 237–248. [Google Scholar] [CrossRef]
- Pereira, S.B.; Mota, R.; Santos, C.L.; de Philippis, R.; Tamagnini, P. Assembly and Export of Extracellular Polymeric Substances (EPS) in Cyanobacteria. Adv. Bot. Res. 2013, 65, 235–279. [Google Scholar]
- Sekar, S.; Paulraj, P. Strategic mining of cyanobacterial patents from the USPTO patent database and analysis of their scope and implications. J. Appl. Phycol. 2007, 19, 277–292. [Google Scholar] [CrossRef]
- Mager, D.M.; Thomas, A.D. Carbohydrates in cyanobacterial soil crusts as a source of carbon in the southwest Kalahari, Botswana. Soil Biol. Biochem. 2010, 42, 313–318. [Google Scholar] [CrossRef]
- Sahlan Ozturk, B.A. Relationship between chromium (VI) resistance and extracellular polymeric substances (EPS) concentration by some cyanobacterial isolates. Environ. Sci. Pollut. Res. Int. 2008, 15, 478–480. [Google Scholar]
- Priester, J.H.; Olson, S.G.; Webb, S.M.; Neu, M.P.; Hersman, L.E.; Holden, P.A. Enhanced exopolymer production and chromium stabilization in Pseudomonas putida unsaturated biofilms. Appl. Environ. Microbiol. 2006, 72, 1988–1996. [Google Scholar] [CrossRef] [PubMed]
- Bar-Zeev, E.; Berman-Frank, I.; Stambler, N.; Vázquez Domínguez, E.; Zohary, T.; Capuzzo, E.; Meeder, E.; Suggett, D.J.; Iluz, D.; Dishon, G.; et al. Transparent exopolymer particles (TEP) link phytoplankton and bacterial production in the Gulf of Aqaba. Aquat. Microb. Ecol. 2009, 56, 217–225. [Google Scholar] [CrossRef]
- Micheletti, E.; Pereira, S.; Mannelli, F.; Moradas-Ferreira, P.; Tamagnini, P.; de Philippis, R. Sheathless Mutant of Cyanobacterium Gloeothece. sp. Strain PCC 6909 with Increased Capacity To Remove Copper Ions from Aqueous Solutions. Appl. Environ. Microbiol. 2008, 74, 2797–2804. [Google Scholar] [CrossRef] [PubMed]
- Vincenzini, M.; de Philippis, R.; Sili, C.; Materassi, R. Studies on exopolysaccharide release by diazotrophic batch cultures of Cyanospira. capsulata. Appl. Microbiol. Biotechnol. 1990, 34, 392–396. [Google Scholar] [CrossRef]
- Di Pippo, F.; Ellwood, N.W.; Gismondi, A.; Bruno, L.; Rossi, F.; Magni, P.; de Philippis, R. Characterization of exopolysaccharides produced by seven biofilm-forming cyanobacterial strains for biotechnological applications. J. Appl. Phycol. 2013, 25, 1697–1708. [Google Scholar] [CrossRef] [Green Version]
- De Philippis, R.; Colica, G.; Micheletti, E. Exopolysaccharide-producing cyanobacteria in heavy metal removal from water: Molecular basis and practical applicability of the biosorption process. Appl. Microbiol. Biotechnol. 2011, 92, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, R.; Rockey, J.; Sutherland, I.W.; Roller, S. Novel bioemulsifiers from microorganisms for use in foods. J. Biotechnol. 1995, 40, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; de Philippis, R. Exocellular Polysaccharides in Microalgae and Cyanobacteria: Chemical Features, Roles and Enzymes and Genes Involved in Their Biosynthesis; Borowitzka, M., Beardall, J., Raven, J.A., Eds.; Springer: Berlin, Germany, 2015. [Google Scholar]
- Hu, C.; Liu, Y.; Paulsen, B.S.; Petersen, D.; Klaveness, D. Extracellular carbohydrate polymers from five desert soil algae with different cohesion in the stabilization of fine sand grain. Carbohydr. Polym. 2003, 54, 33–42. [Google Scholar] [CrossRef]
- Li, P.; Harding, S.E.; Liu, Z. Cyanobacterial exopolysaccharides: Their nature and potential biotechnological applications. Biotechnol. Genet. Eng. Rev. 2001, 18, 375–404. [Google Scholar] [CrossRef] [PubMed]
- Forni, C.; Telo’, F.R.; Caiola, M.G. Comparative analysis of the polysaccharides produced by different species of Microcystis (Chroococcales, Cyanophyta). Phycologia 1997, 36, 181–185. [Google Scholar] [CrossRef]
- Mouhim, R.F.; Cornet, J.-F.; Fontane, T.; Fournet, B.; Dubertret, G. Production, isolation and preliminary characterization of the exopolysaccharide of the cyanobacterium Spirulina platensis. Biotechnol. Lett. 1993, 15, 567–572. [Google Scholar] [CrossRef]
- Gloaguen, V.; Morvan, H.; Hoffmann, L.; Plancke, Y.; Wieruszeski, J.-M.; Lippens, G.; Strecker, G. Capsular polysaccharide produced by the thermophilic cyanobacterium Mastigocladus laminosus. Structural study of an undecasaccharide obtained by lithium degradation. Eur. J. Biochem. FEBS 1999, 266, 762–770. [Google Scholar] [CrossRef]
- Garozzo, D.; Impallomeni, G.; Spina, E.; Sturiale, L. The structure of the exocellular polysaccharide from the cyanobacterium Cyanospira capsulata. Carbohydr. Res. 1998, 307, 113–124. [Google Scholar] [CrossRef] [PubMed]
- De Philippis, R.; Margheri, M.C.; Materassi, R.; Vincenzini, M. Potential of unicellular cyanobacteria from saline environments as exopolysaccharide producers. Appl. Environ. Microbiol. 1998, 64, 1130–1132. [Google Scholar] [PubMed]
- Nicolaus, B.; Panico, A.; Lama, L.; Romano, I.; Manca, M.C.; de Giulio, A.; Gambacorta, A. Chemical composition and production of exopolysaccharides from representative members of heterocystous and non-heterocystous cyanobacteria. Phytochemistry 1999, 52, 639–647. [Google Scholar] [CrossRef]
- Otero, A.; Vincenzini, M. Nostoc (Cyanophyceae) goes nude: Extracellular polysaccharides serve as a sink for reducing power under unbalanced C/N metabolism. J. Phycol. 2004, 40, 74–81. [Google Scholar] [CrossRef]
- Izumo, A.; Fujiwara, S.; Oyama, Y.; Satoh, A.; Fujita, N.; Nakamura, Y.; Tsuzuki, M. Physicochemical properties of starch in Chlorella change depending on the CO2 concentration during growth: Comparison of structure and properties of pyrenoid and stroma starch. Plant Sci. 2007, 172, 1138–1147. [Google Scholar] [CrossRef]
- Sohm, J.A.; Edwards, B.R.; Wilson, B.G.; Webb, E.A. Constitutive Extracellular Polysaccharide (EPS) Production by Specific Isolates of Crocosphaera watsonii. Front. Microbiol. 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Austin, A.; Yahdjian, L.; Stark, J.; Belnap, J.; Porporato, A.; Norton, U.; Ravetta, D.; Schaeffer, S. Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 2004, 141, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.; Vargas, M.A.; Olivares, H.; Rivas, J.; Guerrero, M.G. Exopolysaccharide production by the cyanobacterium Anabaena sp. ATCC 33047 in batch and continuous culture. J. Biotechnol. 1998, 60, 175–182. [Google Scholar] [CrossRef]
- De Philippis, R.; Margheri, M.C.; Pelosi, E.; Ventura, S. Exopolysaccharide production by a unicellular cyanobacterium isolated from a hypersaline habitat. J. Appl. Phycol. 1993, 5, 387–394. [Google Scholar] [CrossRef]
- Liu, H.; Buskey, E.J. Hypersalinity enhances the production of extracellular polymeric substance (EPS) in the Texas brown tide alga, Aureoumbra lagunensis (Pelagophyceae). J. Phycol. 2000, 36, 71–77. [Google Scholar] [CrossRef]
- Mota, R.; Pereira, S.B.; Meazzini, M.; Fernandes, R.; Santos, A.; Evans, C.A.; de Philippis, R.; Wright, P.C.; Tamagnini, P. Effects of heavy metals on Cyanothece sp. CCY 0110 growth, extracellular polymeric substances (EPS) production, ultrastructure and protein profiles. J. Proteomics 2015, 120, 75–94. [Google Scholar] [CrossRef] [PubMed]
- De Philippis, R.; Sili, C.; Vincenzini, M. Response of an exopolysaccharide-producing heterocystous cyanobacterium to changes in metabolic carbon flux. J. Appl. Phycol. 1996, 8, 275–281. [Google Scholar] [CrossRef]
- Lars, P.; Brull, Z.H. Studies of polysaccharides from three edible species of Nostoc (Cyanobacteria) with different colony morphologies: Structural characterization and effect on the complement system of polysaccharides from Nostoc commune. J. Phycol. 2000, 36, 871–881. [Google Scholar] [CrossRef]
- Bellezza, S.; Paradossi, G.; de Philippis, R.; Albertano, P. Leptolyngbya strains from Roman hypogea: Cytochemical and physico-chemical characterisation of exopolysaccharides. J. Appl. Phycol. 2003, 15, 193–200. [Google Scholar] [CrossRef]
- Roeselers, G.; Norris, T.B.; Castenholz, R.W.; Rysgaard, S.; Glud, R.N.; Kühl, M.; Muyzer, G. Diversity of phototrophic bacteria in microbial mats from Arctic hot springs (Greenland). Environ. Microbiol. 2007, 9, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Micheletti, E.; Bruno, L.; Adhikary, S.P.; Albertano, P.; de Philippis, R. Characteristics and role of the exocellular polysaccharides produced by five cyanobacteria isolated from phototrophic biofilms growing on stone monuments. Biofouling 2012, 28, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belnap, J.; Laxalt, M.; Peterson, P. Biological Soil Crusts: Ecology and Management; US Department of the Interior, Bureau of Land Management, National Science and Technology Center, Information and Communications Group: Washington, DC, USA, 2001. [Google Scholar]
- Allison, D.G.; Sutherland, I.W. The role of exopolysaccharides in adhesion of freshwater bacteria. J. Gen. Microbiol. 1987, 133, 1319–1327. [Google Scholar]
- De Caire, G.Z.; de Cano, M.S.; de Mulé, M.C.Z.; Palma, R.M.; Colombo, K. Exopolysaccharide of Nostoc muscorum (Cyanobacteria) in the aggregation of soil particles. J. Appl. Phycol. 1997, 9, 249–253. [Google Scholar] [CrossRef]
- Neu, T.R.; Marshall, K.C. Bacterial polymers: Physicochemical aspects of their interactions at interfaces. J. Biomater. Appl. 1990, 5, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Henriques Vieira, A.A.; Coelho Ortolano, P.I.; Giroldo, D.; Dellamano Oliveira, M.J.; Bittar, T.B.; Lombardi, A.T.; Sartori, A.L.; Paulsen, B.S. Role of hydrophobic extracellular polysaccharide of Aulacoseira granulata (Bacillariophyceae) on aggregate formation in a turbulent and hypereutrophic reservoir. Limnol. Oceanogr. 2008, 53, 1887–1899. [Google Scholar] [CrossRef]
- Chug, R.; Mathur, S. Extracellular Polymeric Substances from Cyanobacteria: Characteristics, Isolation and Biotechnological Applications-A Review. Int. J. Adv. Eng. Sci. Technol. 2013, 3, 49–53. [Google Scholar]
- Fattom, A.; Shilo, M. Production of emulcyan by Phormidium J-1: Its activity and function. FEMS Microbiol. Lett. 1985, 31, 3–9. [Google Scholar] [CrossRef]
- Bar-Or, Y.; Shilo, M. The role of cell-bound flocculants in coflocculation of benthic cyanobacteria with clay particles. FEMS Microbiol. Ecol. 1988, 4, 169–174. [Google Scholar] [CrossRef]
- Yallop, M.L.; de Winder, B.; Paterson, D.M.; Stal, L.J. Comparative structure, primary production and biogenic stabilization of cohesive and non-cohesive marine sediments inhabited by microphytobenthos. Estuar. Coast Shelf Sci. 1994, 39, 565–582. [Google Scholar] [CrossRef]
- Stal, L.J. Microphytobenthos, their Extracellular Polymeric Substances, and the Morphogenesis of Intertidal Sediments. Geomicrobiol. J. 2003, 20, 463–478. [Google Scholar] [CrossRef]
- Kuske, C.R.; Yeager, C.M.; Johnson, S.; Ticknor, L.O.; Belnap, J. Response and resilience of soil biocrust bacterial communities to chronic physical disturbance in arid shrublands. ISME J. 2012, 6, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Desert Restoration Hub. Biological Soil Crusts–Ecology and Management US Bureau of Land Management Technical Reference 1730-2, 2001–Belnap et al. Available online: http://desertrestorationhub.com/publications/soils/publications/biological-soil-crusts-ecology-and-management-us-bureau-of-land-management-technical-reference-1730-2-2001-belnap-et-al/ (accessed on 24 March 2015).
- Chen, L.; Xie, Z.; Hu, C.; Li, D.; Wang, G.; Liu, Y. Man-made desert algal crusts as affected by environmental factors in Inner Mongolia, China. J. Arid Environ. 2006, 67, 521–527. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Y.; Li, D.; Hu, C.; Rao, B. Feasibility of cyanobacterial inoculation for biological soil crusts formation in desert area. Soil Biol. Biochem. 2009, 41, 926–929. [Google Scholar] [CrossRef]
- Colica, G.; Li, H.; Rossi, F.; Li, D.; Liu, Y.; de Philippis, R. Microbial secreted exopolysaccharides affect the hydrological behavior of induced biological soil crusts in desert sandy soils. Soil Biol. Biochem. 2014, 68, 62–70. [Google Scholar] [CrossRef]
- Malam Issa, O.; Défarge, C.; Trichet, J.; Valentin, C.; Rajot, J.L. Microbiotic soil crusts in the Sahel of Western Niger and their influence on soil porosity and water dynamics. Catena 2009, 77, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Decho, A.W. Microbial exopolymer secretions in ocean environments: Their role(s) in food webs and marine processes. Ocean Mar. Biol. Annu. Rev. 1990, 28, 73–153. [Google Scholar]
- Redmile-Gordon, M.A.; Brookes, P.C.; Evershed, R.P.; Goulding, K.W.T.; Hirsch, P.R. Measuring the soil-microbial interface: Extraction of extracellular polymeric substances (EPS) from soil biofilms. Soil Biol. Biochem. 2014, 72, 163–171. [Google Scholar] [CrossRef]
- Rossi, F.; Potrafka, R.M.; Pichel, F.G.; de Philippis, R. The role of the exopolysaccharides in enhancing hydraulic conductivity of biological soil crusts. Soil Biol. Biochem. 2012, 46, 33–40. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Paulsen, B.S.; Klaveness, D. Studies on Polysaccharides from Three Edible Species of Nostoc (Cyanobacteria) with Different Colony Morphologies: Comparison of Monosaccharide Compositions and Viscosities of Polysaccharides from Field Colonies and Suspension Cultures. J. Phycol. 1998, 34, 962–968. [Google Scholar] [CrossRef]
- Hu, C.; Zhang, D.; Huang, Z.; Liu, Y. The vertical microdistribution of cyanobacteria and green algae within desert crusts and the development of the algal crusts. Plant Soil. 2003, 257, 97–111. [Google Scholar] [CrossRef]
- Potts, M. Mechanisms of desiccation tolerance in cyanobacteria. Eur. J. Phycol. 1999, 34, 319–328. [Google Scholar] [CrossRef]
- Knowles, E.J.; Castenholz, R.W. Effect of exogenous extracellular polysaccharides on the desiccation and freezing tolerance of rock-inhabiting phototrophic microorganisms. FEMS Microbiol. Ecol. 2008, 66, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Brussaard, C.P.; Marie, D.; Thyrhaug, R.; Bratbak, G. Flow cytometric analysis of phytoplankton viability following viral infection. Aquat. Microb. Ecol. 2001, 26, 157–166. [Google Scholar] [CrossRef]
- Wierzchos, J.; De Los Ríos, A.; Sancho, L.G.; Ascaso, C. Viability of endolithic micro-organisms in rocks from the McMurdo Dry Valleys of Antarctica established by confocal and fluorescence microscopy. J. Microsc. 2004, 216, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Belnap, J.; Gardner, J.S. Soil microstructure in soils of the Colorado Plateau: The role of the cyanobacterium Microcoleus vaginatus. West N. Am. Nat. 1993, 53, 40–47. [Google Scholar]
- Satoh, K.; Hirai, M.; Nishio, J.; Yamaji, T.; Kashino, Y.; Koike, H. Recovery of photosynthetic systems during rewetting is quite rapid in a terrestrial cyanobacterium, Nostoc commune. Plant Cell Physiol. 2002, 43, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Tamaru, Y.; Takani, Y.; Yoshida, T.; Sakamoto, T. Crucial Role of Extracellular Polysaccharides in Desiccation and Freezing Tolerance in the Terrestrial Cyanobacterium Nostoc commune. Appl. Environ. Microbiol. 2005, 71, 7327–7333. [Google Scholar] [CrossRef] [PubMed]
- Belnap, J.; Büdel, B.; Lange, O.L. Biological Soil Crusts: Characteristics and Distribution. In Biological Soil Crusts: Structure, Function, and Management; Belnap, P.D.J., Lange, P.D.D., Otto, L., Eds.; Springer: Heidelberg, Germany, 2001; pp. 3–30. [Google Scholar]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [PubMed]
- Sakamoto, T.; Kumihashi, K.; Kunita, S.; Masaura, T.; Inoue-Sakamoto, K.; Yamaguchi, M. The extracellular-matrix-retaining cyanobacterium Nostoc verrucosum accumulates trehalose, but is sensitive to desiccation. FEMS Microbiol. Ecol. 2011, 77, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Clegg, J.S. Cryptobiosis—A peculiar state of biological organization. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2001, 128, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Bilger, W.; Scherer, S. UV-B-induced synthesis of photoprotective pigments and extracellular polysaccharides in the terrestrial cyanobacterium Nostoc commune. J. Bacteriol. 1997, 179, 1940–1945. [Google Scholar] [PubMed]
- Garcia-Pichel, F.; Castenholz, R.W. Characterization and Biological Implications of Scytonemin, a Cyanobacterial Sheath Pigment1. J. Phycol. 1991, 27, 395–409. [Google Scholar] [CrossRef]
- Hill, D.R.; Peat, A.; Potts, M. Biochemistry and structure of the glycan secreted by desiccation-tolerant Nostoc commune (Cyanobacteria). Protoplasma 1994, 182, 126–148. [Google Scholar] [CrossRef]
- Garcia-Pichel, F.; Castenholz, R.W. Occurrence of UV-Absorbing, Mycosporine-Like Compounds among Cyanobacterial Isolates and an Estimate of Their Screening Capacity. Appl. Environ. Microbiol. 1993, 59, 163–169. [Google Scholar] [PubMed]
- Wakefield, R.D.; Jones, M.S. An introduction to stone colonizing micro-organisms and biodeterioration of building stone. Q. J. Eng. Geol. Hydrogeol. 1998, 31, 301–313. [Google Scholar] [CrossRef]
- Pointing, S.B.; Belnap, J. Microbial colonization and controls in dryland systems. Nat. Rev. Microbiol. 2012, 10, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Welch, S.A.; Vandevivere, P. Effect of microbial and other naturally occurring polymers on mineral dissolution. Geomicrobiol. J. 1994, 12, 227–238. [Google Scholar] [CrossRef]
- Song, B.; Leff, L.G. Influence of magnesium ions on biofilm formation by Pseudomonas fluorescens. Microbiol. Res. 2006, 161, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, M. Attachment of Pseudomonas fluorescens to glass and influence of electrolytes on bacterium-substratum separation distance. J. Bacteriol. 1988, 170, 2027–2030. [Google Scholar] [PubMed]
- Al-Thani, R.F. Hypolithic cyanobacteria colonization of quartz at south desert. Online J. Biol. Sci. 2014, 14, 57–63. [Google Scholar] [CrossRef]
- De Philippis, R.; Paperi, R.; Sili, C. Heavy metal sorption by released polysaccharides and whole cultures of two exopolysaccharide-producing cyanobacteria. Biodegradation 2007, 18, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Jahn, E. Beiträge zur Botanischen Protistologie; Gebrüder Borntraeger: Leipzig, Germany, 1924. [Google Scholar]
- Hoiczyk, E.; Baumeister, W. The junctional pore complex, a prokaryotic secretion organelle, is the molecular motor underlying gliding motility in cyanobacteria. Curr. Biol. 1998, 8, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Wolgemuth, C.W.; Oster, G. The Junctional Pore Complex and the Propulsion of Bacterial Cells. J. Mol. Microbiol. Biotechnol. 2004, 7, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Hoiczyk, E. Structural and biochemical analysis of the sheath of Phormidium uncinatum. J. Bacteriol. 1998, 180, 3923–3932. [Google Scholar] [PubMed]
- Thomas, A.D.; Hoon, S.R.; Linton, P.E. Carbon dioxide fluxes from cyanobacteria crusted soils in the Kalahari. Appl. Soil Ecol. 2008, 39, 254–263. [Google Scholar] [CrossRef]
- Miralles, I.; Domingo, F.; Cantón, Y.; Trasar-Cepeda, C.; Leirós, M.C.; Gil-Sotres, F. Hydrolase enzyme activities in a successional gradient of biological soil crusts in arid and semi-arid zones. Soil Biol. Biochem. 2012, 53, 124–132. [Google Scholar] [CrossRef]
- Chen, L.; Rossi, F.; Deng, S.; Liu, Y.; Wang, G.; Adessi, A.; De Philippis, R. Macromolecular and chemical features of the excreted extracellular polysaccharides in induced biological soil crusts of different ages. Soil Biol. Biochem. 2014, 78, 1–9. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, F.; De Philippis, R. Role of Cyanobacterial Exopolysaccharides in Phototrophic Biofilms and in Complex Microbial Mats. Life 2015, 5, 1218-1238. https://doi.org/10.3390/life5021218
Rossi F, De Philippis R. Role of Cyanobacterial Exopolysaccharides in Phototrophic Biofilms and in Complex Microbial Mats. Life. 2015; 5(2):1218-1238. https://doi.org/10.3390/life5021218
Chicago/Turabian StyleRossi, Federico, and Roberto De Philippis. 2015. "Role of Cyanobacterial Exopolysaccharides in Phototrophic Biofilms and in Complex Microbial Mats" Life 5, no. 2: 1218-1238. https://doi.org/10.3390/life5021218