
Regulation of CO2 Concentrating Mechanism in Cyanobacteria

Abstract
:1. General Description of Function and Components of the Cyanobacterial CCM

2. Metabolomic Investigations of Carbon Metabolism and How It Pertains to the Ci Acquisition Mechanisms
3. Regulatory Signals that Cause the Activation or Repression of CCM Genes
3.1. Transcriptional Regulation

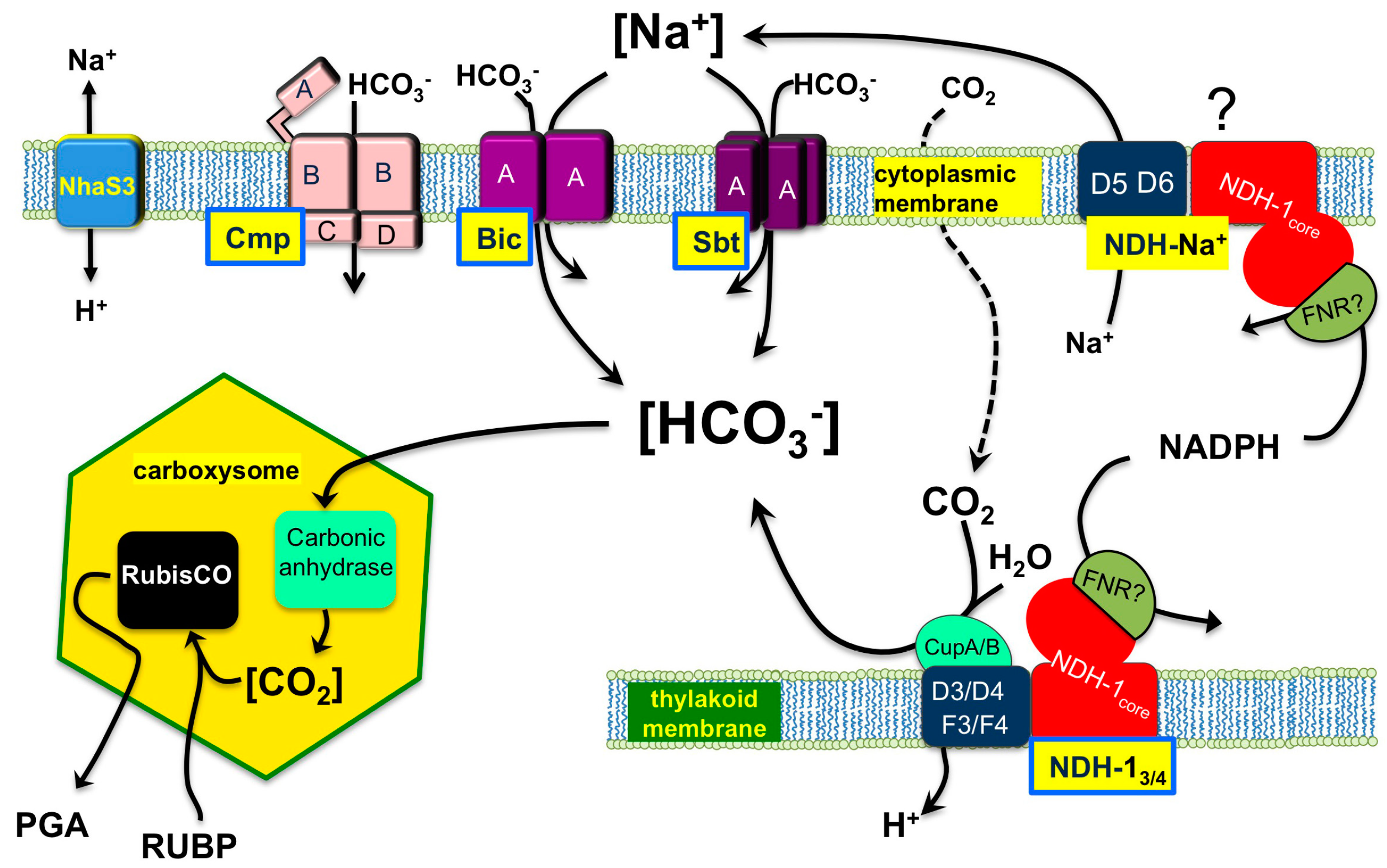{kind=link}
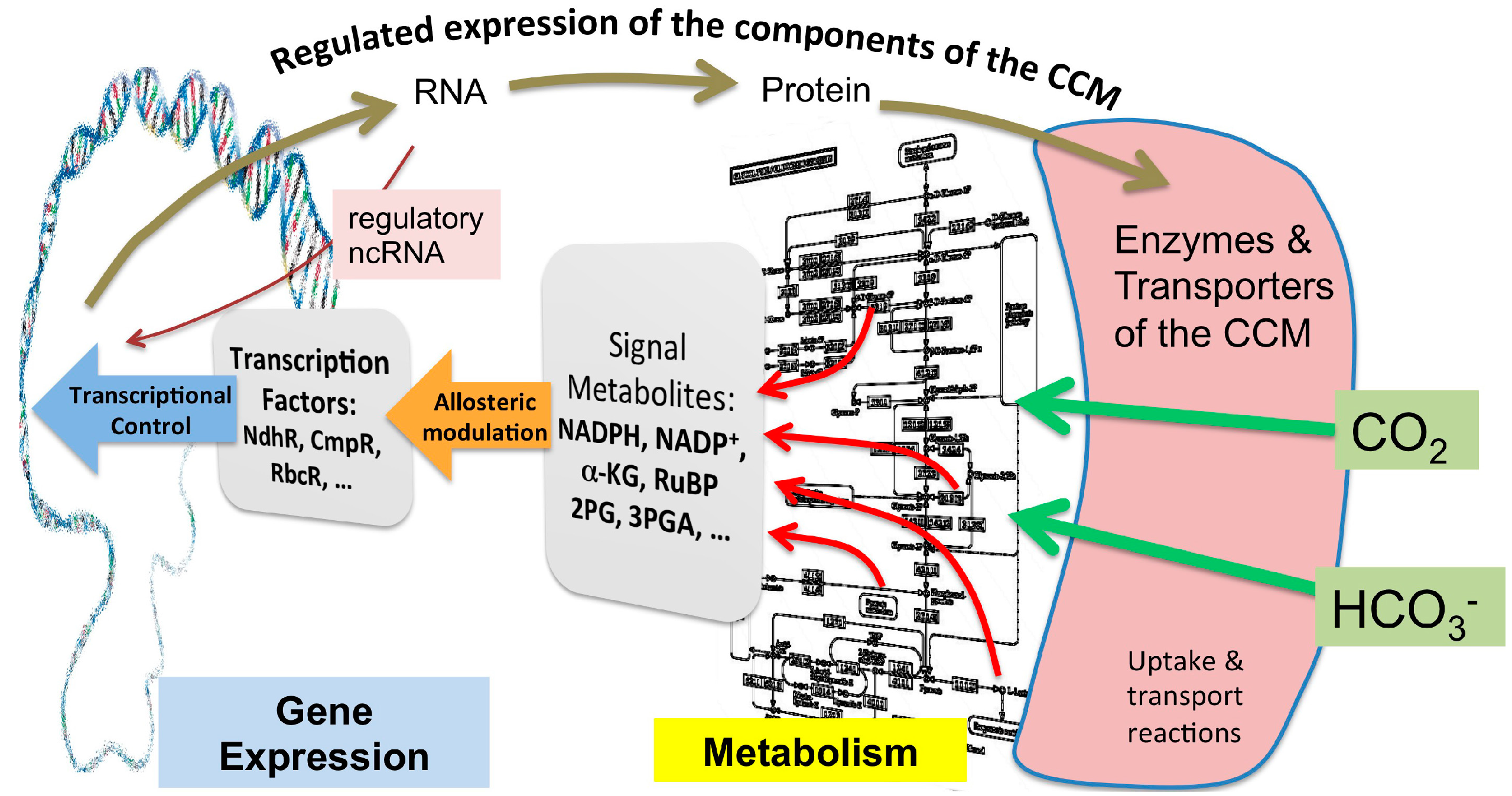{kind=link}
| Gene Name | Synechocystis ORF | Function | Co-regulatory Metabolites | Reference |
|---|---|---|---|---|
| ndhR (ccmR) | sll1594 | Repressor high affinity Ci uptake (genes for CupA, SbtA, Na+-NDH-1) | α-KG, NADP+ | [31,32,89,97] |
| cmpR | sll0030 | Activator of ABC-type bicarbonate transporter (cmp operon and psbA genes) | RuBP, 2PG | [32,35,89,97] |
| ycf30, rbcR | sll0998 | Activation of CBB genes | NADPH, 3PGA, RuBP | [89,97,98] |
| ntcB | slr0395 | Activation of nitrate assimilation genes | nitrite | [99] |
3.2. Post-Transcriptional Regulation of the CCM

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fukuzawa, H.; Ogawa, T.; Kaplan, A. The uptake of CO2 by cyanobacteria and microalgae. In Photosynthesis: Plastid Biology, Energy Conversion and Carbon Assimilation; Eaton Rye, J.J., Tripathy, B.C., Sharkey, T.D., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 34, pp. 625–650. [Google Scholar]
- Kaplan, A.; Reinhold, L. The CO2 concentrating mechanisms in photosynthetic microorganisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 539–570. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Kaplan, A. Inorganic carbon acquisition systems in cyanobacteria. Photosynth. Res. 2003, 77, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 concentrating mechanisms in algae: Mechanisms, environmental modulation, and evolution. Ann. Rev. Plant Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef]
- Price, G.D.; Sültemeyer, D.; Klughammer, B.; Ludwig, M.; Badger, M.R. The functioning of the CO2 concentrating mechanism in several cyanobacterial strains: A review of general physiological characteristics, genes, proteins and recent advances. Can. J. Bot. 1998, 76, 973–1002. [Google Scholar]
- Kupriyanova, E.V.; Sinetova, M.A.; Cho, S.M.; Park, Y.I.; Los, D.; Pronina, N.A. CO2-concentrating mechanism in cyanobacterial photosynthesis: Organization, physiological role, and evolutionary origin. Photosynth. Res. 2013, 117, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Rae, B.D.; Long, B.M.; Badger, M.R.; Price, G.D. Functions, compositions, and evolution of the two types of carboxysomes: Polyhedral microcompartments that facilitate CO2 fixation in cyanobacteria and some proteobacteria. Microbiol. Mol. Biol. Rev. 2013, 77, 357–379. [Google Scholar] [CrossRef] [PubMed]
- Mangan, N.M.; Brenner, M.P. Systems analysis of the CO2 concentrating mechanism in cyanobacteria. eLife 2014, 3. [Google Scholar] [CrossRef]
- Badger, M.R.; Kaplan, A.; Berry, J.A. The internal inorganic carbon pool of Chlamydomonas reinhardtii: Evidence for a CO2 concentrating mechanism. Plant Physiol. 1980, 66, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, A.; Badger, M.R.; Berry, J.A. Photosynthesis and intracellular inorganic carbon pool in the blue-green algae Anabaena variabilis: Response to external CO2 concentration. Planta 1980, 149, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, R.; Reinhold, L.; Kaplan, A. Low activation state of ribulose 1,5-bisphosphate carboxylase/oxygenase in carboxysome-defective Synechococcus mutants. Plant Physiol. 1995, 108, 183–190. [Google Scholar] [PubMed]
- Eisenhut, M.; Ruth, W.; Haimovich, M.; Bauwe, H.; Kaplan, A.; Hagemann, M. The photorespiratory glycolate metabolism is essential for cyanobacteria and might have been conveyed endosymbiontically to plants. Proc. Natl. Acad. Sci. USA 2008, 105, 17199–17204. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, M.; Fernie, A.R.; Espie, G.S.; Kern, R.; Eisenhut, M.; Reumann, S.; Bauwe, H.; Weber, A.P.M. Evolution of the biochemistry of the photorespiratory C2 cycle. Plant Biol. 2013, 15, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Tchernov, D.; Hassidim, M.; Luz, B.; Sukenik, A.; Reinhold, L.; Kaplan, A. Sustained net CO2 evolution during photosynthesis by marine microorganisms. Curr. Biol. 1997, 7, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Bernat, G.; Singh, A.; Mi, H.; Rogner, M.; Pakrasi, H.B.; Ogawa, T. Properties of mutants of Synechocystis sp. strain PCC 6803 lacking inorganic carbon sequestration systems. Plant Cell Physiol. 2008, 49, 1672–1677. [Google Scholar] [CrossRef] [PubMed]
- Tchernov, D.; Helman, Y.; Keren, N.; Luz, B.; Ohad, I.; Reinhold, L.; Ogawa, T.; Kaplan, A. Passive entry of CO2 and its energy-dependent intracellular conversion to HCO3− in cyanobacteria are driven by a photosystem I-generated ∆µH+. J. Biol. Chem. 2001, 276, 23450–23455. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Matsumoto, T.; Gena, P.; Liu, C.; Pellegrini-Calace, M.; Zhong, S.; Sun, X.; Zhu, Y.; Katsuhara, M.; Iwasaki, I.; et al. Water and CO2 permeability of SsAqpZ, the cyanobacterium Synechococcus sp PCC7942 aquaporin. Biol. Cell 2013, 105, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Kaldenhoff, R.; Kai, L.; Uehlein, N. Aquaporins and membrane diffusion of CO2 in living organisms. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 1592–1595. [Google Scholar] [CrossRef]
- Schwarz, D.; Schubert, H.; Georg, J.; Hess, W.R.; Hagemann, M. The gene sml0013 of Synechocystis species strain PCC 6803 encodes for a novel subunit of the NAD(P)H oxidoreductase or complex I that is ubiquitously distributed among cyanobacteria. Plant Physiol. 2013, 163, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gao, F.; Zhao, J.; Ogawa, T.; Wang, Q.; Ma, W. NdhP Is an exclusive subunit of large complex of NADPH dehydrogenase essential to stabilize the complex in Synechocystis sp strain PCC 6803. J. Biol. Chem. 2014, 289, 18770–18781. [Google Scholar] [CrossRef] [PubMed]
- Battchikova, N.; Eisenhut, M.; Aro, E.M. Cyanobacterial NDH-1 complexes: Novel insights and remaining puzzles. Biochim. Biophys. Acta Bioenerg. 2011, 1807, 935–944. [Google Scholar] [CrossRef]
- Ogawa, T.; Harada, T.; Ozaki, H.; Sonoike, K. Disruption of the ndhF1 gene affects Chl fluorescence through state transition in the cyanobacterium Synechocystis sp. PCC 6803, resulting in apparent high efficiency of photosynthesis. Plant Cell Physiol. 2013, 54, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Ohkawa, H.; Kaneko, T.; Fukuzawa, H.; Tabata, S.; Kaplan, A.; Ogawa, T. Distinct constitutive and low-CO2-induced CO2 uptake systems in cyanobacteria: Genes involved and their phylogenetic relationship with homologous genes in other organisms. Proc. Natl. Acad. Sci. USA 2001, 98, 11789–11794. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Ogawa, T. Oxygenic photosynthesis-specific subunits of cyanobacterial NADPH dehydrogenases. IUBMB Life 2015. [Google Scholar] [CrossRef]
- Burnap, R.L.; Nambudiri, R.; Holland, S. Regulation of the carbon-concentrating mechanism in the cyanobacterium Synechocystis sp. PCC6803 in response to changing light intensity and inorganic carbon availability. Photosynth. Res. 2013, 118, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Price, G.D. Inorganic carbon transporters of the cyanobacterial CO2 concentrating mechanism. Photosynth. Res. 2011, 109, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Mi, H. Cyanobacterial NADPH dehydrogenase complexes. Photosynth. Res. 2007, 93, 69–77. [Google Scholar] [CrossRef]
- Zhang, P.; Battchikova, N.; Jansen, T.; Appel, J.; Ogawa, T.; Aro, E.M. Expression and functional roles of the two distinct NDH-1 complexes and the carbon acquisition complex NdhD3/NdhF3/CupA/Sll1735 in Synechocystis sp PCC 6803. Plant Cell 2004, 16, 3326–3340. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Rivero, A.; Leganes, F.; Fernandez-Valiente, E.; Calle, P.; Fernandez-Pinas, F. mrpA, a gene with roles in resistance to Na+ and adaptation to alkaline pH in the cyanobacterium Anabaena sp. PCC7120. Microbiology 2005, 151, 1671–1682. [Google Scholar] [CrossRef] [PubMed]
- Morino, M.; Natsui, S.; Ono, T.; Swartz, T.H.; Krulwich, T.A.; Ito, M. Single site mutations in the hetero-oligomeric Mrp Antiporter from alkaliphilic Bacillus pseudofirmus OF4 that affect Na+/H+ antiport activity, sodium exclusion, individual Mrp protein levels, or Mrp complex formation. J. Biol. Chem. 2010, 285, 30942–30950. [Google Scholar] [CrossRef] [PubMed]
- Daley, S.M.; Kappell, A.D.; Carrick, M.J.; Burnap, R.L. Regulation of the cyanobacterial CO2-concentrating mechanism involves internal sensing of NADP+ and alpha-ketogutarate levels by transcription factor CcmR. PLoS One 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Postier, B.L.; Burnap, R.L. Alterations in global patterns of gene expression in Synechocystis sp. PCC 6803 in response to inorganic carbon limitation and the inactivation of ndhR, a LysR family regulator. J. Biol. Chem. 2004, 279, 5739–5751. [Google Scholar] [CrossRef] [PubMed]
- Battchikova, N.; Wei, L.; Du, L.; Bersanini, L.; Aro, E.-M.; Ma, W. Identification of novel Ssl0352 protein (NdhS), essential for efficient operation of cyclic electron transport around photosystem I, in NADPH: Plastoquinone oxidoreductase (NDH-1) complexes of Synechocystis sp. PCC 6803. J. Biol. Chem. 2012, 287. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.D.; Fewer, D.P.; Talla, E.; Calteau, A.; Cai, F.; Tandeau de Marsac, N.; Rippka, R.; et al. Improving the coverage of the cyanobacterial phylum using diversity-driven genome sequencing. Proc. Natl. Acad. Sci. USA 2012, 110, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Omata, T.; Price, D.G.; Badger, M.R.; Okamura, M.; Gohta, S.; Ogawa, T. Identification of an ATP-binding cassette transporter involved in bicarbonate uptake in the cyanobacterium Synechococcus sp. strain PCC 7942. Proc. Natl. Acad. Sci. USA 1999, 96, 13571–13576. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Katoh, H.; Sonoda, M.; Ohkawa, H.; Shimoyama, M.; Fukuzawa, H.; Kaplan, A.; Ogawa, T. Genes essential to sodium-dependent bicarbonate transport in cyanobacteria: Function and phylogenetic analysis. J. Biol. Chem. 2002, 277, 18658–18664. [Google Scholar] [CrossRef] [PubMed]
- Price, G.D.; Woodger, F.J.; Badger, M.R.; Howitt, S.M.; Tucker, L. Identification of a SulP-type bicarbonate transporter in marine cyanobacteria. Proc. Nat. Acad. Sci. USA 2004, 101, 18228–18233. [Google Scholar] [CrossRef] [PubMed]
- Woodger, F.J.; Bryant, D.A.; Price, G.D. Transcriptional regulation of the CO2-concentrating mechanism in a euryhaline, coastal marine cyanobacterium, Synechococcus sp. Strain PCC 7002: Role of NdhR/CcmR. J. Bacteriol. 2007, 189, 3335–3347. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.; Pisareva, T.; Norling, B. Proteomic studies of the thylakoid membrane of Synechocystis sp. PCC 6803. Proteomics 2005, 5, 4905–4916. [Google Scholar] [CrossRef] [PubMed]
- Sandrini, G.; Matthijs, H.C.P.; Verspagen, J.M.H.; Muyzer, G.; Huisman, J. Genetic diversity of inorganic carbon uptake systems causes variation in CO2 response of the cyanobacterium Microcystis. ISME J. 2014, 8, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Gaudana, S.; Zarzycki, J.; Moparthi, V.; Kerfeld, C. Bioinformatic analysis of the distribution of inorganic carbon transporters and prospective targets for bioengineering to increase Ci uptake by cyanobacteria. Photosynth. Res. 2014. [Google Scholar] [CrossRef]
- So, A.K.C.; Cot, S.S.W.; Espie, G.S. Characterization of the C-terminal extension of carboxysomal carbonic anhydrase from Synechocystis sp PCC6803. Funct. Plant Biol. 2002, 29, 183–194. [Google Scholar] [CrossRef]
- Kinney, J.N.; Axen, S.D.; Kerfeld, C.A. Comparative analysis of carboxysome shell proteins. Photosynth. Res. 2011, 109, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Hess, W.R.; Rocap, G.; Ting, C.S.; Larimer, F.; Stilwagen, S.; Lamerdin, J.; Chisholm, S.W. The photosynthetic apparatus of Prochlorococcus: Insights through comparative genomics. Photosynth. Res. 2001, 70, 53–71. [Google Scholar] [CrossRef] [PubMed]
- Badger, M.R.; Hanson, D.; Price, G.D. Evolution and diversity of CO2 concentrating mechanisms in cyanobacteria. Funct. Plant Biol. 2002, 29, 161–173. [Google Scholar] [CrossRef]
- Yeates, T.O.; Kerfeld, C.A.; Heinhorst, S.; Cannon, G.C.; Shively, J.M. Protein-based organelles in bacteria: Carboxysomes and related microcompartments. Nat. Rev. Microbiol. 2008, 6, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Fukuzawa, H.; Suzuki, E.; Komukai, Y.; Miyachi, S. A gene homologous to chloroplast carbonic anhydrase (icfA) is essential to photosynthetic carbon dioxide fixation by Synechococcus PCC7942. Proc. Natl. Acad. Sci. USA 1992, 89, 4437–4441. [Google Scholar] [CrossRef] [PubMed]
- Pena, K.L.; Castel, S.E.; de Araujo, C.; Espie, G.S.; Kimber, M.S. Structural basis of the oxidative activation of the carboxysomal gamma-carbonic anhydrase, CcmM. Proc. Natl. Acad. Sci. USA 2010, 107, 2455–2460. [Google Scholar] [CrossRef]
- Kupriyanova, E.V.; Sinetova, M.A.; Markelova, A.G.; Allakhverdiev, S.I.; Los, D.A.; Pronina, N.A. Extracellular beta-class carbonic anhydrase of the alkaliphilic cyanobacterium Microcoleus chthonoplastes. J. Photochem. Photobiol. B Biol. 2011, 103, 78–86. [Google Scholar] [CrossRef]
- Cameron, J.C.; Wilson, S.C.; Bernstein, S.L.; Kerfeld, C.A. Biogenesis of a bacterial organelle: The carboxysome assembly pathway. Cell 2013, 155, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Espie, G.S.; Kimber, M.S. Carboxysomes: Cyanobacterial RubisCO comes in small packages. Photosynth. Res. 2011, 109, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Cot, S.S.; So, A.K.; Espie, G.S. A multiprotein bicarbonate dehydration complex essential to carboxysome function in cyanobacteria. J. Bacteriol. 2008, 190, 936–945. [Google Scholar] [CrossRef] [PubMed]
- Benschop, J.J.; Badger, M.R.; Price, D.G. Characterisation of CO2 and HCO3− uptake in the cyanobacterium Synechocystis sp. PCC6803. Photosynth. Res. 2003, 77, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Tabita, F.R. The biochemistry and molecular regulation of carbon dioxide metabolism in cyanobacteria. In The Molecular Biology of Cyanobacteria; Bryant, D.A., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; pp. 437–467. [Google Scholar]
- Weathers, P.J.; Allen, M.M. Variations in short term products of inorganic carbon fixation in exponential and stationary phase cultures of Aphanocapsa 6308. Arch. Microbiol. 1978, 116, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Huege, J.; Goetze, J.; Schwarz, D.; Bauwe, H.; Hagemann, M.; Kopka, J. Modulation of the major paths of carbon in photorespiratory mutants of Synechocystis. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Young, J.D.; Shastri, A.A.; Stephanopoulos, G.; Morgan, J.A. Mapping photoautotrophic metabolism with isotopically nonstationary 13C flux analysis. Metab. Eng. 2011, 13, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Bauwe, H.; Hagemann, M.; Fernie, A.R. Photorespiration: Players, partners and origin. Trends Plant Sci. 2010, 15, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Eisenhut, M.; Huege, J.; Schwarz, D.; Bauwe, H.; Kopka, J.; Hagemann, M. Metabolome phenotyping of inorganic carbon limitation in cells of the wild type and photorespiratory mutants of the cyanobacterium Synechocystis sp strain PCC 6803. Plant Physiol. 2008, 148, 2109–2120. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, R.; Ortleb, S.; Sainis, J.K.; Melzer, M. Immunoelectron microscopy for locating Calvin cycle enzymes in the thylakoids of Synechocystis 6803. Mol. Plant 2009, 2, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Hackenberg, C.; Huege, J.; Engelhardt, A.; Wittink, F.; Laue, M.; Matthijs, H.C.; Kopka, J.; Bauwe, H.; Hagemann, M. Low-carbon acclimation in carboxysome-less and photorespiratory mutants of the cyanobacterium Synechocystis sp. strain PCC 6803. Microbiology 2012, 158, 398–413. [Google Scholar] [CrossRef] [PubMed]
- Jablonsky, J.; Schwarz, D.; Hagemann, M. Multi-level kinetic model explaining diverse roles of isozymes in prokaryotes. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.; Orf, I.; Kopka, J.; Hagemann, M. Recent applications of metabolomics toward cyanobacteria. Metabolites 2013, 3, 72–100. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, T.D.; Burns, R.A.; Campbell, D.A. Carbon status constrains light acclimation in the cyanobacterium Synechococcus elongatus. Plant Physiol. 2004, 136, 3301–3312. [Google Scholar] [CrossRef] [PubMed]
- Pelroy, R.A.; Bassham, J.A. Kinetics of glucose incorporation by Aphanocapsa 6714. J. Bacteriol. 1973, 115, 943–948. [Google Scholar] [PubMed]
- Shimakawa, G.; Hasunuma, T.; Kondo, A.; Matsuda, M.; Makino, A.; Miyake, C. Respiration accumulates Calvin cycle intermediates for the rapid start of photosynthesis in Synechocystis sp. PCC 6803. Biosci. Biotechnol. Biochem. 2014, 78, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Carrieri, D.; Paddock, T.; Maness, P.-C.; Seibert, M.; Yu, J. Photo-catalytic conversion of carbon dioxide to organic acids by a recombinant cyanobacterium incapable of glycogen storage. Energy Environ. Sci. 2012, 5, 9457–9461. [Google Scholar] [CrossRef]
- Xu, Y.; Guerra, L.T.; Li, Z.; Ludwig, M.; Dismukes, G.C.; Bryant, D.A. Altered carbohydrate metabolism in glycogen synthase mutants of Synechococcus sp. strain PCC 7002: Cell factories for soluble sugars. Metab. Eng. 2013, 16, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Bohringer, J.; Fischer, D.; Mosler, G.; Hengge-Aronis, R. UDP-glucose is a potential intracellular signal molecule in the control of expression of sigma S and sigma S-dependent genes in Escherichia coli. J. Bacteriol. 1995, 177, 413–422. [Google Scholar] [PubMed]
- Price, G.D.; Maeda, S.; Omata, T.; Badger, M.R. Modes of active inorganic carbon uptake in the cyanobacterium, Synechococcus sp PCC7942. Funct. Plant Biol. 2002, 29, 131–149. [Google Scholar] [CrossRef]
- Schwarz, D.; Nodop, A.; Huge, J.; Purfurst, S.; Forchhammer, K.; Michel, K.P.; Bauwe, H.; Kopka, J.; Hagemann, M. Metabolic and transcriptomic phenotyping of inorganic carbon acclimation in the cyanobacterium Synechococcus elongatus PCC 7942. Plant Physiol. 2011, 155, 1640–1655. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.; Orf, I.; Kopka, J.; Hagemann, M. Effects of inorganic carbon limitation on the metabolome of the Synechocystis sp. PCC 6803 mutant defective in glnB encoding the central regulator PII of cyanobacterial C/N acclimation. Metabolites 2014, 4, 232–247. [Google Scholar] [CrossRef] [PubMed]
- Muro-Pastor, M.I.; Reyes, J.C.; Florencio, F.J. Cyanobacteria perceive nitrogen status by sensing intracellular 2-oxoglutarate levels. J. Biol. Chem. 2001, 276, 38320–38328. [Google Scholar] [PubMed]
- Gründel, M.; Scheunemann, R.; Lockau, W.; Zilliges, Y. Impaired glycogen synthesis causes metabolic overflow reactions and affects stress responses in the cyanobacterium Synechocystis sp. PCC 6803. Microbiology 2012, 158, 3032–3043. [Google Scholar] [CrossRef] [PubMed]
- Eisenhut, M.; von Wobeser, E.A.; Jonas, L.; Schubert, H.; Ibelings, B.W.; Bauwe, H.; Matthijs, H.C.; Hagemann, M. Long-term response toward inorganic carbon limitation in wild type and glycolate turnover mutants of the cyanobacterium Synechocystis sp. strain PCC 6803. Plant Physiol. 2007, 144, 1946–1959. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Troya, S.; Lopez-Maury, L.; Sanchez-Riego, A.M.; Roldan, M.; Florencio, F.J. Redox regulation of glycogen biosynthesis in the cyanobacterium Synechocystis sp. PCC 6803: Analysis of the AGP and glycogen synthases. Mol. Plant 2014, 7, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, M.; Florencio, F.J. Thioredoxin-linked processes in cyanobacteria are as numerous as in chloroplasts, but targets are different. Proc. Natl. Acad. Sci. USA 2003, 100, 16107–16112. [Google Scholar] [CrossRef] [PubMed]
- Mikkat, S.; Fulda, S.; Hagemann, M. A 2D gel electrophoresis-based snapshot of the phosphoproteome in the cyanobacterium Synechocystis sp. strain PCC 6803. Microbiology 2014, 160, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Rocha, A.G.; Mehlmer, N.; Stael, S.; Mair, A.; Parvin, N.; Chigri, F.; Teige, M.; Vothknecht, U.C. Phosphorylation of Arabidopsis transketolase at Ser428 provides a potential paradigm for the metabolic control of chloroplast carbon metabolism. Biochem. J. 2014, 458, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Zilliges, Y.; Kehr, J.C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Borner, T.; Dittmann, E. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of microcystis under oxidative stress conditions. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Meissner, S.; Steinhauser, D.; Dittmann, E. Metabolomic analysis indicates a pivotal role of the hepatotoxin microcystin in high light adaptation of Microcystis. Environ. Microbiol. 2014. [Google Scholar] [CrossRef]
- Allahverdiyeva, Y.; Ermakova, M.; Eisenhut, M.; Zhang, P.; Richaud, P.; Hagemann, M.; Cournac, L.; Aro, E.M. Interplay between flavodiiron proteins and photorespiration in Synechocystis sp. PCC 6803. J. Biol. Chem. 2011, 286, 24007–24014. [Google Scholar] [CrossRef] [PubMed]
- Hackenberg, C.; Engelhardt, A.; Matthijs, H.C.; Wittink, F.; Bauwe, H.; Kaplan, A.; Hagemann, M. Photorespiratory 2-phosphoglycolate metabolism and photoreduction of O2 cooperate in high-light acclimation of Synechocystis sp. strain PCC 6803. Planta 2009, 230, 625–637. [Google Scholar] [CrossRef] [PubMed]
- McGinn, P.J.; Price, G.D.; Maleszka, R.; Badger, M.R. Inorganic carbon limitation and light control the expression of transcripts related to the CO2-concentrating mechanism in the cyanobacterium Synechocystis sp. strain PCC6803. Plant Physiol. 2003, 132, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Woodger, F.J.; Badger, M.R.; Price, G.D. Inorganic carbon limitation induces transcripts encoding components of the CO2-concentrating mechanism in Synechococcus sp. PCC7942 through a redox-independent pathway. Plant Physiol. 2003, 133, 2069–2080. [Google Scholar] [CrossRef] [PubMed]
- McGinn, P.J.; Price, G.D.; Badger, M.R. High light enhances the expression of low-CO2-inducible transcripts involved in the CO2-concentrating mechanism in Synechocystis sp. PCC6803. Plant Cell Environ. 2004, 27, 615–626. [Google Scholar] [CrossRef]
- Woodger, F.J.; Badger, M.; Price, G.D. Regulation of cyanobacterial CO2-concentrating mechanisms through transcriptional induction of high-affinity Ci-transport systems. Can. J. Bot. 2005, 83, 698–710. [Google Scholar] [CrossRef]
- Woodger, F.J.; Badger, M.R.; Price, G.D. Sensing of inorganic carbon limitation in Synechococcus PCC7942 is correlated with the size of the internal inorganic carbon pool and involves oxygen. Plant Physiol. 2005, 139, 1959–1969. [Google Scholar] [CrossRef] [PubMed]
- Omata, T.; Gohta, S.; Takahashi, Y.; Harano, Y.; Maeda, S. Involvement of a CbbR homolog in low CO2-induced activation of the bicarbonate transporter operon in cyanobacteria. J. Bacteriol. 2001, 183, 1891–1898. [Google Scholar] [CrossRef] [PubMed]
- Momany, C.; Neidle, E.L. Defying stereotypes: The elusive search for a universal model of LysR-type regulation. Mol. Microbiol. 2012, 83, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, S.E.; Oyston, P.C. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 2008, 154, 3609–3623. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Takahashi, Y.; Yamaguchi, O.; Suzuki, H.; Maeda, S.I.; Omata, T. Mechanism of low CO2-induced activation of the cmp bicarbonate transporter operon by a LysR family protein in the cyanobacterium Synechococcus elongatus strain PCC 7942. Mol. Microbiol. 2008, 68, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Haimovich-Dayan, M.; Lieman-Hurwitz, J.; Orf, I.; Hagemann, M.; Kaplan, A. Does 2-phosphoglycolate serve as an internal signal molecule of inorganic carbon deprivation in the cyanobacterium Synechocystis sp. PCC 6803? Environ. Microbiol. 2014. [Google Scholar] [CrossRef]
- Gibson, J.L.; Tabita, F.R. The molecular regulation of the reductive pentose phosphate pathway in Proteobacteria and Cyanobacteria. Arch. Microbiol. 1996, 166, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Joshi, G.S.; Zianni, M.; Bobst, C.E.; Tabita, F.R. Regulatory twist and synergistic role of metabolic coinducer- and response regulator-mediated CbbR-cbb(I) interactions in Rhodopseudomonas palustris CGA010. J. Bacteriol. 2013, 195, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Yamaguchi, O.; Omata, T. Roles of CmpR, a LysR family transcriptional regulator, in acclimation of the cyanobacterium Synechococcus sp. strain PCC 7942 to low-CO and high-light conditions. Mol. Microbiol. 2004, 52, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Figge, R.M.; Cassier-Chauvat, C.; Chauvat, F.; Cerff, R. Characterization and analysis of an NAD(P)H dehydrogenase transcriptional regulator critical for the survival of cyanobacteria facing inorganic carbon starvation and osmotic stress. Mol. Microbiol. 2001, 39, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Minoda, A.; Weber, A.P.; Tanaka, K.; Miyagishima, S.Y. Nucleus-independent control of the RubisCO operon by the plastid-encoded transcription factor Ycf30 in the red alga Cyanidioschyzon merolae. Plant Physiol. 2010, 154, 1532–1540. [Google Scholar] [CrossRef] [PubMed]
- Aichi, M.; Takatani, N.; Omata, T. Role of NtcB in activation of nitrate assimilation genes in the cyanobacterium Synechocystis sp. strain PCC 6803. J. Bacteriol. 2001, 183, 5840–5847. [Google Scholar] [CrossRef] [PubMed]
- Lieman-Hurwitz, J.; Haimovich, M.; Shalev-Malul, G.; Ishii, A.; Hihara, Y.; Gaathon, A.; Lebendiker, M.; Kaplan, A. A cyanobacterial AbrB-like protein affects the apparent photosynthetic affinity for CO2 by modulating low-CO2-induced gene expression. Environ. Microbiol. 2009, 11, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Marcus, Y.; Harel, E.; Kaplan, A. Adaptation of the cyanobacterium Anabaena variabilis to low CO2 concentration in their environment. Plant Physiol. 1983, 71, 208–210. [Google Scholar] [CrossRef] [PubMed]
- Muro-Pastor, M.I.; Reyes, J.C.; Florencio, F.J. Ammonium assimilation in cyanobacteria. Photosynth. Res. 2005, 83, 135–150. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Dominguez, M.; Reyes, J.C.; Florencio, F.J. NtcA represses transcription of gifA and gifB, genes that encode inhibitors of glutamine synthetase type I from Synechocystis sp PCC 6803. Mol. Microbiol. 2000, 35, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Galmozzi, C.V.; Fernandez-Avila, M.J.; Reyes, J.C.; Florencio, F.J.; Muro-Pastor, M.I. The ammonium-inactivated cyanobacterial glutamine synthetase I is reactivated in vivo by a mechanism involving proteolytic removal of its inactivating factors. Mol. Microbiol. 2007, 65, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Tichi, M.A.; Tabita, F.R. Metabolic Signals That Lead to Control of CBB Gene Expression in Rhodobacter capsulatus. J. Bacteriol. 2002, 184, 1905–1915. [Google Scholar] [CrossRef] [PubMed]
- Maier, U.-G.; Fraunholz, M.; Zauner, S.; Penny, S.; Douglas, S. A Nucleomorph-Encoded CbbX and the Phylogeny of RuBisCO Regulators. Mol. Biol. Evol. 2000, 17, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Hihara, Y.; Kamei, A.; Kanehisa, M.; Kaplan, A.; Ikeuchi, M. DNA microarray analysis of cyanobacterial gene expression during acclimation to high light. Plant Cell 2001, 13, 793–806. [Google Scholar] [CrossRef] [PubMed]
- Rippka, R.; Deruelles, J.; Waterbury, J.-B.; Herdman, M.; Stanier, R.-Y. Genetic assignments, strain histories and properties of pure cultures of cyanobateria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Stal, L.J.; Moezelaar, R. Fermentation in cyanobacteria. FEMS Microbiol. Rev. 1997, 21, 179–211. [Google Scholar] [CrossRef]
- Azuma, M.; Osanai, T.; Hirai, M.Y.; Tanaka, K. A response regulator Rre37 and an RNA polymerase sigma factor SigE represent two parallel pathways to activate sugar catabolism in a cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 2011, 52, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Osanai, T.; Oikawa, A.; Azuma, M.; Tanaka, K.; Saito, K.; Hirai, M.Y.; Ikeuchi, M. Genetic engineering of group 2 sigma factor SigE widely activates expressions of sugar catabolic genes in Synechocystis species PCC 6803. J. Biol. Chem. 2011, 286, 30962–30971. [Google Scholar] [CrossRef] [PubMed]
- Bloye, S.A.; Silman, N.J.; Mann, N.H.; Carr, N.G. Bicarbonate concentration by Synechocystis PCC6803: Modulation of protein phosphorylation and inorganic carbon transport by glucose. Plant Physiol. 1992, 99, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Haimovich-Dayan, M.; Kahlon, S.; Hihara, Y.; Hagemann, M.; Ogawa, T.; Ohad, I.; Kaplan, A. Cross-talk between photomixotrophic growth and CO2-concentrating mechanism in Synechocystis sp strain PCC 6803. Environ. Microbiol. 2011, 13, 1767–1777. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, A.; Hagemann, M.; Bauwe, H.; Kahlon, S.; Ogawa, T. Carbon acquisition by cyanobacteria: Mechanisms, comparative genomics and evolution. In The Cyanobacteria: Molecular Biology, Genomics and Evolution; Herrero, A., Flores, E., Eds.; Caister Academic Press: Norwich, UK, 2008; pp. 305–323. [Google Scholar]
- Chen, Y.; Cann, M.J.; Litvin, T.N.; Iourgenko, V.; Sinclair, M.L.; Levin, L.R.; Buck, J. Soluble adenylyl cyclase as an evolutionarily conserved bicarbonate sensor. Science 2000, 289, 625–628. [Google Scholar] [CrossRef] [PubMed]
- Cann, M.J.; Hammer, A.; Zhou, J.; Kanacher, T. A defined subset of adenylyl cyclases is regulated by bicarbonate ion. J. Biol. Chem. 2003, 278, 35033–35038. [Google Scholar] [CrossRef]
- Hammer, A.; Hodgson, D.R.; Cann, M.J. Regulation of prokaryotic adenylyl cyclases by CO2. Biochem. J. 2006, 396, 215–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terauchi, K.; Ohmori, M. An adenylate cyclase, Cya1, regulates cell motility in the cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 1999, 40, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Georg, J.; Hess, W.R. cis-antisense RNA, another level of gene regulation in bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Mitschke, J.; Georg, J.; Scholz, I.; Sharma, C.M.; Dienst, D.; Bantscheff, J.; Voss, B.; Steglich, C.; Wilde, A.; Vogel, J.; et al. An experimentally anchored map of transcriptional start sites in the model cyanobacterium Synechocystis sp. PCC6803. Proc. Natl. Acad. Sci. USA 2011, 108, 2124–2129. [Google Scholar] [CrossRef] [PubMed]
- Mitschke, J.; Vioque, A.; Haas, F.; Hess, W.R.; Muro-Pastor, A.M. Dynamics of transcriptional start site selection during nitrogen stress-induced cell differentiation in Anabaena sp. PCC7120. Proc. Natl. Acad. Sci. USA 2011, 108, 20130–20135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopf, M.; Klahn, S.; Scholz, I.; Matthiessen, J.K.; Hess, W.R.; Voss, B. Comparative analysis of the primary transcriptome of Synechocystis sp. PCC 6803. DNA Res. 2014, 21, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Eisenhut, M.; Georg, J.; Klahn, S.; Sakurai, I.; Mustila, H.; Zhang, P.; Hess, W.R.; Aro, E.M. The antisense RNA As1_flv4 in the cyanobacterium Synechocystis sp. PCC 6803 prevents premature expression of the flv4–2 operon upon shift in inorganic carbon supply. J. Biol. Chem. 2012, 287, 33153–33162. [Google Scholar] [CrossRef] [PubMed]
- Georg, J.; Dienst, D.; Schurgers, N.; Wallner, T.; Kopp, D.; Stazic, D.; Kuchmina, E.; Klahn, S.; Lokstein, H.; Hess, W.R.; et al. The small regulatory RNA SyR1/PsrR1 controls photosynthetic functions in cyanobacteria. Plant Cell 2014, 26, 3661–3679. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burnap, R.L.; Hagemann, M.; Kaplan, A. Regulation of CO2 Concentrating Mechanism in Cyanobacteria. Life 2015, 5, 348-371. https://doi.org/10.3390/life5010348
Burnap RL, Hagemann M, Kaplan A. Regulation of CO2 Concentrating Mechanism in Cyanobacteria. Life. 2015; 5(1):348-371. https://doi.org/10.3390/life5010348
Chicago/Turabian StyleBurnap, Robert L., Martin Hagemann, and Aaron Kaplan. 2015. "Regulation of CO2 Concentrating Mechanism in Cyanobacteria" Life 5, no. 1: 348-371. https://doi.org/10.3390/life5010348