
Successful Production of Offspring Derived from Phospholipase C Zeta-Deficient Sperm by Additional Artificial Activation
 , and
, and 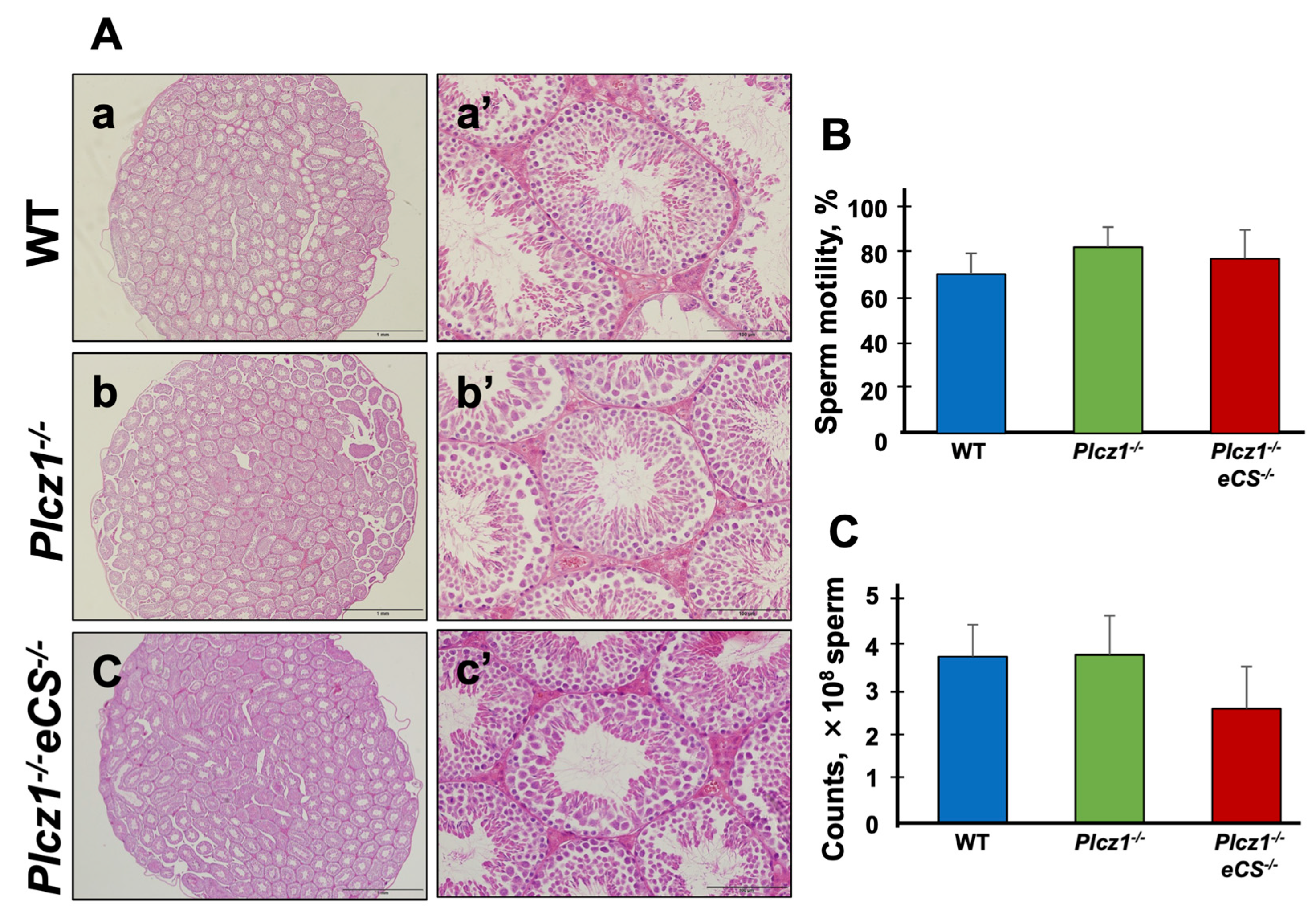{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Histology
2.4. Sperm Cryopreservation and Sperm Motility Analysis
2.5. Oocyte Preparation
2.6. Preparation of Mouse PLCζ mRNA and Its Microinjection
2.7. Production of Fertilized Embryos by Intracytoplasmic Sperm Injection
2.8. In Vitro Culture and Embryo Transfer
2.9. Immunostaining
2.10. Statistical Analysis
3. Results
3.1. The Phenotype of Testes and Sperm Derived from Plcz1−/− or Plcz1−/−eCS−/− Male Mice
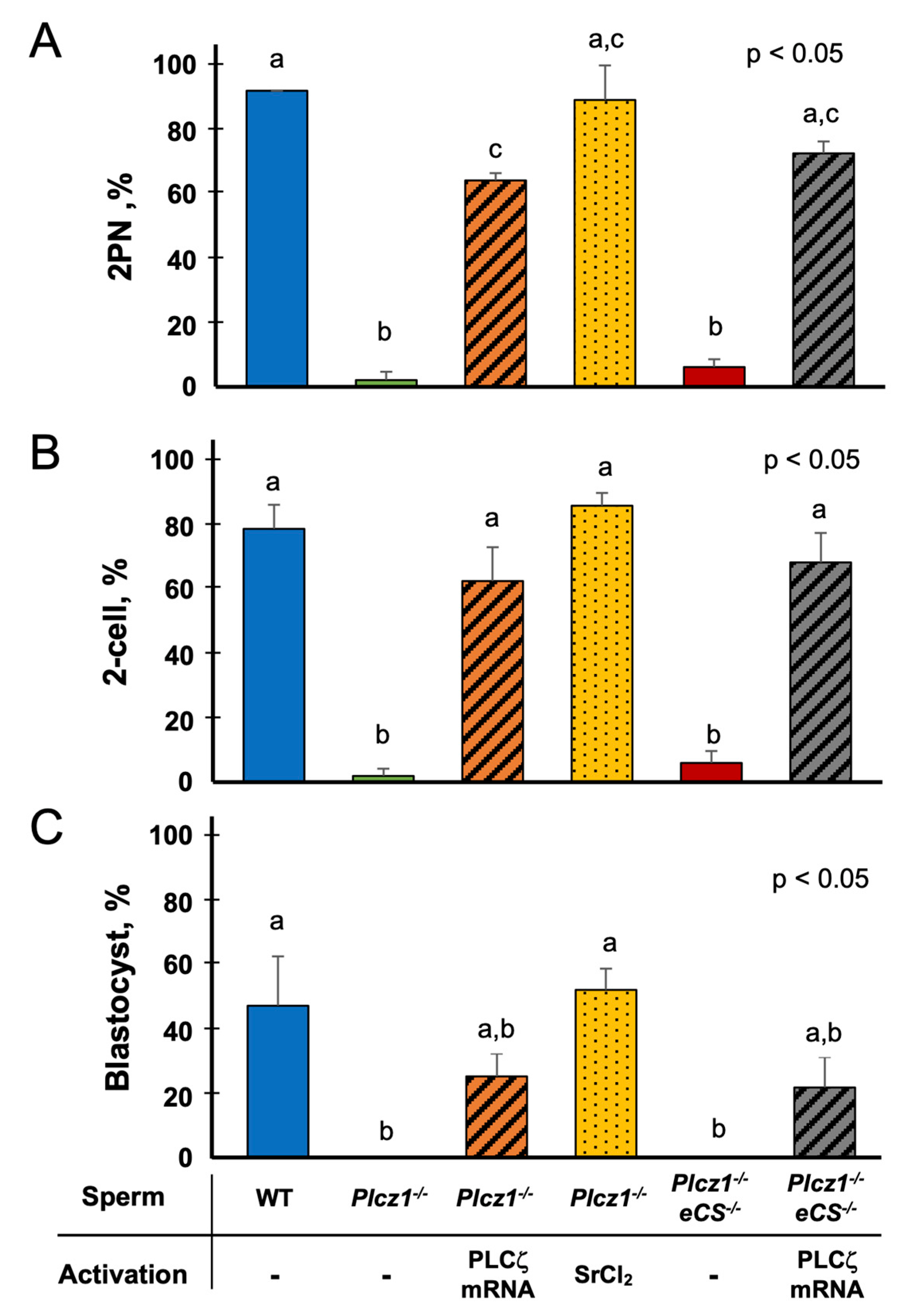
3.2. The Effects of the Different Regimens on the Activation and Embryonic Development of Oocytes Injected with Plcz1−/− or Plcz1−/−eCS−/− Sperm
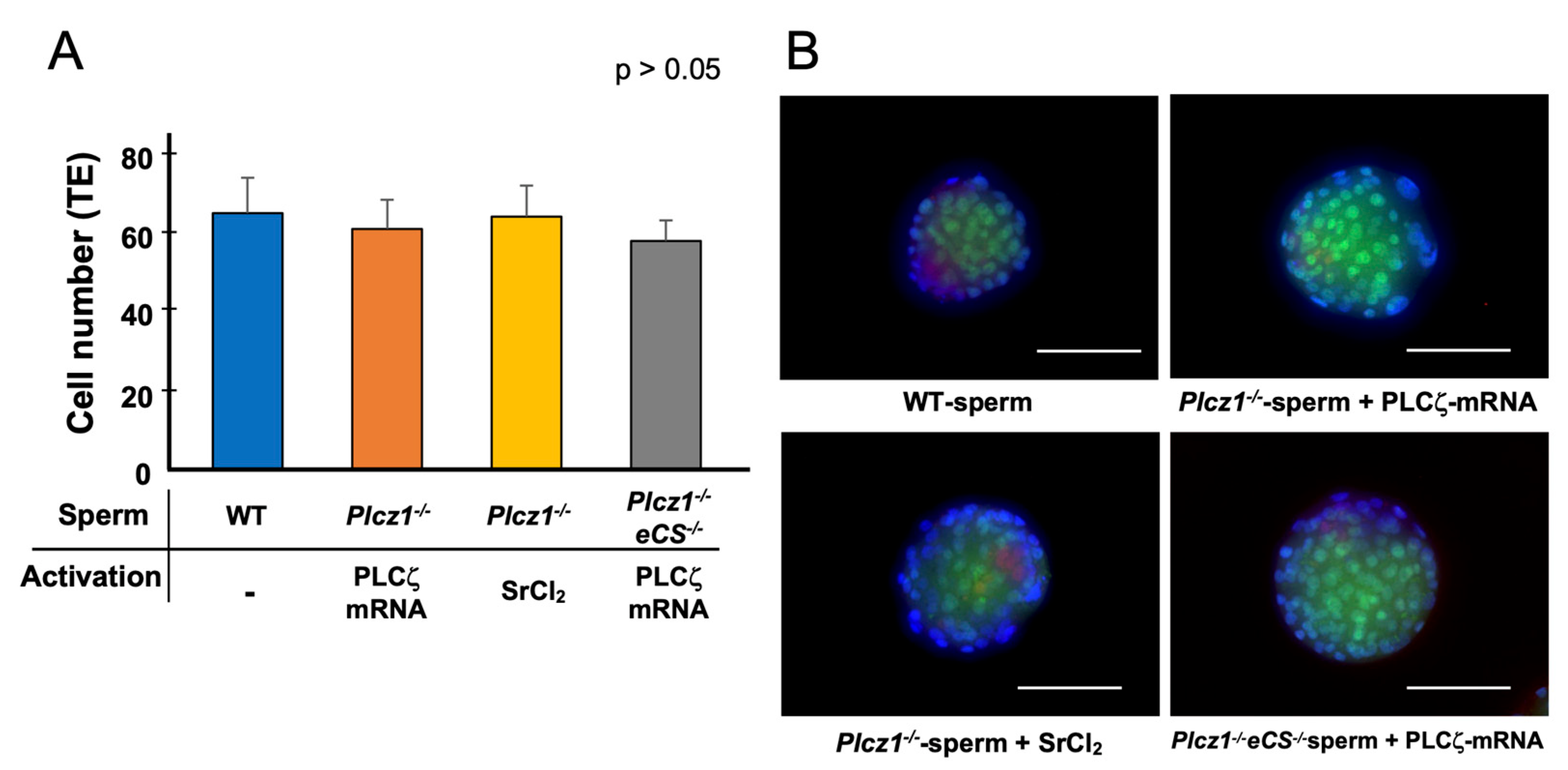
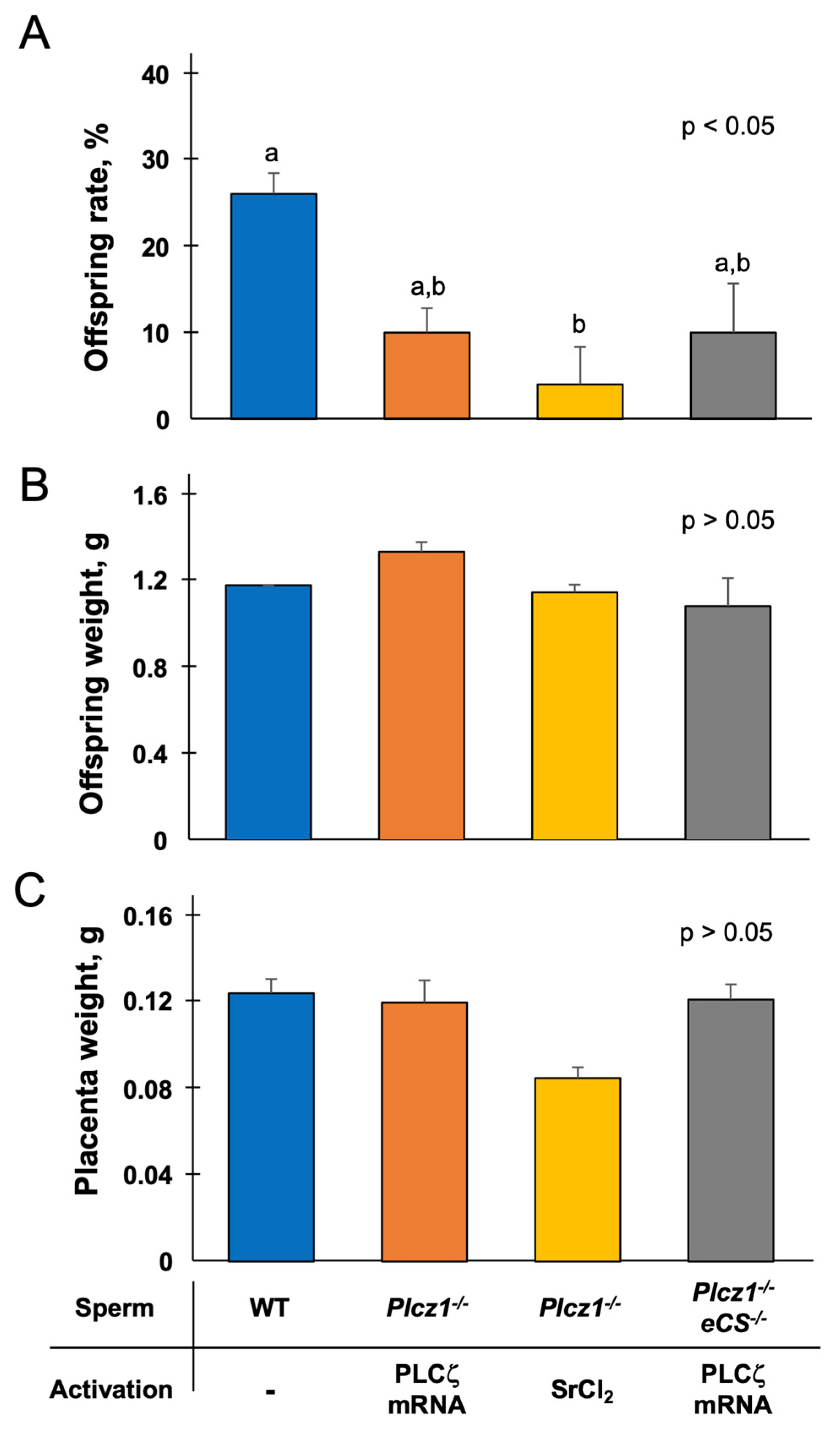
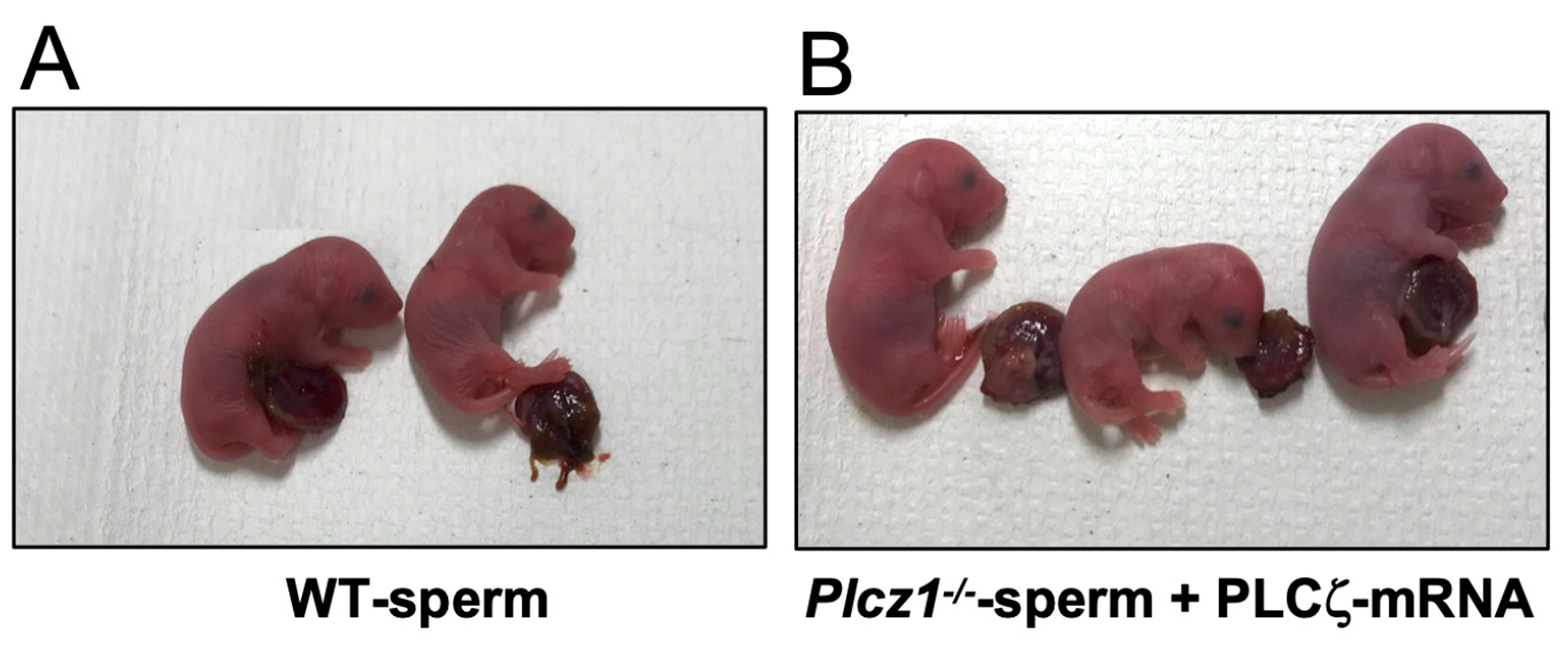
3.3. In Vivo Development of Embryos Derived from Plcz1−/− or Plcz1−/−eCS−/− Sperm by Different Activation Regimens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ito, J.; Parrington, J.; Fissore, R.A. PLCζ and its role as a trigger of development in verte-brates. Mol. Reprod. Dev. 2011, 78, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Ducibella, T.; Fissore, R. The roles of Ca2+, downstream protein kinases, and oscillatory signaling in regulating fertilization and the activation of development. Dev. Biol. 2008, 315, 257–279. [Google Scholar] [CrossRef] [PubMed]
- Schultz, R.M.; Kopf, G.S. Molecular basis of mammalian egg activation. Curr. Top. Dev. Biol. 1995, 30, 21–62. [Google Scholar] [PubMed]
- Saunders, C.M.; Larman, M.G.; Parrington, J.; Cox, L.J.; Royse, J.; Blayney, L.M.; Swann, K.; Lai, F.A. PLC zeta: A sperm-specific trigger of Ca(2+) oscillations in eggs and embryo development. Development 2002, 129, 3533–3544. [Google Scholar] [CrossRef]
- Ito, M.; Shikano, T.; Oda, S.; Horiguchi, T.; Tanimoto, S.; Awaji, T.; Mitani, H.; Miyazaki, S. Difference in Ca2+ oscillation-inducing activity and nuclear translocation ability of PLCZ1, an egg-activating sperm factor candidate, between mouse, rat, human, and medaka fish. Biol. Reprod. 2008, 78, 1081–1090. [Google Scholar] [CrossRef]
- Ito, M.; Shikano, T.; Kuroda, K.; Miyazaki, S. Relationship between nuclear sequestration of PLCzeta and termination of PLCzeta-induced Ca2+ oscillations in mouse eggs. Cell Calcium 2008, 44, 400–410. [Google Scholar] [CrossRef]
- Cox, L.J.; Larman, M.G.; Saunders, C.M.; Hashimoto, K.; Swann, K.; Lai, F.A. Sperm phospholipase Czeta from humans and cynomolgus monkeys triggers Ca2+ oscillations, activation and development of mouse oocytes. Reproduction 2002, 124, 611–623. [Google Scholar] [CrossRef]
- Cooney, M.A.; Malcuit, C.; Cheon, B.; Holland, M.K.; Fissore, R.A.; D’Cruz, N.T. Specific differences in the activity and nuclear localization of murine and bovine phospholipase C zeta 1. Biol. Reprod. 2010, 83, 92–101. [Google Scholar] [CrossRef]
- Malcuit, C.; Knott, J.G.; He, C.; Wainwright, T.; Parys, J.B.; Robl, J.M.; Fissore, R.A. Fertilization and inositol 1,4,5-trisphosphate (IP3)-induced calcium release in type-1 inositol 1,4,5-trisphosphate receptor down-regulated bovine eggs. Biol. Reprod. 2005, 73, 2–13. [Google Scholar] [CrossRef]
- Kurokawa, M.; Sato, K.; Fissore, R.A. Mammalian fertilization: From sperm factor to phospholipase Czeta. Biol. Cell 2004, 96, 37–45. [Google Scholar] [CrossRef]
- Yoneda, A.; Kashima, M.; Yoshida, S.; Terada, K.; Nakagawa, S.; Sakamoto, A.; Hayakawa, K.; Suzuki, K.; Ueda, J.; Watanabe, T. Molecular cloning, testicular postnatal expression, and oocyte-activating potential of porcine phospholipase Czeta. Reproduction 2006, 132, 393–401. [Google Scholar] [CrossRef]
- Sato, K.; Wakai, T.; Seita, Y.; Takizawa, A.; Fissore, R.A.; Ito, J.; Kashiwazaki, N. Molecular characteristics of horse phospholipase C zeta (PLCζ). Anim. Sci. J. 2013, 84, 359–368. [Google Scholar] [CrossRef]
- Nakai, M.; Ito, J.; Suyama, A.; Kageyama, A.; Tobari, Y.; Kashiwazaki, N. Phospholipase Czeta (PLCzeta) versus postacrosomal sheath WW domain-binding protein (PAWP): Which molecule will survive as a sperm factor? Anim. Sci. J. 2020, 91, e13345. [Google Scholar] [CrossRef]
- Kouchi, Z.; Fukami, K.; Shikano, T.; Oda, S.; Nakamura, Y.; Takenawa, T.; Miyazaki, S. Recombinant phospholipase Czeta has high Ca2+ sensitivity and induces Ca2+ oscillations in mouse eggs. J. Biol. Chem. 2004, 279, 10408–10412. [Google Scholar] [CrossRef]
- Heytens, E.; Parrington, J.; Coward, K.; Young, C.; Lambrecht, S.; Yoon, S.; Fissore, R.A.; Hamer, R.; Deane, C.M.; Ruas, M.; et al. Reduced amounts and abnormal forms of phospholipase C zeta (PLCzeta) in spermatozoa from infertile men. Hum. Reprod. 2009, 24, 2417–2428. [Google Scholar] [CrossRef]
- Yoon, S.; Jellerette, T.; Salicioni, A.M.; Lee, H.C.; Yoo, M.; Coward, K.; Parrington, J.; Grow, D.; Cibelli, J.B.; Visconti, P.E.; et al. Human sperm devoid of PLC, zeta 1 fail to induce Ca(2+) release and are unable to initiate the first step of embryo development. J. Clin. Invest. 2008, 118, 3671–3681. [Google Scholar] [CrossRef]
- Kashir, J.; Konstantinidis, M.; Jones, C.; Heindryckx, B.; De Sutter, P.; Parrington, J.; Wells, D.; Coward, K. Characterization of two heterozygous mutations of the oocyte activation factor phospholipase C zeta (PLCzeta) from an infertile man by use of minisequencing of individual sperm and expression in somatic cells. Fertil. Steril. 2012, 98, 423–431. [Google Scholar] [CrossRef]
- Escoffier, J.; Lee, H.C.; Yassine, S.; Zouari, R.; Martinez, G.; Karaouzene, T.; Coutton, C.; Kherraf, Z.; Halouani, L.; Triki, C.; et al. Homozygous mutation of PLCZ1 leads to defective human oocyte activation and infertility that is not rescued by the WW-binding protein PAWP. Hum. Mol. Genet. 2016, 25, 878–891. [Google Scholar] [CrossRef]
- Hachem, A.; Godwin, J.; Ruas, M.; Lee, H.C.; Ferrer Buitrago, M.; Ardestani, G.; Bassett, A.; Fox, S.; Navarrete, F.; de Sutter, P.; et al. PLCζ is the physiological trigger of the Ca2+ oscillations that induce embryogenesis in mammals but conception can occur in its absence. Development 2017, 144, 2914–2924. [Google Scholar]
- Nozawa, K.; Satouh, Y.; Fujimoto, T.; Oji, A.; Ikawa, M. Sperm-borne phospholipase C zeta-1 ensures monospermic fertilization in mice. Sci. Rep. 2018, 8, 1315. [Google Scholar] [CrossRef]
- Wilmut, I.; Schnieke, A.E.; McWhir, J.; Kind, A.J.; Campbell, K.H.S. Viable offspring derived from fetal and adult mammalian cells. Cloning Stem. Cells 2007, 9, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Tani, T.; Sotomaru, Y.; Kurokawa, K.; Kato, J.; Doguchi, H.; Yasue, H.; Tsunoda, Y. Eight calves cloned from somatic cells of a single adult. Science 1998, 282, 2095–2098. [Google Scholar] [CrossRef] [PubMed]
- Wakayama, T.; Perry, A.C.; Zuccotti, M.; Johnson, K.R.; Yanagimachi, R. Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature 1998, 394, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Onishi, A.; Iwamoto, M.; Akita, T.; Mikawa, S.; Takeda, K.; Awata, T.; Hanada, H.; Perry, A.C. Pig cloning by microinjection of fetal fibroblast nuclei. Science 2000, 289, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
- Kishigami, S.; Wakayama, S.; Nguyen, V.T.; Wakayama, T. Similar time restriction for intracytoplasmic sperm injection and round spermatid injection into activated oocytes for efficient offspring production. Biol. Reprod. 2004, 70, 1863–1869. [Google Scholar] [CrossRef]
- Ogura, A. Cloning Mice. Cold Spring Harb. Protoc. 2017, 2017, pdb.prot094425. [Google Scholar] [CrossRef]
- Kishigami, S.; Wakayama, T. Efficient strontium-induced activation of mouse oocytes in standard culture media by chelating calcium. J. Reprod. Dev. 2007, 53, 1207–1215. [Google Scholar] [CrossRef]
- Kang, W.; Harada, Y.; Yamatoya, K.; Kawano, N.; Kanai, S.; Miyamoto, Y.; Nakamura, A.; Miyado, M.; Hayashi, Y.; Kuroki, Y.; et al. Extra-mitochondrial citrate synthase initiates calcium oscillation and suppresses age-dependent sperm dysfunction. Lab. Invest. 2020, 100, 583–595. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Hirose, N.; Kamimura, S.; Wakayama, S.; Ito, J.; Ooga, M.; Wakayama, T. Production of mouse offspring from inactivated spermatozoa using horse PLCζ mRNA. J. Reprod. Dev. 2020, 66, 67–73. [Google Scholar] [CrossRef]
- Kimura, Y.; Yanagimachi, R. Intracytoplasmic sperm injection in the mouse. Biol. Reprod. 1995, 52, 709–720. [Google Scholar] [CrossRef]
- Chatot, C.L.; Ziomek, C.A.; Bavister, B.D.; Lewis, J.L.; Torres, I. An improved culture medium supports development of random-bred 1-cell mouse embryos in vitro. J. Reprod. Fertil. 1989, 86, 679–688. [Google Scholar] [CrossRef]
- Hirose, N.; Wakayama, S.; Inoue, R.; Ito, J.; Ooga, M.; Wakayama, T. Birth of offspring from spermatid or somatic cell by co-injection of PLCζ-cRNA. Reproduction 2020, 160, 319–330. [Google Scholar] [CrossRef]
- Swann, K. Sperm Factors and Egg Activation: PLCzeta as the sperm factor that activates eggs: 20 years on. Reproduction 2022, 164, E1–E4. [Google Scholar] [CrossRef]
- Harada, Y.; Matsumoto, T.; Hirahara, S.; Nakashima, A.; Ueno, S.; Oda, S.; Miyazaki, S.; Iwao, Y. Characterization of a sperm factor for egg activation at fertilization of the newt Cynops pyrrhogaster. Dev. Biol. 2007, 306, 797–808. [Google Scholar] [CrossRef]
- Chesné, P.; Adenot, P.G.; Viglietta, C.; Baratte, M.; Boulanger, L.; Renard, J. Cloned rabbits produced by nuclear transfer from adult somatic cells. Nat. Biotechnol. 2002, 20, 366–369. [Google Scholar] [CrossRef]
- Zhou, Q.; Renard, J.; Le Friec, G.; Brochard, V.; Beaujean, N.; Cherifi, Y.; Fraichard, A.; Cozzi, J. Generation of fertile cloned rats by regulating oocyte activation. Science 2003, 302, 1179. [Google Scholar] [CrossRef]
- Ogonuki, N.; Tsuchiya, H.; Hirose, Y.; Okada, H.; Ogura, A.; Sankai, T. Pregnancy by the tubal transfer of embryos developed after injection of round spermatids into oocyte cytoplasm of the cynomolgus monkey (Macaca fascicularis). Hum. Reprod. 2003, 18, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, M.; Kato, M.; Kitada, K.; Ohnami, N.; Hirao, M.; Hochi, S. Activation regimens for full-term development of rabbit oocytes injected with round spermatids. Mol. Reprod. Dev. 2009, 76, 573–579. [Google Scholar] [CrossRef]
- Ozil, J.; Banrezes, B.; Tóth, S.; Pan, H.; Schultz, R.M. Ca2+ oscillatory pattern in fertilized mouse eggs affects gene expression and development to term. Dev. Biol. 2006, 300, 534–544. [Google Scholar] [CrossRef]
- Wakai, T.; Zhang, N.; Vangheluwe, P.; Fissore, R.A. Regulation of endoplasmic reticulum Ca(2+) oscillations in mammalian eggs. J. Cell Sci. 2013, 126, 5714–5724. [Google Scholar]
- Anifandis, G.; Michopoulos, A.; Daponte, A.; Chatzimeletiou, K.; Simopoulou, M.; Messini, C.I.; Polyzos, N.P.; Vassiou, K.; Dafopoulos, K.; Goulis, D.G. Artificial oocyte activation: Physiological, pathophysiological and ethical aspects. Syst. Biol. Reprod. Med. 2019, 65, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.; Kashir, J.; Thanassoulas, A.; Safieh-Garabedian, B.; Lai, F.A.; Nomikos, M. Essential Role of Sperm-Specific PLC-Zeta in Egg Activation and Male Factor Infertility: An Update. Front. Cell Dev. Biol. 2020, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Thanassoulas, A.; Swann, K.; Lai, F.A.; Nomikos, M. Sperm Factors and Egg Activation: The structure and function relationship of sperm PLCZ1. Reproduction 2022, 164, F1–F8. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirose, N.; Kikuchi, Y.; Kageyama, A.; Sugita, H.; Sakurai, M.; Kawata, Y.; Terakawa, J.; Wakayama, T.; Ito, J.; Kashiwazaki, N. Successful Production of Offspring Derived from Phospholipase C Zeta-Deficient Sperm by Additional Artificial Activation. Life 2023, 13, 980. https://doi.org/10.3390/life13040980
Hirose N, Kikuchi Y, Kageyama A, Sugita H, Sakurai M, Kawata Y, Terakawa J, Wakayama T, Ito J, Kashiwazaki N. Successful Production of Offspring Derived from Phospholipase C Zeta-Deficient Sperm by Additional Artificial Activation. Life. 2023; 13(4):980. https://doi.org/10.3390/life13040980
Chicago/Turabian StyleHirose, Naoki, Yasuyuki Kikuchi, Atsuko Kageyama, Hibiki Sugita, Miu Sakurai, Yui Kawata, Jumpei Terakawa, Teruhiko Wakayama, Junya Ito, and Naomi Kashiwazaki. 2023. "Successful Production of Offspring Derived from Phospholipase C Zeta-Deficient Sperm by Additional Artificial Activation" Life 13, no. 4: 980. https://doi.org/10.3390/life13040980