
Alterations in Oxidative Stress Markers and Na,K-ATPase Enzyme Properties in Kidney after Fructose Intake and Quercetin Intervention in Rats

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
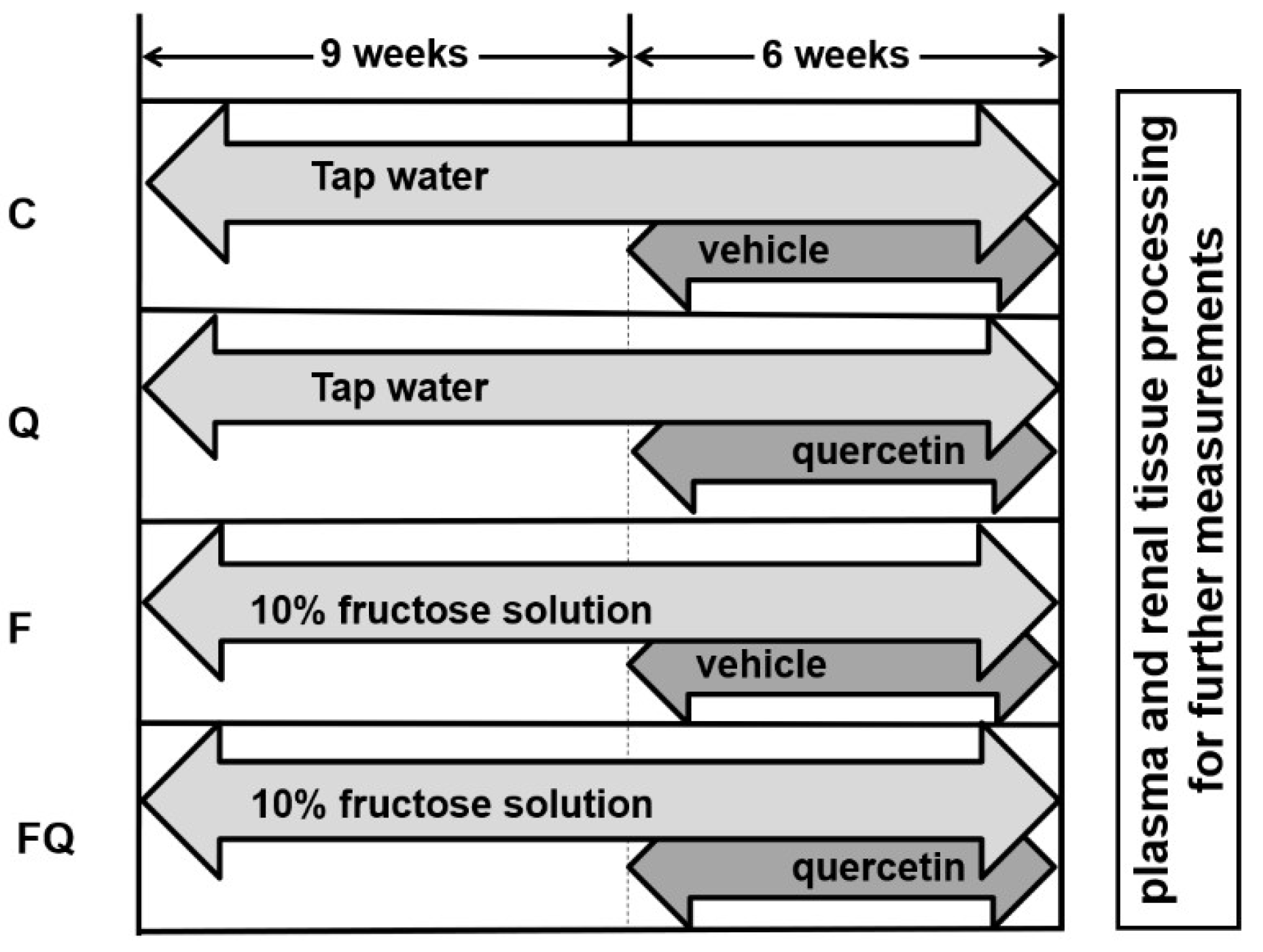
2.1. Experimental Model
2.2. Biochemical Analyses
2.3. Markers of Oxidative Stress, Carbonyl Stress, and Antioxidant Status
2.4. Na,K-ATPase-Enzyme Kinetic Measurements
2.5. Na,K-ATPase-Electrophoresis and Immunochemical Western Blot Analysis
2.6. Statistical Analysis
3. Results
3.1. Basic Characteristics of Experimental Animals
3.2. Biochemical Analysis of Blood Plasma
3.3. Markers of Antioxidant Status and Oxidative and Carbonyl Stress in the Renal Tissue
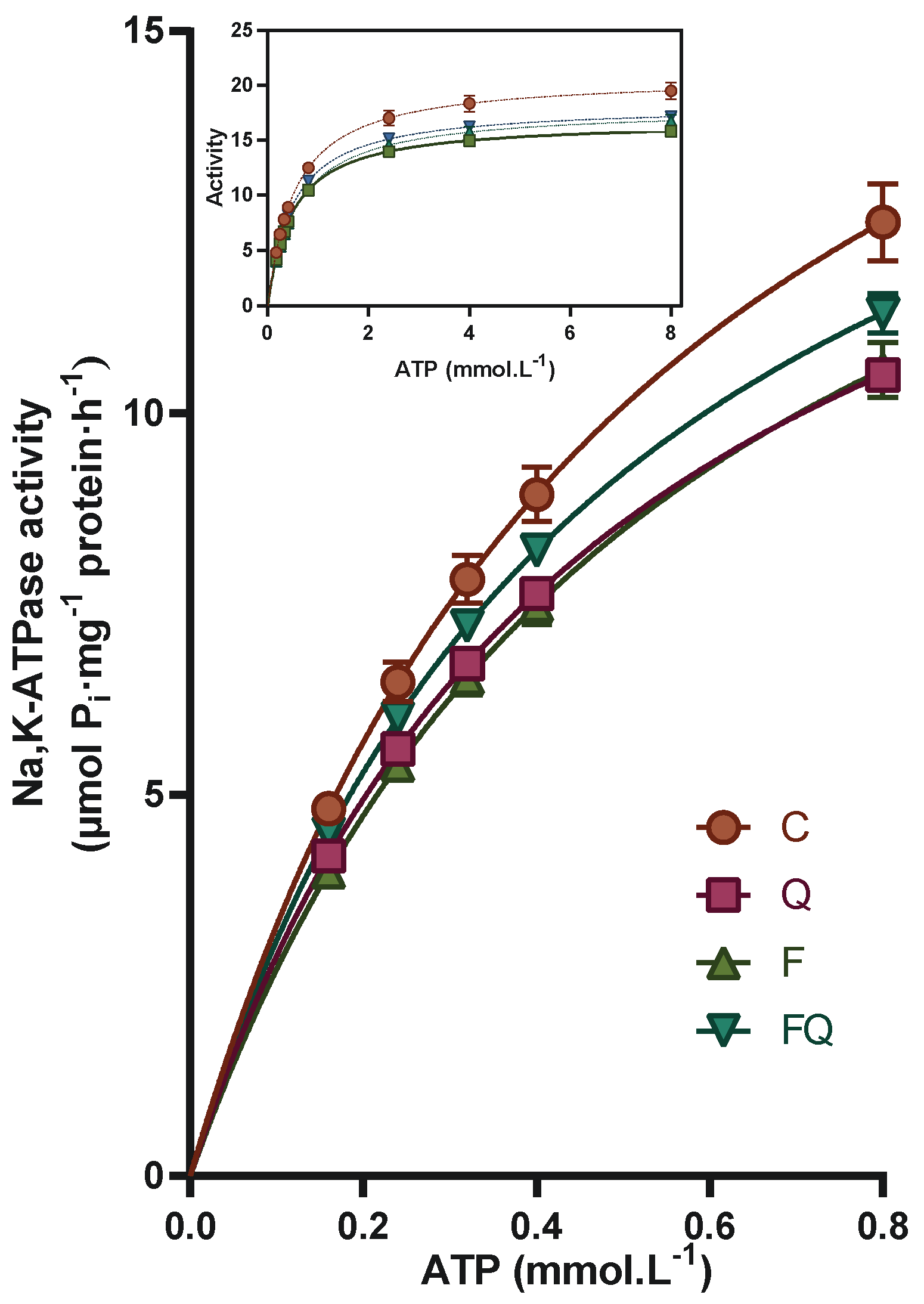
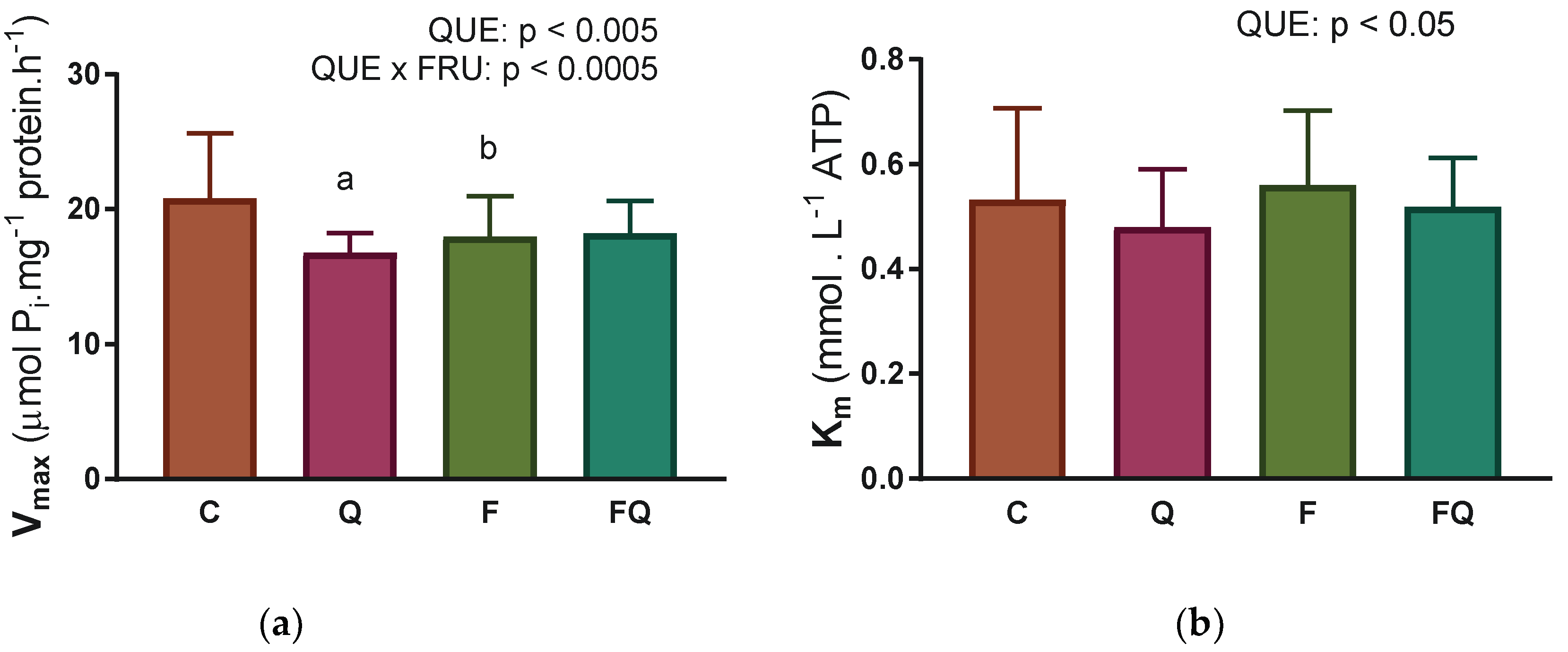
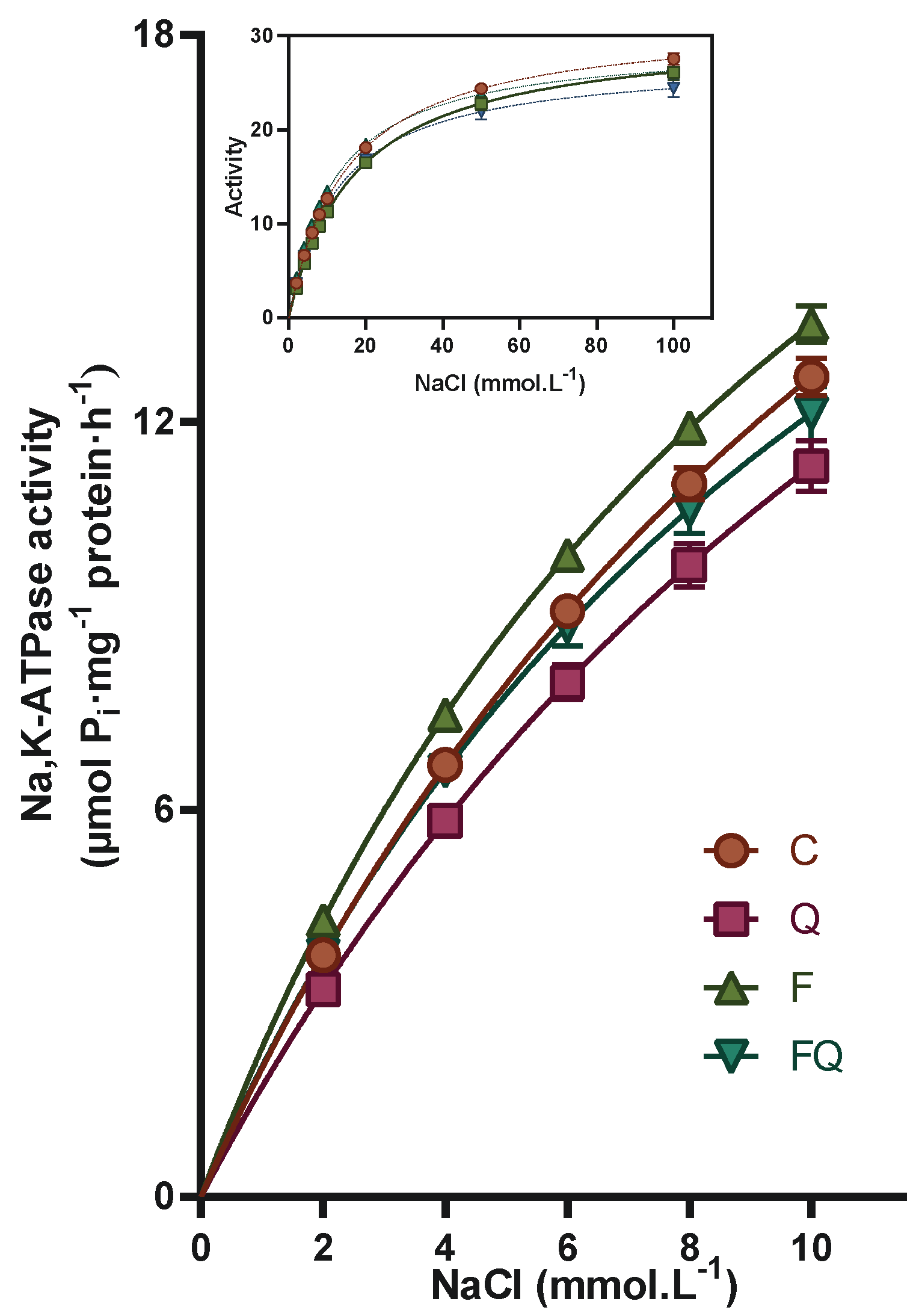
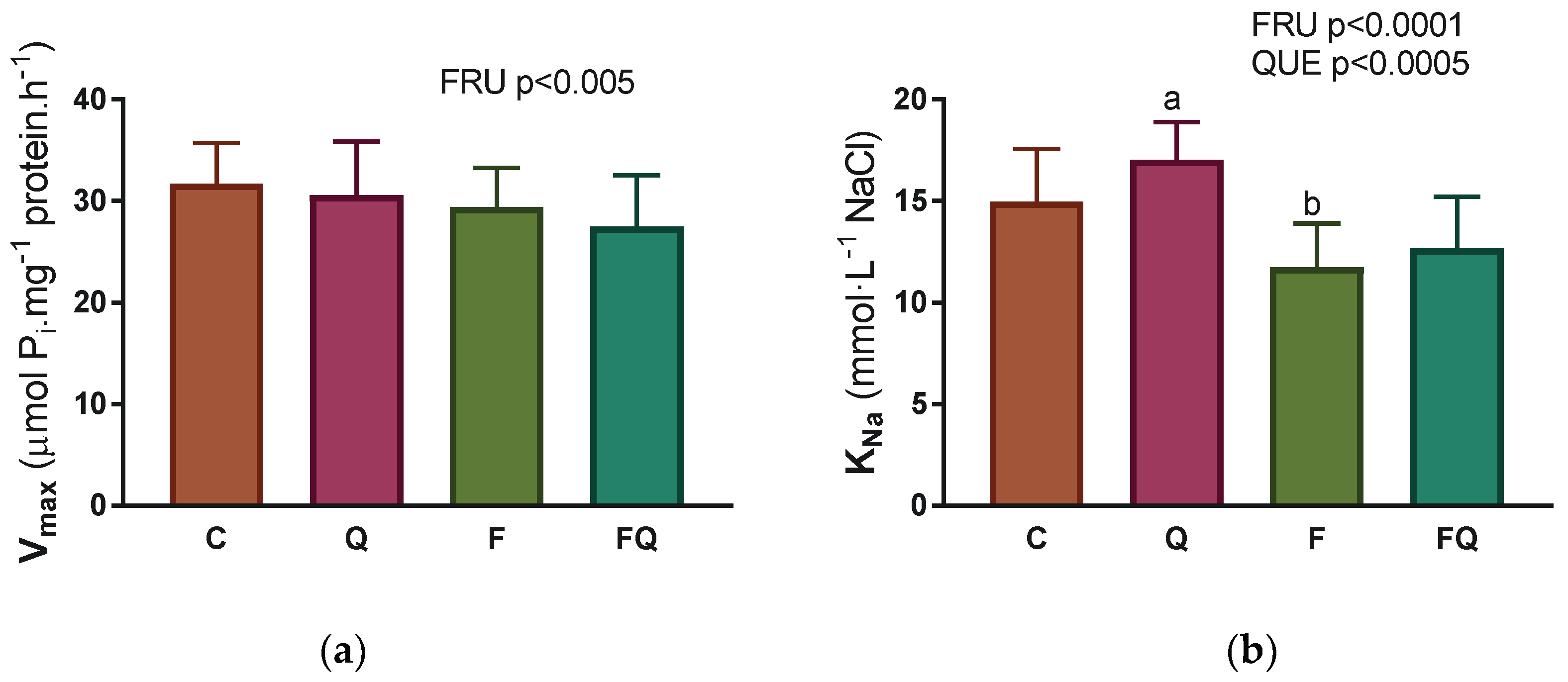
3.4. Renal Na,K-ATPase Enzyme Characterization of Kinetic Parameters
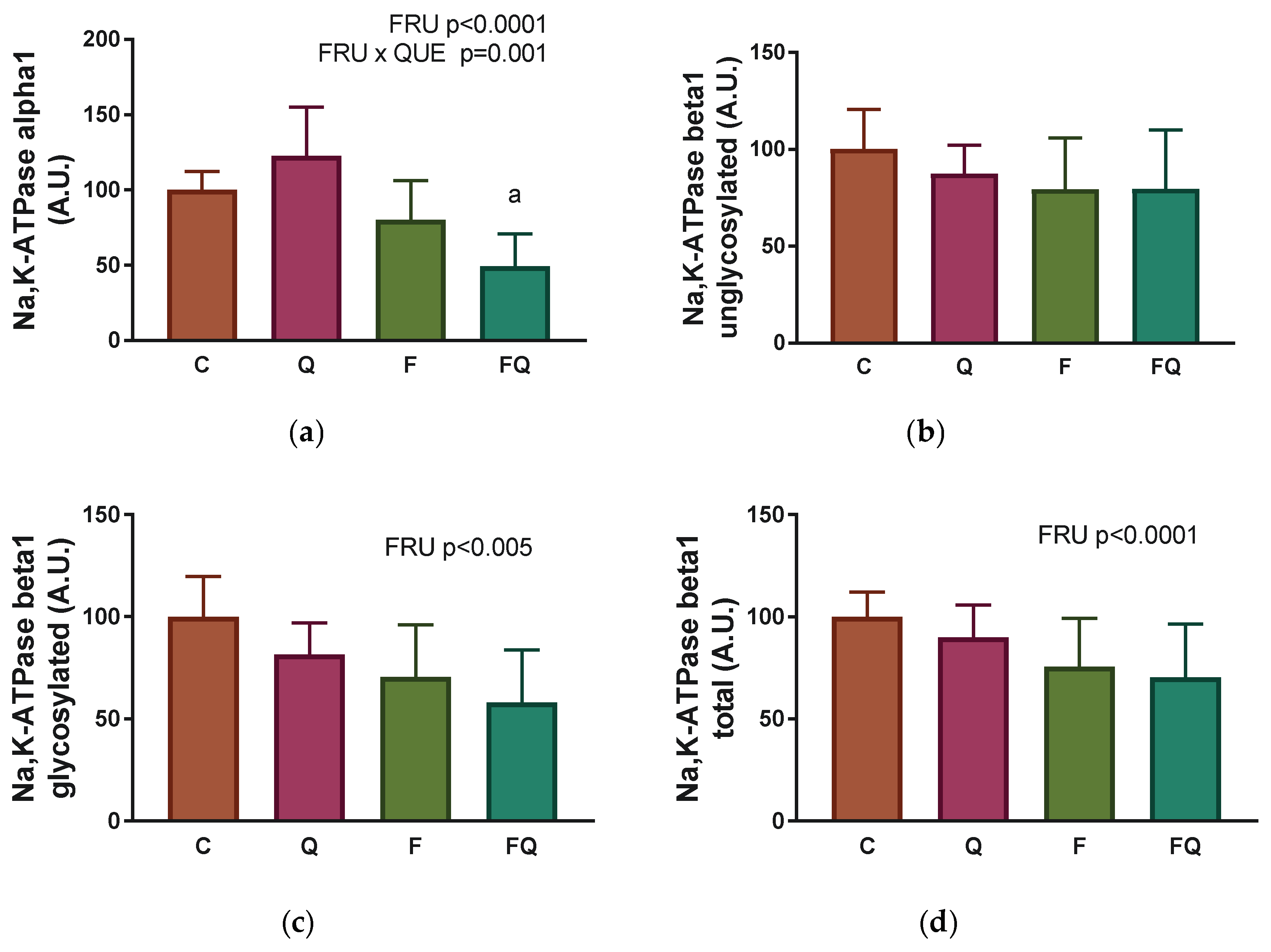
3.5. Renal Na,K-ATPase Enzyme-Subunit Expressions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Walker, R.W.; Dumke, K.A.; Goran, M.I. Fructose Content in Popular Beverages Made with and without High-Fructose Corn Syrup. Nutrition 2014, 30, 928–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiNicolantonio, J.J.; Lucan, S.C.; O’Keefe, J.H. The Evidence for Saturated Fat and for Sugar Related to Coronary Heart Disease. Prog. Cardiovasc. Dis. 2016, 58, 464–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mock, K.; Lateef, S.; Benedito, V.A.; Tou, J.C. High-Fructose Corn Syrup-55 Consumption Alters Hepatic Lipid Metabolism and Promotes Triglyceride Accumulation. J. Nutr. Biochem. 2017, 39, 32–39. [Google Scholar] [CrossRef]
- Taskinen, M.-R.; Söderlund, S.; Bogl, L.H.; Hakkarainen, A.; Matikainen, N.; Pietiläinen, K.H.; Räsänen, S.; Lundbom, N.; Björnson, E.; Eliasson, B.; et al. Adverse Effects of Fructose on Cardiometabolic Risk Factors and Hepatic Lipid Metabolism in Subjects with Abdominal Obesity. J. Intern. Med. 2017, 282, 187–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rukavina Mikusic, N.L.; Kouyoumdzian, N.M.; Uceda, A.; Del Mauro, J.S.; Pandolfo, M.; Gironacci, M.M.; Puyó, A.M.; Toblli, J.E.; Fernández, B.E.; Choi, M.R. Losartan Prevents the Imbalance between Renal Dopaminergic and Renin Angiotensin Systems Induced by Fructose Overload. l-Dopa/Dopamine Index as New Potential Biomarker of Renal Dysfunction. Metabolism 2018, 85, 271–285. [Google Scholar] [CrossRef]
- Gasparova, Z.; Ruskova, E.; Michalikova, D.S.; Brnoliakova, Z.; Svik, K.; Slovak, L.; Bezek, S.; Knezl, V.; Sotnikova, R. High-Fructose Intake-Induced Dyslipidemia and Oxidative Stress Accompanied by Hippocampal Dysfunctions in Hypertensive but Not Hypertriacylglycerolemic Rats. Gen. Physiol. Biophys. 2023, 42, 25–36. [Google Scholar] [CrossRef]
- Jurgoński, A.; Juśkiewicz, J.; Zduńczyk, Z.; Król, B. Caffeoylquinic Acid-Rich Extract from Chicory Seeds Improves Glycemia, Atherogenic Index, and Antioxidant Status in Rats. Nutrition 2012, 28, 300–306. [Google Scholar] [CrossRef]
- Nandhini, A.T.A.; Thirunavukkarasu, V.; Ravichandran, M.K.; Anuradha, C.V. Effect of Taurine on Biomarkers of Oxidative Stress in Tissues of Fructose-Fed Insulin-Resistant Rats. Singap. Med. J. 2005, 46, 82–87. [Google Scholar]
- Pasko, P.; Barton, H.; Zagrodzki, P.; Izewska, A.; Krosniak, M.; Gawlik, M.; Gawlik, M.; Gorinstein, S. Effect of Diet Sup-plemented with Quinoa Seeds on Oxidative Status in Plasma and Selected Tissues of High Fructose-Fed Rats. Plant Foods Hum. Nutr. 2010, 65, 146–151. [Google Scholar] [CrossRef]
- Rajasekar, P.; Viswanathan, P.; Anuradha, C.V. Renoprotective Action of L-Carnitine in Fructose-Induced Metabolic Syndrome: Renoprotective Action of CA. Diabetes. Obes. Metab. 2007, 10, 171–180. [Google Scholar] [CrossRef]
- Toklu, H.Z.; Şehirli, Ö.; Erşahin, M.; Süleymanoğlu, S.; Yiğiner, Ö.; Emekli-Alturfan, E.; Yarat, A.; Yeğen, B.Ç.; Şener, G. Resveratrol Improves Cardiovascular Function and Reduces Oxidative Organ Damage in the Renal, Cardiovascular and Cerebral Tissues of Two-Kidney, One-Clip Hypertensive Rats. J. Pharm. Pharmacol. 2010, 62, 1784–1793. [Google Scholar] [CrossRef] [PubMed]
- Vijaya Padma, V.; Kalai Selvi, P.; Sravani, S. Protective Effect of Ellagic Acid against TCDD-Induced Renal Oxidative Stress: Modulation of CYP1A1 Activity and Antioxidant Defense Mechanisms. Mol. Biol. Rep. 2014, 41, 4223–4232. [Google Scholar] [CrossRef] [PubMed]
- Kaločayová, B.; Kovačičová, I.; Radošinská, J.; Tóthová, Ľ.; Jagmaševič-Mézešová, L.; Fülöp, M.; Slezák, J.; Babál, P.; Janega, P.; Vrbjar, N. Alteration of Renal Na,K-ATPase in Rats Following the Mediastinal γ-Irradiation. Physiol. Rep. 2019, 7, e13969. [Google Scholar] [CrossRef] [Green Version]
- Hebbani, A.V.; Bulle, S.; Kanu, V.R.; Balachandrababu Malini, A.; Reddy, V.D.; Chakravarthula, V.N. Nephro-Protective Activity of Wheatgrass Juice against Alcohol-Induced Oxidative Damage in Rats. Toxicol. Mech. Methods 2020, 30, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Vicente, A.; Cabral, P.; Hong, N.; Asirwatham, J.; Yang, N.; Berthiaume, J.; Dominici, F.; Garvin, J. Dietary Fructose Enhances the Ability of Low Concentrations of Angiotensin II to Stimulate Proximal Tubule Na+ Reabsorption. Nutrients 2017, 9, 885. [Google Scholar] [CrossRef] [Green Version]
- Suganthi, R.; Rajamani, S.; Ravichandran, M.K.; Anuradha, C.V. Effect of Food Seasoning Spices Mixture on Biomarkers of Oxidative Stress in Tissues of Fructose-Fed Insulin-Resistant Rats. J. Med. Food 2007, 10, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Maithili Karpaga Selvi, N. Curcumin Attenuates Oxidative Stress and Activation of Redox-Sensitive Kinases in High Fructose-and High-Fat-Fed Male Wistar Rats. Sci. Pharm. 2015, 83, 159–175. [Google Scholar] [CrossRef] [Green Version]
- Fotschki, B.; Juśkiewicz, J.; Kołodziejczyk, K.; Jurgoński, A.; Kosmala, M.; Milala, J.; Ognik, K.; Zduńczyk, Z. Protective Effects of Ellagitannin-Rich Strawberry Extracts on Biochemical and Metabolic Disturbances in Rats Fed a Diet High in Fructose. Nutrients 2018, 10, 445. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Deng, H.; Guo, X.; Yan, S.; Lu, C.; Zhao, Z.; Feng, X.; Li, Q.; Wang, J.; Zeng, J.; et al. Effective Dose/Duration of Natural Flavonoid Quercetin for Treatment of Diabetic Nephropathy: A Systematic Review and Meta-Analysis of Rodent Data. Phytomedicine 2022, 105, 154348. [Google Scholar] [CrossRef]
- Heeba, G.H.; Mahmoud, M.E. Dual Effects of Quercetin in Doxorubicin-Induced Nephrotoxicity in Rats and Its Modulation of the Cytotoxic Activity of Doxorubicin on Human Carcinoma Cells: Effects of Quercetin on Dox-Induced Nephrotoxicity. Environ. Toxicol. 2014, 31, 624–636. [Google Scholar] [CrossRef]
- Hsieh, C.-L.; Peng, C.; Cheng, Y.-M.; Lin, L.-Y.; Ker, Y.-B.; Chang, C.-H.; Chen, K.-C.; Peng, R.Y. Quercetin and Ferulic Acid Aggravate Renal Carcinoma in Long-Term Diabetic Victims. J. Agric. Food Chem. 2010, 58, 9273–9280. [Google Scholar] [CrossRef] [PubMed]
- Stanišić, J.; Korićanac, G.; Ćulafić, T.; Romić, S.; Stojiljković, M.; Kostić, M.; Pantelić, M.; Tepavčević, S. Low Intensity Exercise Prevents Disturbances in Rat Cardiac Insulin Signaling and Endothelial Nitric Oxide Synthase Induced by High Fructose Diet. Mol. Cell. Endocrinol. 2016, 420, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Vrbjar, N.; Jasenovec, T.; Kollarova, M.; Snurikova, D.; Chomova, M.; Radosinska, D.; Shawkatova, I.; Tothova, L.; Ra-dosinska, J. Na,K-ATPase Kinetics and Oxidative Stress in Kidneys of Zucker Diabetic Fatty (Fa/Fa) Rats Depending on the Diabetes Severity—Comparison with Lean (Fa/+) and Wistar Rats. Biology 2022, 11, 1519. [Google Scholar] [CrossRef]
- Jasenovec, T.; Radosinska, D.; Kollarova, M.; Balis, P.; Zorad, S.; Vrbjar, N.; Bernatova, I.; Cacanyiova, S.; Tothova, L.; Ra-dosinska, J. Effects of Taxifolin in Spontaneously Hypertensive Rats with a Focus on Erythrocyte Quality. Life 2022, 12, 2045. [Google Scholar] [CrossRef] [PubMed]
- Jasenovec, T.; Radosinska, D.; Kollarova, M.; Vrbjar, N.; Balis, P.; Trubacova, S.; Paulis, L.; Tothova, L.; Shawkatova, I.; Radosinska, J. Monocrotaline-Induced Pulmonary Arterial Hypertension and Bosentan Treatment in Rats: Focus on Plasma and Erythrocyte Parameters. Pharmaceuticals 2022, 15, 1227. [Google Scholar] [CrossRef] [PubMed]
- Dobrocsyova, V.; Krskova, K.; Capcarova, M.; Zorad, S. Modulation of Adipogenesis and Oxidative Status by Quercetin and Ochratoxin A: Positive or Negative Impact on Rat Adipocyte Metabolism? Molecules 2019, 24, 3726. [Google Scholar] [CrossRef] [Green Version]
- Bartekova, M.; Radosinska, J.; Pancza, D.; Barancik, M.; Ravingerova, T. Cardioprotective Effects of Quercetin Against Ischemia-Reperfusion Injury Are Age-Dependent. Physiol. Res. 2016, 65, 101–107. [Google Scholar] [CrossRef]
- Mezesova, L.; Bartekova, M.; Javorkova, V.; Vlkovicova, J.; Breier, A.; Vrbjar, N. Effect of Quercetin on Kinetic Properties of Renal Na,K-ATPase in Normotensive and Hypertensive Rats. J. Physiol. Pharmacol. 2010, 61, 593–598. [Google Scholar]
- Jasenovec, T.; Radosinska, D.; Kollarova, M.; Balis, P.; Ferenczyova, K.; Kalocayova, B.; Bartekova, M.; Tothova, L.; Radosinska, J. Beneficial Effect of Quercetin on Erythrocyte Properties in Type 2 Diabetic Rats. Molecules 2021, 26, 4868. [Google Scholar] [CrossRef]
- Feyisa, T.O.; Melka, D.S.; Menon, M.; Labisso, W.L.; Habte, M.L. Investigation of the Effect of Coffee on Body Weight, Serum Glucose, Uric Acid and Lipid Profile Levels in Male Albino Wistar Rats Feeding on High-Fructose Diet. Lab. Anim. Res. 2019, 35, 29. [Google Scholar] [CrossRef]
- Eid, B.G.; Neamatallah, T.; Hanafy, A.; El-Bassossy, H.M.; Aldawsari, H.M.; Vemuri, K.; Makriyannis, A. Effects of the CB1 Receptor Antagonists AM6545 and AM4113 on Insulin Resistance in a High-Fructose High-Salt Rat Model of Metabolic Syndrome. Medicina 2020, 56, 573. [Google Scholar] [CrossRef]
- Alemán, M.N.; Sánchez, S.S.; Honoré, S.M. Daily Intake of Smallanthus Sonchifolius (Yacon) Roots Reduces the Progression of Non-Alcoholic Fatty Liver in Rats Fed a High Fructose Diet. Plant Foods Hum. Nutr. 2022, 77, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Gordish, K.L.; Kassem, K.M.; Ortiz, P.A.; Beierwaltes, W.H. Moderate (20%) Fructose-Enriched Diet Stimulates Salt-Sensitive Hypertension with Increased Salt Retention and Decreased Renal Nitric Oxide. Physiol. Rep. 2017, 5, e13162. [Google Scholar] [CrossRef] [PubMed]
- Stanhope, K.L.; Medici, V.; Bremer, A.A.; Lee, V.; Lam, H.D.; Nunez, M.V.; Chen, G.X.; Keim, N.L.; Havel, P.J. A Dose-Response Study of Consuming High-Fructose Corn Syrup–Sweetened Beverages on Lipid/Lipoprotein Risk Factors for Cardiovascular Disease in Young Adults. Am. J. Clin. Nutr. 2015, 101, 1144–1154. [Google Scholar] [CrossRef] [Green Version]
- Sigala, D.M.; Hieronimus, B.; Medici, V.; Lee, V.; Nunez, M.V.; Bremer, A.A.; Cox, C.L.; Price, C.A.; Benyam, Y.; Abdelhafez, Y.; et al. The Dose-Response Effects of Consuming High Fructose Corn Syrup-Sweetened Beverages on Hepatic Lipid Content and Insulin Sensitivity in Young Adults. Nutrients 2022, 14, 1648. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Hu, H.; Zharikov, S.; Tuttle, K.R.; Short, R.A.; Glushakova, O.; Ouyang, X.; Feig, D.I.; Block, E.R.; Herrera-Acosta, J.; et al. A Causal Role for Uric Acid in Fructose-Induced Metabolic Syndrome. Am. J. Physiol.-Ren. Physiol. 2006, 290, 625–631. [Google Scholar] [CrossRef] [Green Version]
- Russo, E.; Leoncini, G.; Esposito, P.; Garibotto, G.; Pontremoli, R.; Viazzi, F. Fructose and Uric Acid: Major Mediators of Cardiovascular Disease Risk Starting at Pediatric Age. Int. J. Mol. Sci. 2020, 21, 4479. [Google Scholar] [CrossRef]
- Derosa, G.; D’Angelo, A.; Maffioli, P. The Role of Selected Nutraceuticals in Management of Prediabetes and Diabetes: An Updated Review of the Literature. Phytother. Res. 2022, 36, 3709–3765. [Google Scholar] [CrossRef]
- Houtmeyers, A.; Duchateau, L.; Grünewald, B.; Hermans, K. Reference Intervals for Biochemical Blood Variables, Packed Cell Volume, and Body Temperature in Pet Rats (Rattus Norvegicus) Using Point-of-Care Testing. Vet. Clin. Pathol. 2016, 45, 669–679. [Google Scholar] [CrossRef]
- Gao, L.; Yuan, P.; Zhang, Q.; Fu, Y.; Hou, Y.; Wei, Y.; Zheng, X.; Feng, W. Taxifolin Improves Disorders of Glucose Metabolism and Water-Salt Metabolism in Kidney via PI3K/AKT Signaling Pathway in Metabolic Syndrome Rats. Life Sci. 2020, 263, 118713. [Google Scholar] [CrossRef]
- Huang, D.Y.; Boini, K.M.; Friedrich, B.; Metzger, M.; Just, L.; Osswald, H.; Wulff, P.; Kuhl, D.; Vallon, V.; Lang, F. Blunted Hypertensive Effect of Combined Fructose and High-Salt Diet in Gene-Targeted Mice Lacking Functional Serum- and Glu-cocorticoid-Inducible Kinase SGK1. American Journal of Physiology-Regulatory. Integr. Comp. Physiol. 2006, 290, 935–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welch, W.J. Intrarenal Oxygen and Hypertension. Clin. Exp. Pharm. Physiol 2006, 33, 1002–1005. [Google Scholar] [CrossRef] [PubMed]
- Vagin, O.; Tokhtaeva, E.; Sachs, G. The Role of the Β1 Subunit of the Na,K-ATPase and Its Glycosylation in Cell-Cell Adhesion. J. Biol. Chem. 2006, 281, 39573–39587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Yuan, C.; Clough, D.; Haddy, F.J.; Pamnani, M.B. Role of Digitalis-Like Substance in Experimental Insulin-Dependent Diabetes Mellitus Hypertension. J. Cardiovasc. Pharmacol. 1993, 22, 20–21. [Google Scholar] [CrossRef] [PubMed]
- Vlkovicová, J.; Javorková, V.; Stefek, M.; Kysel’ová, Z.; Gajdosíková, A.; Vrbjar, N. Effect of the Pyridoindole Antioxidant Stobadine on the Cardiac Na(+),K(+)-ATPase in Rats with Streptozotocin-Induced Diabetes. Gen. Physiol. Biophys 2006, 25, 111–124. [Google Scholar] [PubMed]
- Chen, S.; Yuan, C.; Clough, D.; Schooley, J.; Haddy, F.J.; Pamnani, M.B. Role of Digitalis-Like Substance in the Hypertension of Streptozotocin-Induced Diabetes in Reduced Renal Mass Rats. Am. J. Hypertens. 1993, 6, 397–406. [Google Scholar] [CrossRef]
- Seraphim, D.C.C.; Punaro, G.R.; de Oliveira Fernandes, T.; Ginoza, M.; Lopes, G.S.; Higa, E.M.S. Assessment of Fructose Overload in the Metabolic Profile and Oxidative/Nitrosative Stress in the Kidney of Senescent Female Rats. Exp. Gerontol. 2017, 99, 53–60. [Google Scholar] [CrossRef]
- Girard, A.; Madani, S.; Boukortt, F.; Cherkaoui-Malki, M.; Belleville, J.; Prost, J. Fructose-Enriched Diet Modifies Antioxidant Status and Lipid Metabolism in Spontaneously Hypertensive Rats. Nutrition 2006, 22, 758–766. [Google Scholar] [CrossRef]
- Boots, A.W.; Li, H.; Schins, R.P.F.; Duffin, R.; Heemskerk, J.W.M.; Bast, A.; Haenen, G.R.M.M. The Quercetin Paradox. Toxicol. Appl. Pharmacol. 2007, 222, 89–96. [Google Scholar] [CrossRef]
- Jazvinšćak Jembrek, M.; Čipak Gašparović, A.; Vuković, L.; Vlainić, J.; Žarković, N.; Oršolić, N. Quercetin Supplementation: Insight into the Potentially Harmful Outcomes of Neurodegenerative Prevention. Naunyn-Schmiedeberg’s Arch. Pharm. 2012, 385, 1185–1197. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Experimental Groups | Two-Way ANOVA | |||||
|---|---|---|---|---|---|---|---|
| C (n = 12) | Q (n = 12) | F (n = 12) | FQ (n = 12) | QUE | FRU | Int. | |
| BW start (g) | 54 ± 3 | 53 ± 5 | 53 ± 3 | 53 ± 4 | |||
| BW end (g) | 535 ± 37 | 539 ± 52 | 577 ± 42 | 572 ± 39 | ** | ||
| BW gain (g) | 481 ± 36 | 486 ± 50 | 524 ± 42 | 519 ± 40 | ** | ||
| Food intake (g/rat/day) | 24.5 ± 0.7 | 24.0 ± 0.8 | 17.8 ± 1.0 a | 19.3 ± 1.1 b | * | **** | *** |
| Liquid intake (mL/rat/day) | 61.0 ± 3.9 | 58.8 ± 7.3 | 81.8 ± 9.5 a | 58.1 ± 12.5 c | **** | *** | *** |
| Energy intake (kJ/rat/day) | 269 ± 8 | 264 ± 9 | 336 ± 27 a | 312 ± 30 d | * | **** | |
| KW (g) | 1.76 ± 0.16 | 1.67 ± 0.13 | 1.82 ± 0.18 | 1.78 ± 0.13 | * | ||
| KW/BW (mg/g) | 3.29 ± 0.23 | 3.10 ± 0.19 | 3.23 ± 0.17 | 3.12 ± 0.19 | * | ||
| Parameter | Experimental Groups | Two-Way ANOVA | |||||
|---|---|---|---|---|---|---|---|
| C (n = 12) | Q (n = 11) | F (n = 11) | FQ (n = 12) | QUE | FRU | Int. | |
| Glucose (mmol/L) | 6.45 ± 0.46 | 6.13 ± 0.49 | 7.15 ± 0.63 a | 5.93 ± 0.46 b | **** | ** | |
| Urea (mmol/L) | 6.3 ± 0.6 | 6.1 ± 0.7 | 6.7 ± 0.7 | 7.3 ± 1.5 | ** | ||
| Creatinine (μmol/L) | 35.1 ± 4.1 | 32.9 ± 4.9 | 34.5 ± 6.7 | 40.3 ± 5.0 c | * | * | |
| Sodium (mmol/L) | 141.2 ± 3.9 | 144.4 ± 1.8 | 147.9 ± 6.6 d | 146.3 ± 0.9 | *** | * | |
| Potassium (mmol/L) | 7.3 ± 0.5 | 7.1 ± 0.7 | 6.2 ± 0.7 e | 5.9 ± 0.5 | **** | ||
| Experimental Groups | Two-Way ANOVA | ||||||
|---|---|---|---|---|---|---|---|
| C (n = 11) | Q (n = 12) | F (n = 12) | FQ (n = 12) | QUE | FRU | Int. | |
| TBARS (μmol/L) | 4.6 ± 1.6 | 5.4 ± 1.2 | 6.3 ± 1.3 a | 6.4 ± 1.2 | *** | ||
| AOPP (μmol/g) | 3.3 ± 0.5 | 2.5 ± 0.2 b | 3.0 ± 0.4 | 2.4 ± 0.2 c | **** | ||
| AGEs (mg/g) | 7.9 ± 1.1 | 8.5 ± 0.6 | 7.6 ± 0.9 | 8.1 ± 1.0 | * | ||
| FRUC (μmol/g) | 32.1 ± 6.3 | 16.0 ± 2.0 | 41.1 ± 22.9 | 22.1 ± 7.6 d | *** | ||
| TAC (mmol/L) | 1.3 ± 0.3 | 1.2 ± 0.2 | 1.5 ± 0.2 | 1.4 ± 0.2 | ** | ||
| FRAP (mmol/L) | 1.06 ± 0.20 | 1.33 ± 0.24 e | 1.36 ± 0.18 f | 1.25 ± 0.12 | * | ** | |
| GSH/GSSG | 0.34 ± 0.04 | 0.31 ± 0.02 e | 0.30 ± 0.02 f | 0.28 ± 0.02 | *** | **** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vrbjar, N.; Vlkovicova, J.; Snurikova, D.; Kalocayova, B.; Zorad, S.; Culafic, T.; Tepavcevic, S.; Tothova, L.; Radosinska, D.; Kollarova, M.; et al. Alterations in Oxidative Stress Markers and Na,K-ATPase Enzyme Properties in Kidney after Fructose Intake and Quercetin Intervention in Rats. Life 2023, 13, 931. https://doi.org/10.3390/life13040931
Vrbjar N, Vlkovicova J, Snurikova D, Kalocayova B, Zorad S, Culafic T, Tepavcevic S, Tothova L, Radosinska D, Kollarova M, et al. Alterations in Oxidative Stress Markers and Na,K-ATPase Enzyme Properties in Kidney after Fructose Intake and Quercetin Intervention in Rats. Life. 2023; 13(4):931. https://doi.org/10.3390/life13040931
Chicago/Turabian StyleVrbjar, Norbert, Jana Vlkovicova, Denisa Snurikova, Barbora Kalocayova, Stefan Zorad, Tijana Culafic, Snezana Tepavcevic, Lubomira Tothova, Dominika Radosinska, Marta Kollarova, and et al. 2023. "Alterations in Oxidative Stress Markers and Na,K-ATPase Enzyme Properties in Kidney after Fructose Intake and Quercetin Intervention in Rats" Life 13, no. 4: 931. https://doi.org/10.3390/life13040931