

The Effects of Freezing Media on the Characteristics of Male and Female Chicken Primordial Germ Cell Lines
 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Maintenance of Domestic Fowl Experimental Stocks
2.3. Isolation, Establishment, and Maintenance of PGC Lines
2.4. Freezing and Thawing of PGC Lines
2.5. Measurements of Cell Number and Viability of PGC Lines
2.6. DNA Isolation and Sex Determination
2.7. RNA Isolation and Synthesis of cDNA
2.8. qPCR
2.9. Immunohistochemical Staining of PGCs
2.10. Statistical Analysis
3. Results
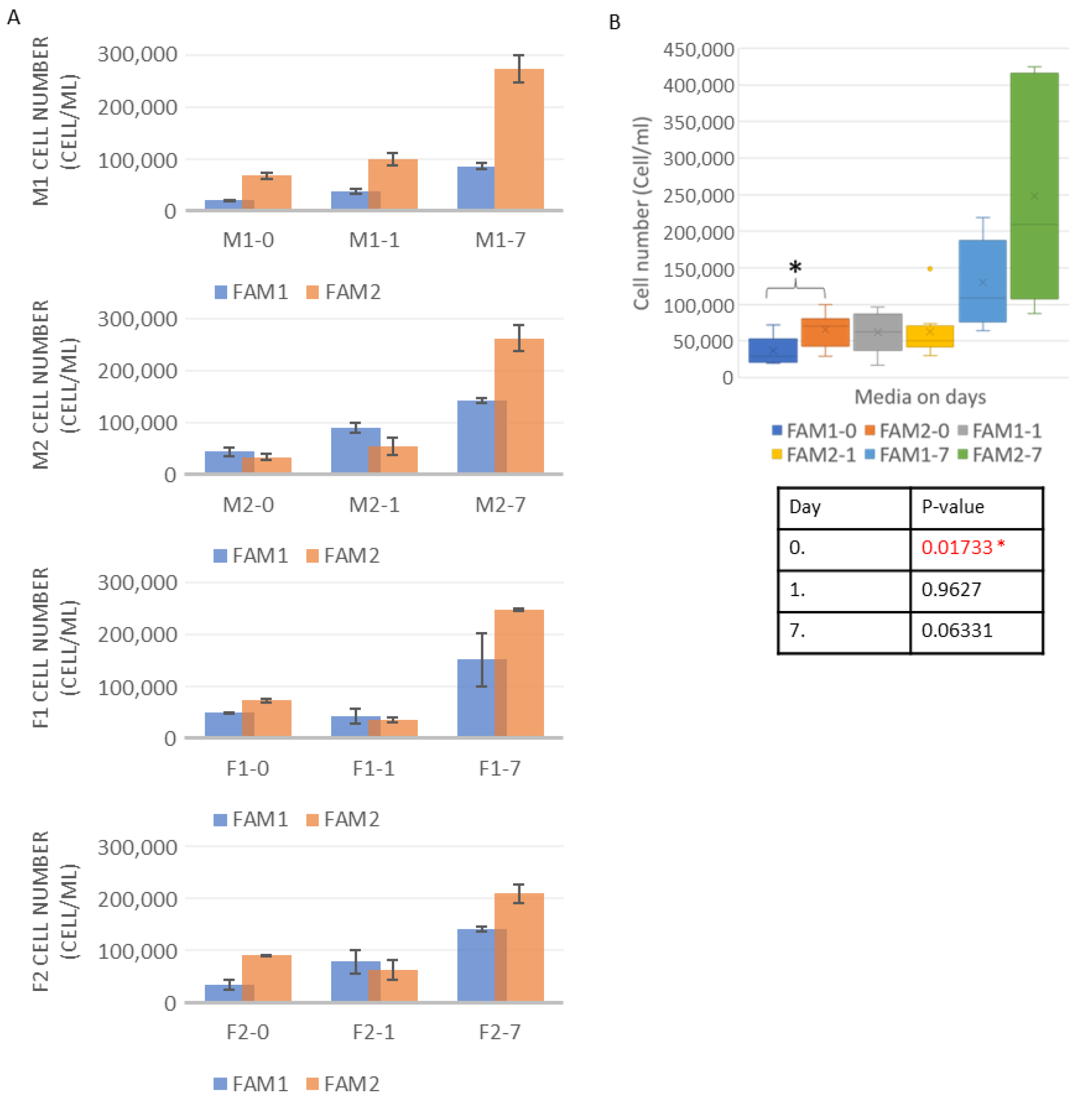
3.1. Cell Number Measurements
3.2. Cell Viability Measurements
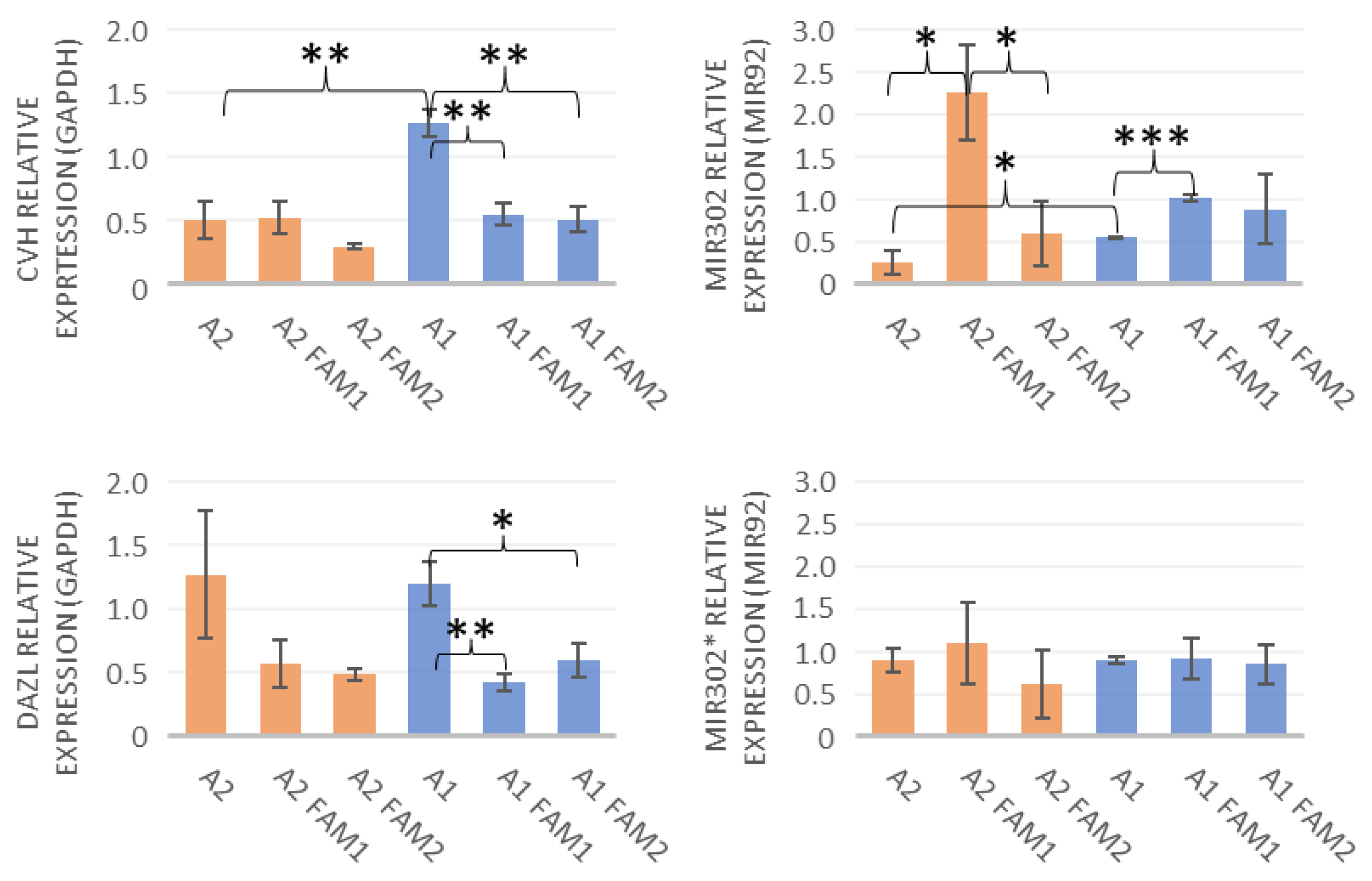
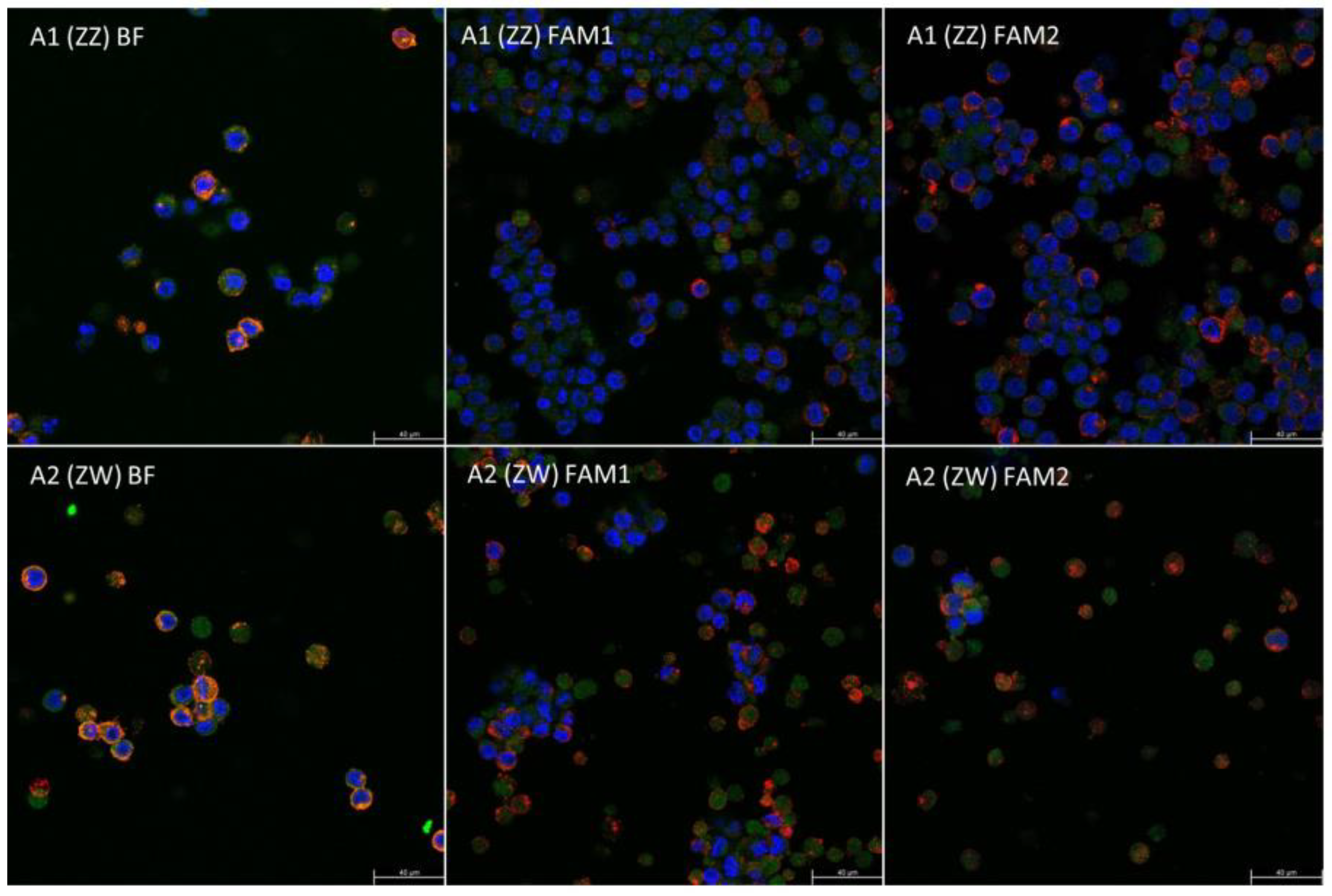
3.3. qPCR, MicroRNA Expression, and Immunohistochemical Staining
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sumner, J.H. Incredible goals of the world’s poultry industry. In Proceedings of the International Poultry Day Conference, Budapest, Hungary, 9–11 May 2018. [Google Scholar]
- Barna, J.; Liptói, K.; Barna, E.; Liptói, K.; Patakiné Várkonyi, E. Save what can be saved-new possibilities in in vitro gene preservation of poultry species (in Hungarian). Hungarian Vet. J. 2016, 138, 621–630. [Google Scholar]
- Liu, J.; Cheng, K.M.; Silversides, F.G. Production of live offspring from testicular tissue cryopreserved by vitrification procedures in Japanese quail (Coturnix japonica). Biol. Reprod. 2013, 88, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Cheng, K.M.; Silversides, F.G. Fundamental principles of cryobiology and application to ex situ conservation of avian species. Avian Biol. Res. 2013, 6, 187–197. [Google Scholar] [CrossRef]
- Silversides, F.G.; Robertson, M.C.; Liu, J. Cryoconservation of avian gonads in Canada. Poult. Sci. 2013, 92, 2613–2617. [Google Scholar] [CrossRef]
- Xiong, H.; Pu, Y.; Hu, Q.; Shan, Z.; Hu, P.; Guan, W.; Ma, Y. Embryoid bodies formation from chicken primordial germ cells. Anim. Cells Syst. 2015, 19, 168–174. [Google Scholar] [CrossRef]
- Eyal-Giladi, H.; Ginsburg, M.; Farbarov, A. Avian primordial germ cells are of epiblastic origin. J. Embryol. Exp. Morphol. 1981, 65, 139–147. [Google Scholar] [CrossRef]
- Kochav, S.; Ginsburg, M.; Eyal-Giladi, H. From Cleavage to Primitive Streak Formation: A Complementary Normal Table and a New Look at the First Stages of the Development of the Chick II. Microscopic Anatomy and Cell Population Dynamics. Dev. Biol. 1980, 79, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Yamamoto, Y.; Usui, F.; Mushika, T.; Ono, T.; Setioko, A.R.; Takeda, K.; Nirasawa, K.; Kagami, H.; Tagami, T. Migration and proliferation of primordial germ cells in the early chicken embryo. Poult. Sci. 2007, 86, 2182–2193. [Google Scholar] [CrossRef] [PubMed]
- Ginsburg, M.; Eyal-Giladi, H. Temporal and spatial aspects of the gradual migration of primordial germ cells from the epiblast into the germinal crescent in the avian embryo. J. Embryol. Exp. Morphol. 1986, 95, 53–71. [Google Scholar] [CrossRef]
- Naito, M.; Tajima, A.; Tagami, T.; Yasuda, Y.; Kuwana, T. Preservation of chick primordial germ cells in liquid nitrogen and subsequent production of viable offspring. J. Reprod. Fertil. 1994, 102, 321–325. [Google Scholar] [CrossRef] [Green Version]
- Naito, M. Cryopreservation of avian germline cells and subsequent production of viable offsprings. J. Poult. Sci. 2003, 40, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Setioko, A.R.; Tagami, T.; Tase, H.; Nakamura, Y.; Takeda, K.; Nirasawa, K. Cryopreservation of premordial germ cells (PGCs) from White Leghorn embryos using commercial cryoprotectants. J. Poult. Sci. 2007, 44, 73–77. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Usui, F.; Miyahara, D.; Mori, T.; Watanabe, H.; Ono, T.; Takeda, K.; Nirasawa, K.; Kagami, H.; Tagami, T. Viability and functionality of primordial germ cells after freeze-thaw in chickens. J. Poult. Sci. 2011, 48, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Nandi, S.; Whyte, J.; Taylor, L.; Sherman, A.; Nair, V.; Kaiser, P.; Mcgrew, M.J. Cryopreservation of specialized chicken lines using cultured primordial germ cells. Poult. Sci. 2016, 95, 1905–1911. [Google Scholar] [CrossRef]
- Kino, K.; Pain, B.; Leibo, S.P.; Cochran, M.; Clark, M.E.; Etches, R.J. Production of Chicken Chimeras from Injection of Frozen-Thawed Blastodermal Cells. Poult. Sci. 1997, 76, 753–760. [Google Scholar] [CrossRef]
- Ono, T.; Matsumoto, T.; Arisawa, Y. Production of Donor-Derived Offspring by Transfer of Primordial Germ Cells in Japanese Quail. Exp. Anim. 1998, 47, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Tajima, A.; Naito, M.; Yasuda, Y.; Kuwana, T. Production of germ-line chimeras by transfer of cryopreserved gonadal primordial germ cells (gPGCs) in chicken. J. Exp. Zool. 1998, 280, 265–267. [Google Scholar] [CrossRef]
- Trefil, P.; Aumann, D.; Koslová, A.; Mucksová, J.; Benešová, B.; Kalina, J.; Wurmser, C.; Fries, R.; Elleder, D.; Schusser, B.; et al. Male fertility restored by transplanting primordial germ cells into testes: A new way towards efficient transgenesis in chicken. Sci. Rep. 2017, 7, 14246. [Google Scholar] [CrossRef]
- Bednarczyk, M.; Dunislawska, A.; Stadnicka, K.; Grochowska, E. Chicken embryo as a model in epigenetic research. Poult. Sci. 2021, 100, 101164. [Google Scholar] [CrossRef]
- Petitte, J.N. Avian germplasm preservation: Embryonic stem cells or primordial germ cells? Poult. Sci. 2006, 85, 237–242. [Google Scholar] [CrossRef]
- Liu, J.; Cheng, K.M.; Silversides, F.G. Novel needle-in-straw vitrification can effectively preserve the follicle morphology, viability, and vascularization of ovarian tissue in Japanese quail (Coturnix japonica). Anim. Reprod. Sci. 2012, 134, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y. Poultry genetic resource conservation using primordial germ cells. J. Reprod. Dev. 2016, 62, 431–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whyte, J.; Glover, J.D.; Woodcock, M.; Brzeszczynska, J.; Taylor, L.; Sherman, A.; Kaiser, P.; McGrew, M.J. FGF, Insulin, and SMAD Signaling Cooperate for Avian Primordial Germ Cell Self-Renewal. Stem Cell Rep. 2015, 5, 1171–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.; Qiu, L.; Guo, Q.; Chen, Y.; Zhang, X.; Chen, B.; Zhang, Y.; Chang, G. Long-term in vitro culture and preliminary establishment of chicken primordial germ cell lines. PLoS ONE 2018, 13, e0196459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.F.; Kuwana, T. Purification of avian circulating primordial germ cells by Nycodenz density gradient centrifugation. Br. Poult. Sci. 2003, 44, 30–35. [Google Scholar] [CrossRef]
- Griffiths, R.; Double, M.C.; Orr, K.; Dawson, R.J.G. A DNA test to sex most birds. Mol. Ecol. 1998, 7, 1071–1075. [Google Scholar] [CrossRef]
- Lázár, B.; Anand, M.; Tóth, R.; Várkonyi, E.P.; Liptói, K.; Gócza, E. Comparison of the MicroRNA expression profiles of male and female avian primordial germ cell lines. Stem Cells Int. 2018, 2018, 1780679. [Google Scholar] [CrossRef] [Green Version]
- Chaipipat, S.; Sritabtim, K.; Piyasanti, Y.; Prukudom, S.; Jurutha, J.; Phetpila, V.; Sinsiri, R.; Kammongkun, J.; Molee, A.; Thiangtum, K.; et al. Initiative on Avian Primordial Germ Cell Cryobanking in Thailand. Biopreserv. Biobank. 2022; online ahead of print. [Google Scholar] [CrossRef]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef]
- Divya, D.; Shukla, R.; Chatterjee, R.; Sagar, G.; Rajendra Prasad, A.; Bhattacharya, T. Production of Transgenic Chimeric Chicken from Cryopreserved Primordial Germ Cells and its Validation by Developing shRNA Transgenic Chicken Chimera. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Tsunekawa, N.; Naito, M.; Sakai, Y.; Nishida, T.; Noce, T. Isolationof chicken vasa homolog gene and tracing the origin of primordial germ cells. Development 2000, 127, 2741–2750. [Google Scholar] [CrossRef]
- Kim, Y.M.; Han, J.Y. The early development of germ cells in chicken. Int. J. Dev. Biol. 2018, 62, 141–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lázár, B.; Molnár, M.; Sztán, N.; Végi, B.; Drobnyák, Á.; Tóth, R.; Tokodyné Szabadi, N.; McGrew, M.J.; Gócza, E.; Patakiné Várkonyi, E. Successful cryopreservation and regeneration of a partridge colored Hungarian native chicken breed using primordial germ cells. Poult. Sci. 2021, 100, 101207. [Google Scholar] [CrossRef] [PubMed]
- Ezaki, R.; Hirose, F.; Furusawa, S.; Horiuchi, H. An improved protocol for stable and efficient culturing of chicken primordial germ cells using small-molecule inhibitors. Cytotechnology 2020, 72, 397–405. [Google Scholar] [CrossRef]
- Lejong, M.; Choa-Duterre, M.; Vanmuylder, N.; Louryan, S. Is Vasa such a highly specific marker for primordial germ cells? A comparison of VASA and HSP90 proteins expression in young chicken embryos. Morphologie 2020, 104, 20–26. [Google Scholar] [CrossRef]
- Mazzoleni, S.; Němec, P.; Albrecht, T.; Lymberakis, P.; Kratochvíl, P.; Rovatsos, M. Long-term stability of sex chromosome gene content allows accurate qPCR-based molecular sexing across birds. Mol. Ecol. 2021, 21, 2013–2021. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FAM1 | |
| Avian KO DMEM | 3665 µL |
| Na Pyruvate | 20 µL |
| Chicken serum 10% | 500 µL |
| CaCl2 | 15 µL |
| DMSO | 800 µL |
| FAM2 | |
| FBS | 4000 µL |
| DMSO | 1000 µL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ecker, A.; Lázár, B.; Tóth, R.I.; Urbán, M.; Tokodyné Szabadi, N.; Salinas Aponte, M.T.; Adnan, M.; Várkonyi, E.; Gócza, E. The Effects of Freezing Media on the Characteristics of Male and Female Chicken Primordial Germ Cell Lines. Life 2023, 13, 867. https://doi.org/10.3390/life13040867
Ecker A, Lázár B, Tóth RI, Urbán M, Tokodyné Szabadi N, Salinas Aponte MT, Adnan M, Várkonyi E, Gócza E. The Effects of Freezing Media on the Characteristics of Male and Female Chicken Primordial Germ Cell Lines. Life. 2023; 13(4):867. https://doi.org/10.3390/life13040867
Chicago/Turabian StyleEcker, András, Bence Lázár, Roland Imre Tóth, Martin Urbán, Nikolett Tokodyné Szabadi, Maria Teresa Salinas Aponte, Mohd Adnan, Eszter Várkonyi, and Elen Gócza. 2023. "The Effects of Freezing Media on the Characteristics of Male and Female Chicken Primordial Germ Cell Lines" Life 13, no. 4: 867. https://doi.org/10.3390/life13040867