

Increased Trypanosoma cruzi Growth during Infection of Macrophages Cultured on Collagen I Matrix

 , , , and
, , , and 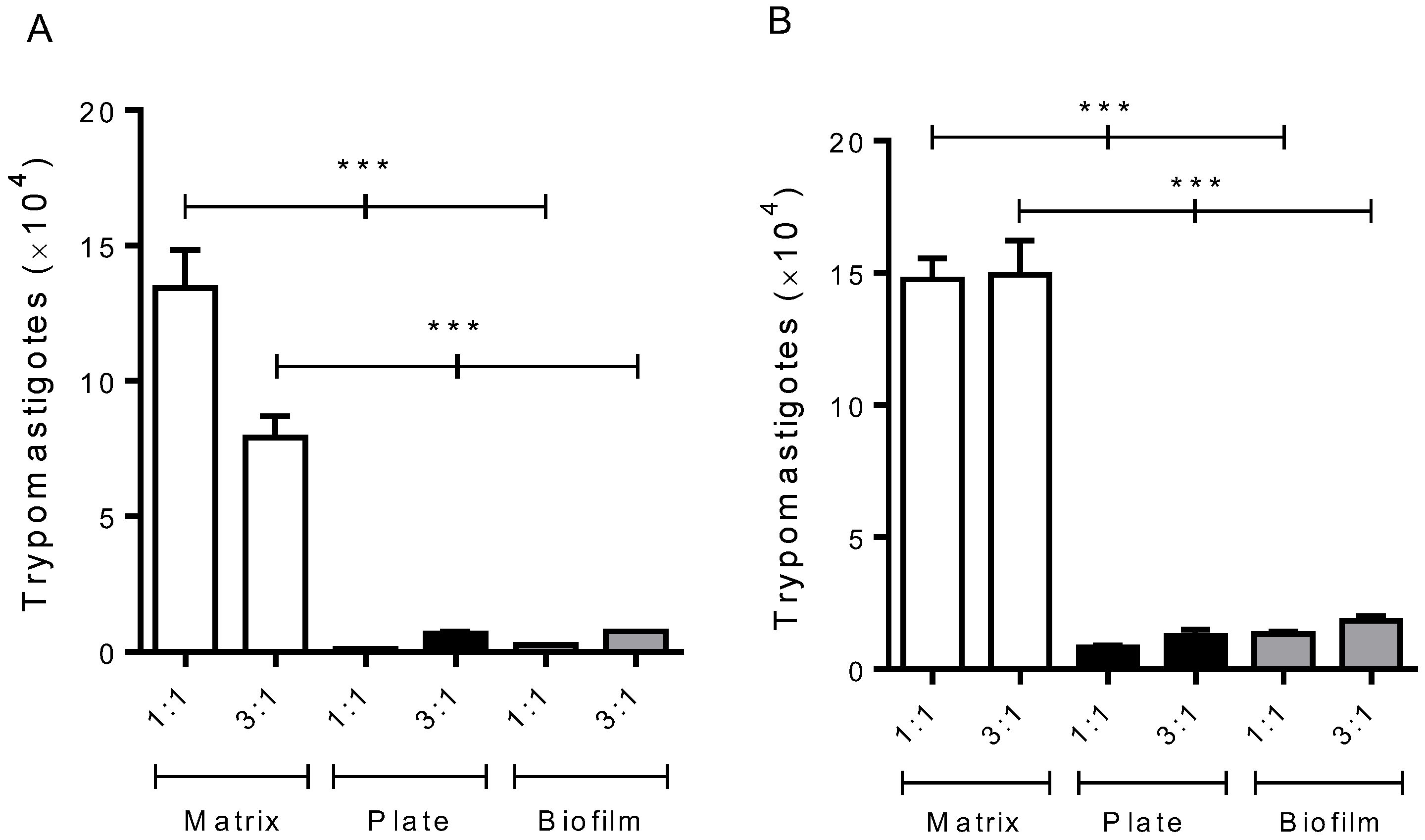{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Parasites
2.3. Collagen I Source
2.4. Macrophage Harvesting
2.5. Interaction Models In Vitro
2.5.1. D Collagen Type I Biofilm
2.5.2. D Collagen Type I Matrix
2.6. Parasitic Burden Assay
2.7. Determination of Cytokines
2.8. Determination of Nitric Oxide
2.9. Scanning Electron Microscopy (SEM)
2.10. Statistical Analysis
3. Results
3.1. Infected Macrophages Grown in a 3D Environment Release More Trypomastigotes Forms
3.2. Infected Macrophages Cultured on a 3D Matrix System Release More Amastigotes into the Extracellular Environment
3.3. Culture of Infected Macrophages on the 3D Matrix Induces the Early Release of Trypomastigote Forms
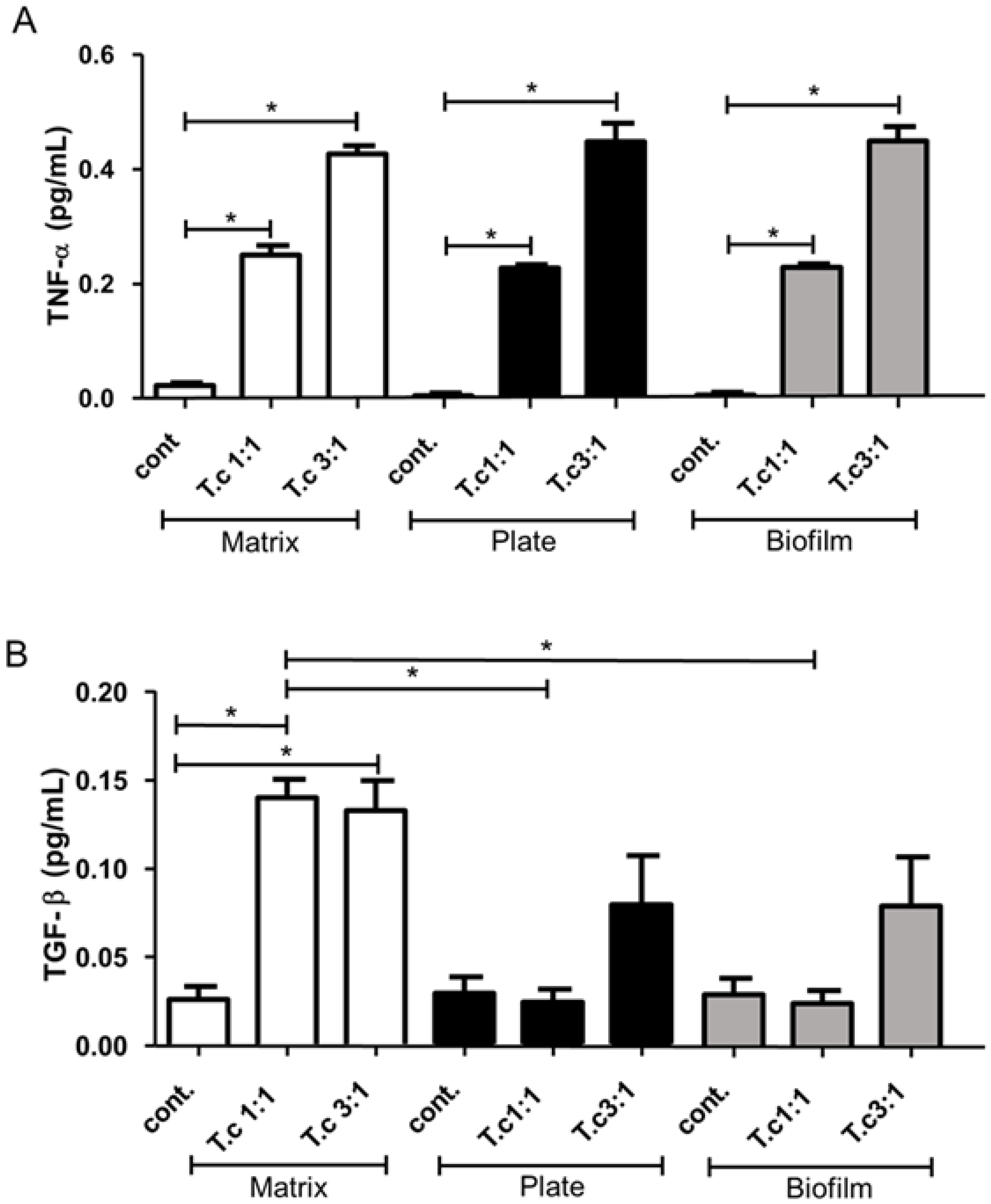
3.4. Detection of TNF-α and TGF-β
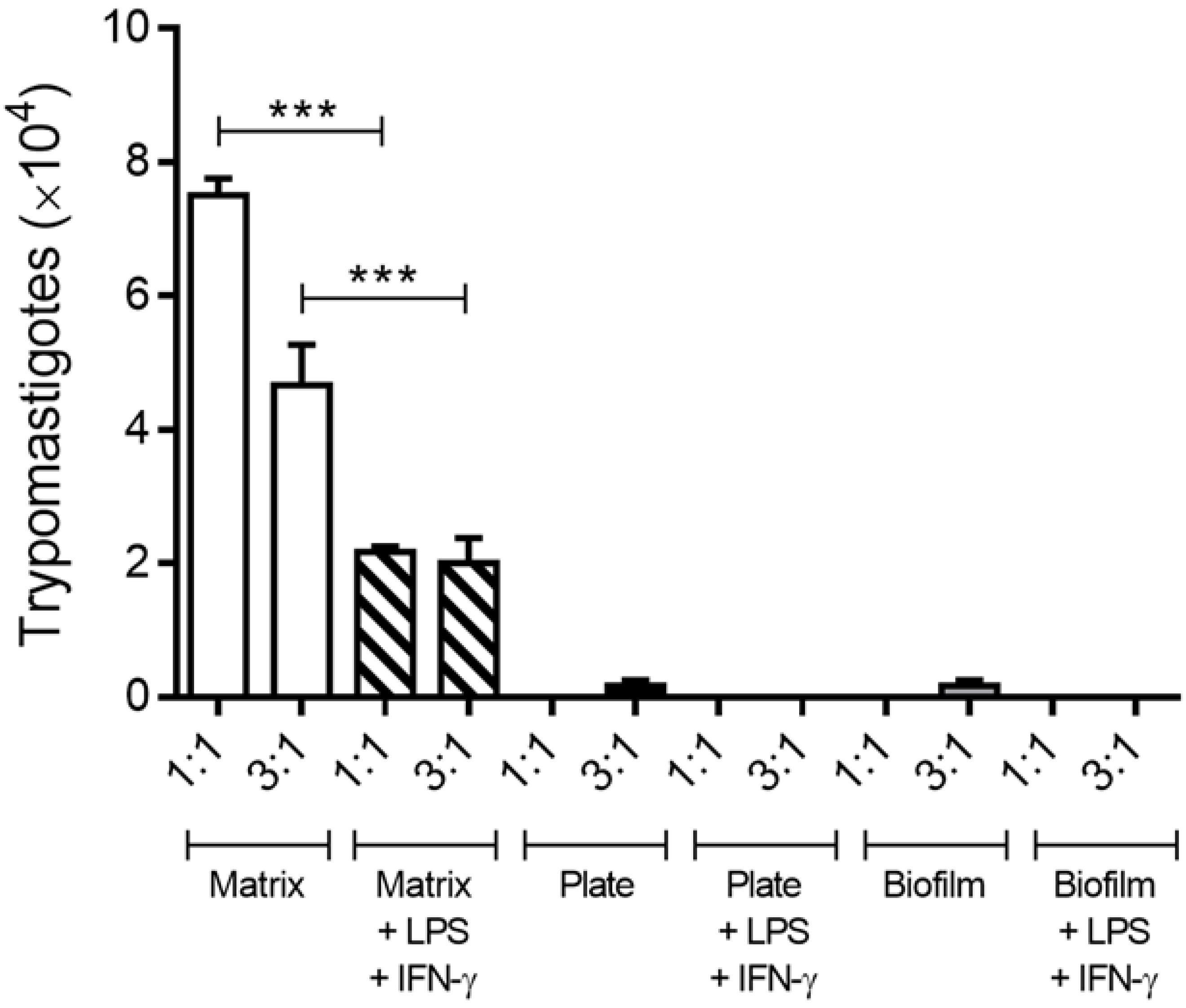
3.5. Activated Macrophages Reduced Trypomastigote Release in the 3D Matrix In Vitro
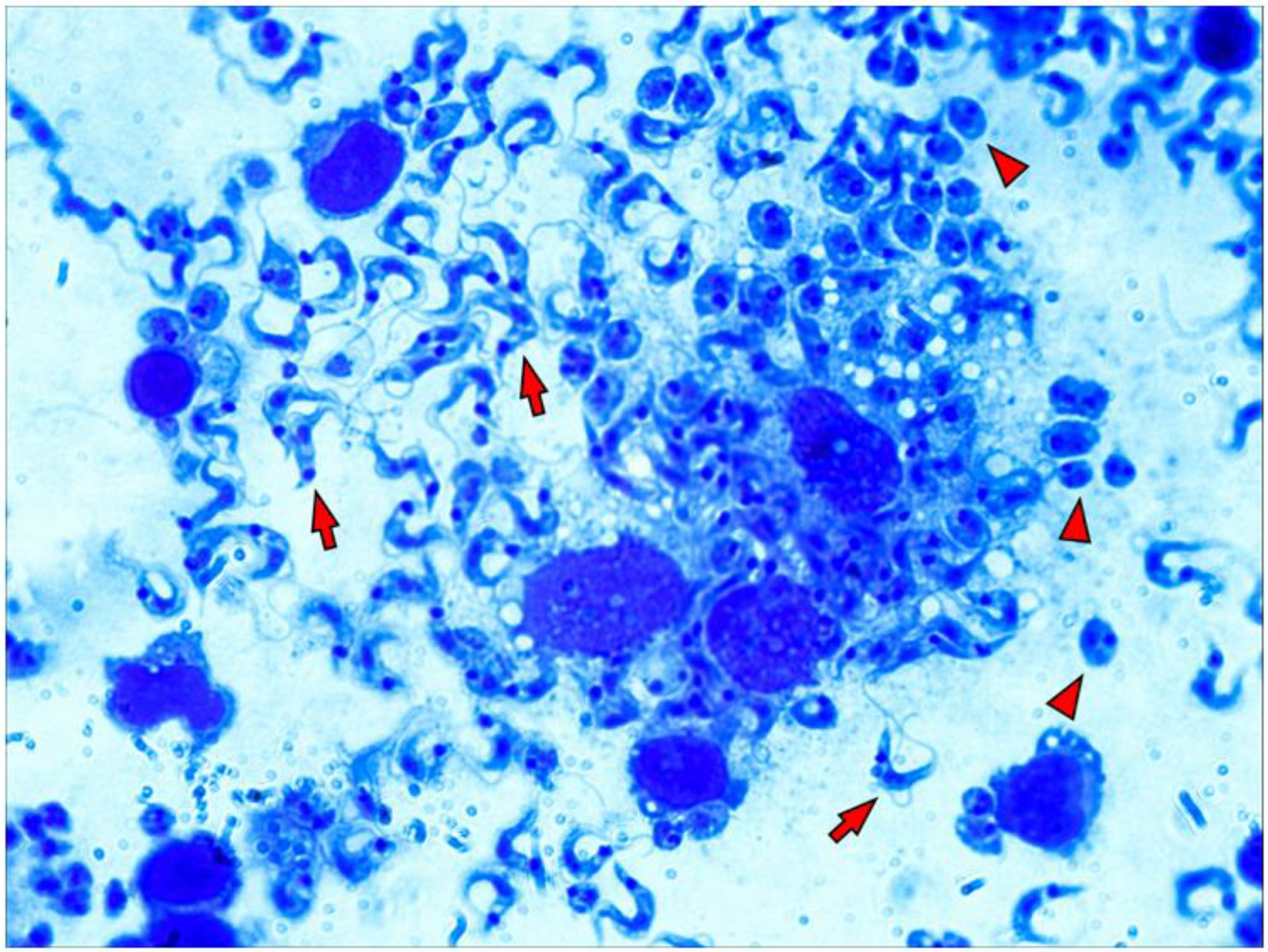
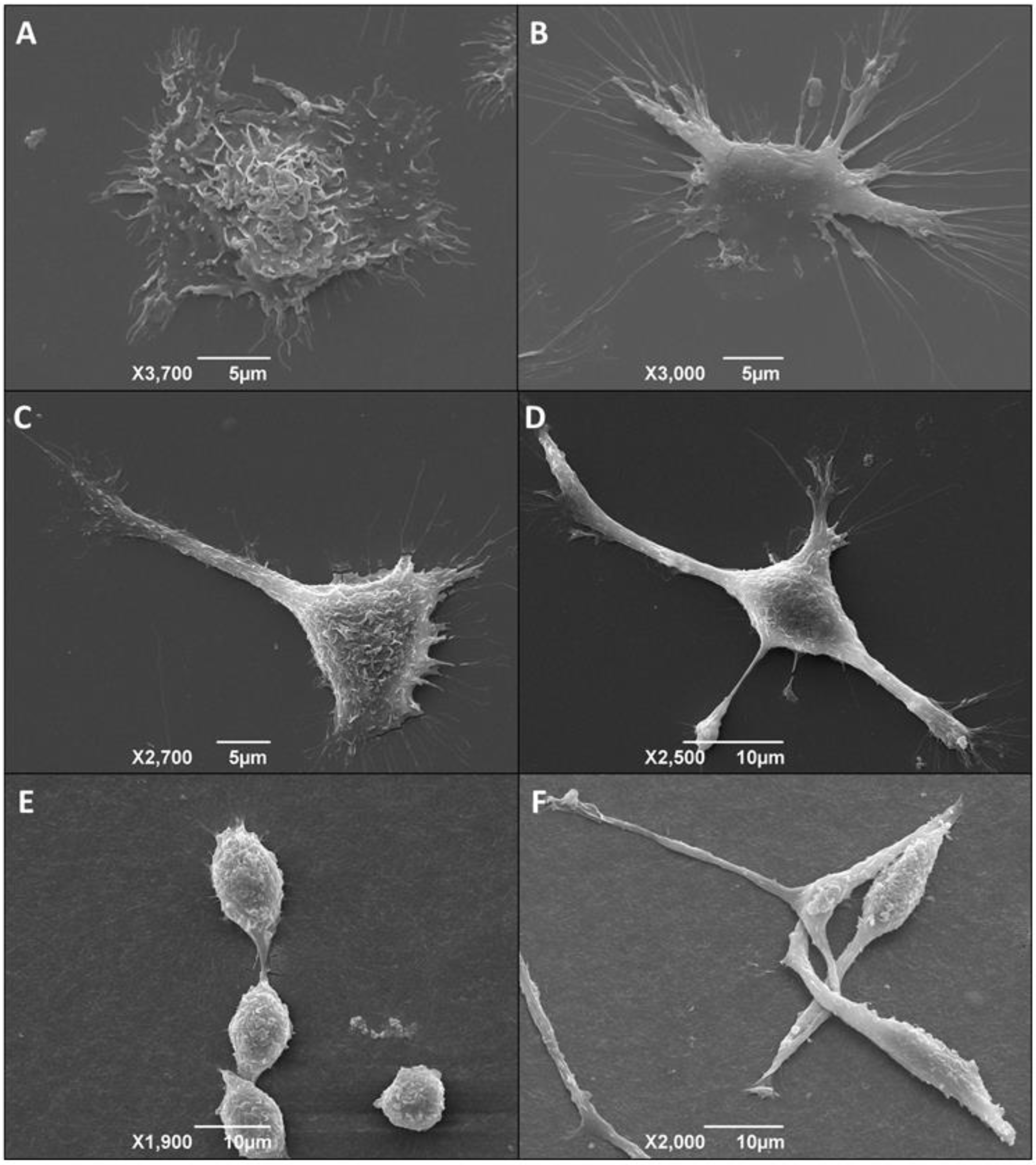
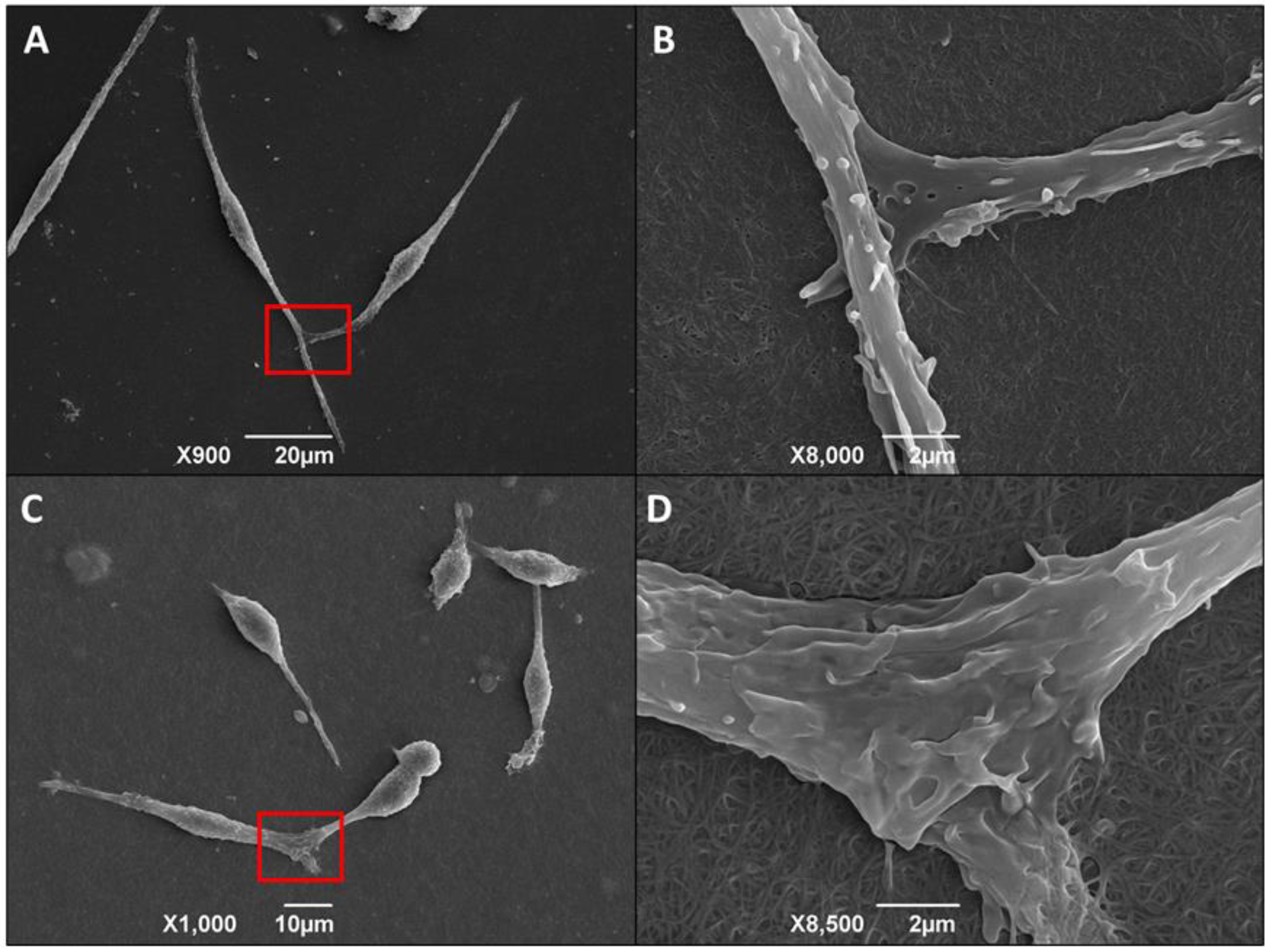
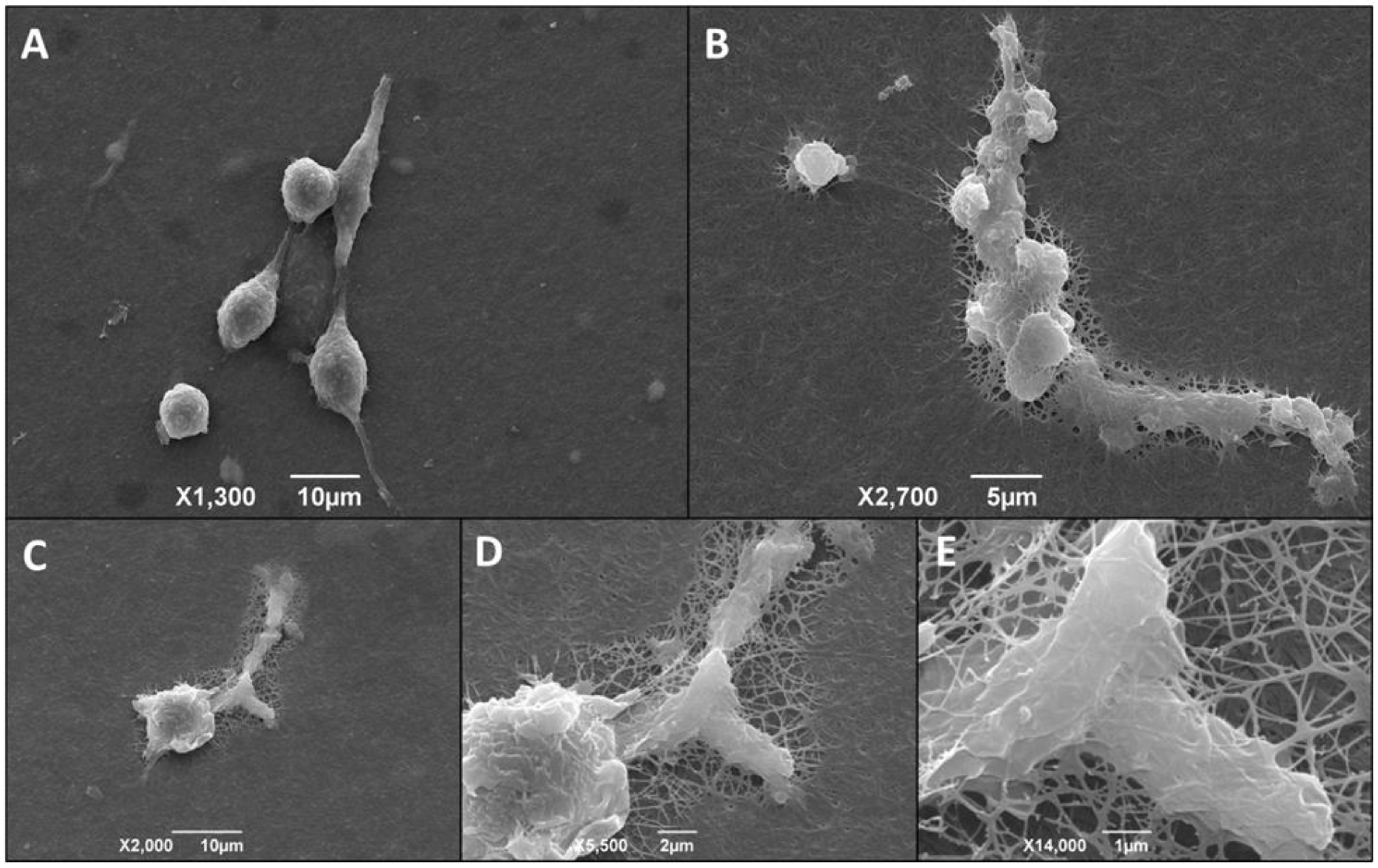
3.6. Morphological Alteration of Infected Macrophages Cultured in 3D Matrix Cultures
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chaudhuri, O.; Cooper-White, J.; Janmey, P.A.; Mooney, D.J.; Shenoy, V.B. Effects of extracellular matrix viscoelasticity on cellular behaviour. Nature 2020, 584, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Bissell, M.J.; Barcellos-Hoff, M.H. The influence of extracellular matrix on gene expression: Is structure the message? J. Cell Sci. Suppl. 1987, 8, 327–343. [Google Scholar] [CrossRef]
- Schenk, S.; Quaranta, V. Tales from the crypt[ic] sites of the extracellular matrix. Trends Cell Biol. 2003, 13, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Nallanthighal, S.; Heiserman, J.P.; Cheon, D.J. The Role of the Extracellular Matrix in Cancer Stemness. Front. Cell Dev. Biol. 2019, 7, 86. [Google Scholar] [CrossRef] [PubMed]
- Halper, J.; Kjaer, M. Basic components of connective tissues and extracellular matrix: Elastin, fibrillin, fibulins, fibrinogen, fibronectin, laminin, tenascins and thrombospondins. Adv. Exp. Med. Biol. 2014, 802, 31–47. [Google Scholar] [CrossRef]
- Gao, X.; Wang, Y.; Chen, J.; Peng, J. The role of peripheral nerve ECM components in the tissue engineering nerve construction. Rev. Neurosci. 2013, 24, 443–453. [Google Scholar] [CrossRef]
- Naomi, R.; Ridzuan, P.M.; Bahari, H. Current Insights into Collagen Type I. Polymers 2021, 13, 2642. [Google Scholar] [CrossRef]
- He, Y.; Liu, W.; Guan, L.; Chen, J.; Duan, L.; Jia, Z.; Huang, J.; Li, W.; Liu, J.; Xiong, J.; et al. A 3D-Printed PLCL Scaffold Coated with Collagen Type I and Its Biocompatibility. Biomed. Res. Int. 2018, 2018, 5147156. [Google Scholar] [CrossRef]
- Idrees, A.; Chiono, V.; Ciardelli, G.; Shah, S.; Viebahn, R.; Zhang, X.; Salber, J. Validation of in vitro assays in three-dimensional human dermal constructs. Int. J. Artif. Organs 2018, 41, 779–788. [Google Scholar] [CrossRef]
- Habanjar, O.; Diab-Assaf, M.; Caldefie-Chezet, F.; Delort, L. 3D Cell Culture Systems: Tumor Application, Advantages, and Disadvantages. Int. J. Mol. Sci. 2021, 22, 12200. [Google Scholar] [CrossRef]
- Chiquet, M.; Matthisson, M.; Koch, M.; Tannheimer, M.; Chiquet-Ehrismann, R. Regulation of extracellular matrix synthesis by mechanical stress. Biochem. Cell Biol. 1996, 74, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.J.; Liu, J.X.; Liu, M.N.; Yao, Y.D.; Liu, Z.Q.; Liu, L.; He, H.H.; Zhou, H. Macrophage 3D migration: A potential therapeutic target for inflammation and deleterious progression in diseases. Pharmacol. Res. 2021, 167, 105563. [Google Scholar] [CrossRef] [PubMed]
- Friedl, P.; Wolf, K.; Lammerding, J. Nuclear mechanics during cell migration. Curr. Opin. Cell Biol. 2011, 23, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A.J.; Schwartz, M.A.; Burridge, K.; Firtel, R.A.; Ginsberg, M.H.; Borisy, G.; Parsons, J.T.; Horwitz, A.R. Cell migration: Integrating signals from front to back. Science 2003, 302, 1704–1709. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Gordon, S.; Pluddemann, A. Tissue macrophages: Heterogeneity and functions. BMC Biol. 2017, 15, 53. [Google Scholar] [CrossRef]
- Freire-de-Lima, C.G.; Xiao, Y.Q.; Gardai, S.J.; Bratton, D.L.; Schiemann, W.P.; Henson, P.M. Apoptotic cells, through transforming growth factor-beta, coordinately induce anti-inflammatory and suppress pro-inflammatory eicosanoid and NO synthesis in murine macrophages. J. Biol. Chem. 2006, 281, 38376–38384. [Google Scholar] [CrossRef]
- Digiacomo, G.; Tusa, I.; Bacci, M.; Cipolleschi, M.G.; Dello Sbarba, P.; Rovida, E. Fibronectin induces macrophage migration through a SFK-FAK/CSF-1R pathway. Cell Adh. Migr. 2017, 11, 327–337. [Google Scholar] [CrossRef]
- Jacob, S.S.; Sudhakaran, P.R. Monocyte-macrophage differentiation in three dimensional collagen lattice. Biochim. Biophys. Acta 2001, 1540, 50–58. [Google Scholar] [CrossRef]
- Zhao, Y.; Zou, W.; Du, J. The origins and homeostasis of monocytes and tissue-resident macrophages in physiological situation. J. Cell Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Faul, C.; Asanuma, K.; Yanagida-Asanuma, E.; Kim, K.; Mundel, P. Actin up: Regulation of podocyte structure and function by components of the actin cytoskeleton. Trends Cell Biol. 2007, 17, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.N.; Martin, A.C. Actin-based force generation and cell adhesion in tissue morphogenesis. Curr. Biol. 2021, 31, R667–R680. [Google Scholar] [CrossRef] [PubMed]
- de Souza, W.; de Carvalho, T.M.; Barrias, E.S. Review on Trypanosoma cruzi: Host Cell Interaction. Int. J. Cell Biol. 2010, 2010, 295394. [Google Scholar] [CrossRef]
- Lopez, M.; Tanowitz, H.B.; Garg, N.J. Pathogenesis of Chronic Chagas Disease: Macrophages, Mitochondria, and Oxidative Stress. Curr. Clin. Microbiol. Rep. 2018, 5, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Song, Z.; Bianco, A. How macrophages respond to two-dimensional materials: A critical overview focusing on toxicity. J. Environ. Sci. Health B 2021, 56, 333–356. [Google Scholar] [CrossRef]
- Hoffmann, E.J.; Ponik, S.M. Biomechanical Contributions to Macrophage Activation in the Tumor Microenvironment. Front. Oncol. 2020, 10, 787. [Google Scholar] [CrossRef]
- Huleihel, L.; Dziki, J.L.; Bartolacci, J.G.; Rausch, T.; Scarritt, M.E.; Cramer, M.C.; Vorobyov, T.; LoPresti, S.T.; Swineheart, I.T.; White, L.J.; et al. Macrophage phenotype in response to ECM bioscaffolds. Semin. Immunol. 2017, 29, 2–13. [Google Scholar] [CrossRef]
- Han, C.; Jin, J.; Xu, S.; Liu, H.; Li, N.; Cao, X. Integrin CD11b negatively regulates TLR-triggered inflammatory responses by activating Syk and promoting degradation of MyD88 and TRIF via Cbl-b. Nat. Immunol. 2010, 11, 734–742. [Google Scholar] [CrossRef]
- Hsieh, J.Y.; Keating, M.T.; Smith, T.D.; Meli, V.S.; Botvinick, E.L.; Liu, W.F. Matrix crosslinking enhances macrophage adhesion, migration, and inflammatory activation. APL Bioeng. 2019, 3, 016103. [Google Scholar] [CrossRef]
- Goldman, R.; Harvey, J.; Hogg, N. VLA-2 is the integrin used as a collagen receptor by leukocytes. Eur. J. Immunol. 1992, 22, 1109–1114. [Google Scholar] [CrossRef]
- Calderwood, D.A.; Tuckwell, D.S.; Eble, J.; Kuhn, K.; Humphries, M.J. The integrin alpha1 A-domain is a ligand binding site for collagens and laminin. J. Biol. Chem. 1997, 272, 12311–12317. [Google Scholar] [CrossRef] [PubMed]
- Moretti, L.; Stalfort, J.; Barker, T.H.; Abebayehu, D. The interplay of fibroblasts, the extracellular matrix, and inflammation in scar formation. J. Biol. Chem. 2022, 298, 101530. [Google Scholar] [CrossRef]
- Contreras, V.T.; Salles, J.M.; Thomas, N.; Morel, C.M.; Goldenberg, S. In vitro differentiation of Trypanosoma cruzi under chemically defined conditions. Mol. Biochem. Parasitol. 1985, 16, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Habermehl, J.; Skopinska, J.; Boccafoschi, F.; Sionkowska, A.; Kaczmarek, H.; Laroche, G.; Mantovani, D. Preparation of ready-to-use, stockable and reconstituted collagen. Macromol. Biosci. 2005, 5, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Court, M.; Malier, M.; Millet, A. 3D type I collagen environment leads up to a reassessment of the classification of human macrophage polarizations. Biomaterials 2019, 208, 98–109. [Google Scholar] [CrossRef]
- Kwon, N.S.; Nathan, C.F.; Stuehr, D.J. Reduced biopterin as a cofactor in the generation of nitrogen oxides by murine macrophages. J. Biol. Chem. 1989, 264, 20496–20501. [Google Scholar] [CrossRef]
- Freire-de-Lima, C.G.; Nunes, M.P.; Corte-Real, S.; Soares, M.P.; Previato, J.O.; Mendonca Previato, L.; Dos Reis, J.S. Proapoptotic Activity of a Trypanosoma cruzi Ceramide-Containing Glycolipid Turned on in Host Macrophages by IFN-γ. J. Immunol. 1998, 161, 4909–4916. [Google Scholar] [CrossRef]
- Kanbara, H.; Yanagi, T.; Nakazawa, S.; Fukuma, T. Comparison of virulent trypomastigotes with low-virulent ones derived from a single strain of Trypanosoma cruzi: Infectivity to new fibroblasts from various organs of the mouse. Zentralbl. Bakteriol. Mikrobiol. Hyg. A 1987, 264, 407–413. [Google Scholar] [CrossRef]
- Tomlinson, S.; Vandekerckhove, F.; Frevert, U.; Nussenzweig, V. The induction of Trypanosoma cruzi trypomastigote to amastigote transformation by low pH. Parasitology 1995, 110 Pt 5, 547–554. [Google Scholar] [CrossRef]
- Berbert, L.R.; Gonzalez, F.B.; Villar, S.R.; Vigliano, C.; Lioi, S.; Beloscar, J.; Bottasso, O.A.; Silva-Barbosa, S.D.; Savino, W.; Perez, A.R. Enhanced Migratory Capacity of T Lymphocytes in Severe Chagasic Patients Is Correlated With VLA-4 and TNF-alpha Expression. Front. Cell Infect. Microbiol. 2021, 11, 713150. [Google Scholar] [CrossRef]
- Freire-de-Lima, C.G.; Nascimento, D.O.; Soares, M.B.; Bozza, P.T.; Castro-Faria-Neto, H.C.; de Mello, F.G.; DosReis, G.A.; Lopes, M.F. Uptake of apoptotic cells drives the growth of a pathogenic trypanosome in macrophages. Nature 2000, 403, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Luna-Gomes, T.; Filardy, A.A.; Rocha, J.D.; Decote-Ricardo, D.; LaRocque-de-Freitas, I.F.; Morrot, A.; Bozza, P.T.; Castro-Faria-Neto, H.C.; DosReis, G.A.; Nunes, M.P.; et al. Neutrophils increase or reduce parasite burden in Trypanosoma cruzi-infected macrophages, depending on host strain: Role of neutrophil elastase. PLoS ONE 2014, 9, e90582. [Google Scholar] [CrossRef] [PubMed]
- Arcanjo, A.F.; LaRocque-de-Freitas, I.F.; Rocha, J.D.; Zamith, D.; Costa-da-Silva, A.C.; Nunes, M.P.; Mesquita-Santos, F.P.; Morrot, A.; Filardy, A.A.; Mariano, M.; et al. The PGE2/IL-10 Axis Determines Susceptibility of B-1 Cell-Derived Phagocytes (B-1CDP) to Leishmania major Infection. PLoS ONE 2015, 10, e0124888. [Google Scholar] [CrossRef]
- Decote-Ricardo, D.; Nunes, M.P.; Morrot, A.; Freire-de-Lima, C.G. Implication of Apoptosis for the Pathogenesis of Trypanosoma cruzi Infection. Front. Immunol. 2017, 8, 518. [Google Scholar] [CrossRef]
- Mendonca, P.H.B.; da Rocha, R.; Moraes, J.B.B.; LaRocque-de-Freitas, I.F.; Logullo, J.; Morrot, A.; Nunes, M.P.; Freire-de-Lima, C.G.; Decote-Ricardo, D. Canine Macrophage DH82 Cell Line As a Model to Study Susceptibility to Trypanosoma cruzi Infection. Front. Immunol. 2017, 8, 604. [Google Scholar] [CrossRef] [PubMed]
- Alexandre-Moreira, M.S.; Freire-de-Lima, C.G.; Trindade, M.N.; Castro-Faria-Neto, H.C.; Piuvezam, M.R.; Pecanha, L.M. Cissampelos sympodialis Eichl (Menispermaceae) leaf extract induces interleukin-10-dependent inhibition of Trypanosoma cruzi killing by macrophages. Braz. J. Med. Biol. Res. 2003, 36, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Cukierman, E.; Pankov, R.; Stevens, D.R.; Yamada, K.M. Taking cell-matrix adhesions to the third dimension. Science 2001, 294, 1708–1712. [Google Scholar] [CrossRef]
- Maltman, D.J.; Przyborski, S.A. Developments in three-dimensional cell culture technology aimed at improving the accuracy of in vitro analyses. Biochem. Soc. Trans. 2010, 38, 1072–1075. [Google Scholar] [CrossRef]
- Petropolis, D.B.; Faust, D.M.; Deep Jhingan, G.; Guillen, N. A new human 3D-liver model unravels the role of galectins in liver infection by the parasite Entamoeba histolytica. PLoS Pathog. 2014, 10, e1004381. [Google Scholar] [CrossRef]
- Petropolis, D.B.; Faust, D.M.; Tolle, M.; Riviere, L.; Valentin, T.; Neuveut, C.; Hernandez-Cuevas, N.; Dufour, A.; Olivo-Marin, J.C.; Guillen, N. Human Liver Infection in a Dish: Easy-To-Build 3D Liver Models for Studying Microbial Infection. PLoS ONE 2016, 11, e0148667. [Google Scholar] [CrossRef]
- Nickerson, C.A.; Richter, E.G.; Ott, C.M. Studying host-pathogen interactions in 3-D: Organotypic models for infectious disease and drug development. J. Neuroimmune Pharmacol. 2007, 2, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Burleigh, B.A.; Andrews, N.W. Signaling and host cell invasion by Trypanosoma cruzi. Curr. Opin. Microbiol. 1998, 1, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Burleigh, B.A. Host cell signaling and Trypanosoma cruzi invasion: Do all roads lead to lysosomes? Sci. STKE 2005, 2005, pe36. [Google Scholar] [CrossRef] [PubMed]
- Hyde, T.P.; Dvorak, J.A. Trypanosoma cruzi: Interaction with vertebrate cells in vitro. 2. Quantitative analysis of the penetration phase. Exp. Parasitol. 1973, 34, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Bonito, V.; Smits, A.; Goor, O.; Ippel, B.D.; Driessen-Mol, A.; Munker, T.; Bosman, A.W.; Mes, T.; Dankers, P.Y.W.; Bouten, C.V.C. Modulation of macrophage phenotype and protein secretion via heparin-IL-4 functionalized supramolecular elastomers. Acta Biomater. 2018, 71, 247–260. [Google Scholar] [CrossRef]
- da Costa Santos, M.A.R.; Dos Reis, J.S.; do Nascimento Santos, C.A.; da Costa, K.M.; Barcelos, P.M.; de Oliveira Francisco, K.Q.; Barbosa, P.; da Silva, E.D.S.; Freire-de-Lima, C.G.; Morrot, A.; et al. Expression of O-glycosylated oncofetal fibronectin in alternatively activated human macrophages. Immunol. Res. 2023, 71, 92–104. [Google Scholar] [CrossRef]
- Calvet, C.M.; Oliveira, F.O., Jr.; Araujo-Jorge, T.C.; Pereira, M.C. Regulation of extracellular matrix expression and distribution in Trypanosoma cruzi-infected cardiomyocytes. Int. J. Med. Microbiol. 2009, 299, 301–312. [Google Scholar] [CrossRef]
- Kajahn, J.; Franz, S.; Rueckert, E.; Forstreuter, I.; Hintze, V.; Moeller, S.; Simon, J.C. Artificial extracellular matrices composed of collagen I and high sulfated hyaluronan modulate monocyte to macrophage differentiation under conditions of sterile inflammation. Biomatter 2012, 2, 226–236. [Google Scholar] [CrossRef]
- Ming, M.; Ewen, M.E.; Pereira, M.E. Trypanosome invasion of mammalian cells requires activation of the TGF beta signaling pathway. Cell 1995, 82, 287–296. [Google Scholar] [CrossRef]
- Waghabi, M.C.; Keramidas, M.; Feige, J.J.; Araujo-Jorge, T.C.; Bailly, S. Activation of transforming growth factor beta by Trypanosoma cruzi. Cell Microbiol. 2005, 7, 511–517. [Google Scholar] [CrossRef]
- Waghabi, M.C.; de Souza, E.M.; de Oliveira, G.M.; Keramidas, M.; Feige, J.J.; Araujo-Jorge, T.C.; Bailly, S. Pharmacological inhibition of transforming growth factor beta signaling decreases infection and prevents heart damage in acute Chagas’ disease. Antimicrob. Agents Chemother. 2009, 53, 4694–4701. [Google Scholar] [CrossRef] [PubMed]
- Cazzulo, J.J.; Couso, R.; Raimondi, A.; Wernstedt, C.; Hellman, U. Further characterization and partial amino acid sequence of a cysteine proteinase from Trypanosoma cruzi. Mol. Biochem. Parasitol. 1989, 33, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.A.; Ferreira, L.F.C.; Pereira, M.C.S.; Calvet, C.M. Differential Role of TGF-beta in Extracellular Matrix Regulation During Trypanosoma cruzi-Host Cell Interaction. Int. J. Mol. Sci. 2019, 20, 4836. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Bejarano, O.H.; Avendano, C.; Patarroyo, M.A. Mechanisms Associated with Trypanosoma cruzi Host Target Cell Adhesion, Recognition and Internalization. Life 2021, 11, 534. [Google Scholar] [CrossRef]
- Hall, B.S.; Pereira, M.A. Dual role for transforming growth factor beta-dependent signaling in Trypanosoma cruzi infection of mammalian cells. Infect. Immun. 2000, 68, 2077–2081. [Google Scholar] [CrossRef]
- Da Rocha, R.F.D.B.; LaRocque-de-Freitas, I.F.; Arcanjo, A.F.; Logullo, J.; Nunes, M.P.; Freire-de-Lima, C.G.; Decote-Ricardo, D. B-1 Cells May Drive Macrophages Susceptibility to Trypanosoma cruzi Infection. Front. Microbiol. 2019, 10, 1598. [Google Scholar] [CrossRef]
- Gutierrez, F.R.; Mineo, T.W.; Pavanelli, W.R.; Guedes, P.M.; Silva, J.S. The effects of nitric oxide on the immune system during Trypanosoma cruzi infection. Memórias Inst. Oswaldo Cruz 2009, 104 (Suppl. 1), 236–245. [Google Scholar] [CrossRef]
- Rebelo, S.P.; Pinto, C.; Martins, T.R.; Harrer, N.; Estrada, M.F.; Loza-Alvarez, P.; Cabecadas, J.; Alves, P.M.; Gualda, E.J.; Sommergruber, W.; et al. 3D-3-culture: A tool to unveil macrophage plasticity in the tumour microenvironment. Biomaterials 2018, 163, 185–197. [Google Scholar] [CrossRef]
- Yoshida, N. Molecular basis of mammalian cell invasion by Trypanosoma cruzi. An. Acad. Bras. Cienc. 2006, 78, 87–111. [Google Scholar] [CrossRef]
- Freire, E.; Coelho-Sampaio, T. Self-assembly of laminin induced by acidic pH. J. Biol. Chem. 2000, 275, 817–822. [Google Scholar] [CrossRef]
- Goncharenko, A.V.; Malyuchenko, N.V.; Moisenovich, A.M.; Kotlyarova, M.S.; Arkhipova, A.Y.; Kon’kov, A.S.; Agapov, I.I.; Molochkov, A.V.; Moisenovich, M.M.; Kirpichnikov, M.P. Changes in morphology of actin filaments and expression of alkaline phosphatase at 3D cultivation of MG-63 osteoblast-like cells on mineralized fibroin scaffolds. Dokl. Biochem. Biophys. 2016, 470, 368–370. [Google Scholar] [CrossRef] [PubMed]
- Cavo, M.; Caria, M.; Pulsoni, I.; Beltrame, F.; Fato, M.; Scaglione, S. A new cell-laden 3D Alginate-Matrigel hydrogel resembles human breast cancer cell malignant morphology, spread and invasion capability observed “in vivo”. Sci. Rep. 2018, 8, 5333. [Google Scholar] [CrossRef]
- Baig, M.S.; Yaqoob, U.; Cao, S.; Saqib, U.; Shah, V.H. Non-canonical role of matrix metalloprotease (MMP) in activation and migration of hepatic stellate cells (HSCs). Life Sci. 2016, 155, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Van Goethem, E.; Poincloux, R.; Gauffre, F.; Maridonneau-Parini, I.; Le Cabec, V. Matrix architecture dictates three-dimensional migration modes of human macrophages: Differential involvement of proteases and podosome-like structures. J. Immunol. 2010, 184, 1049–1061. [Google Scholar] [CrossRef]
- Halper, J. Basic Components of Vascular Connective Tissue and Extracellular Matrix. Adv. Pharmacol. 2018, 81, 95–127. [Google Scholar] [PubMed]
- Sapudom, J.; Waschke, J.; Franke, K.; Hlawitschka, M.; Pompe, T. Quantitative label-free single cell tracking in 3D biomimetic matrices. Sci. Rep. 2017, 7, 14135. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Logullo, J.; Diniz-Lima, I.; Rocha, J.D.B.; Cortê-Real, S.; Silva-Júnior, E.B.d.; Guimarães-de-Oliveira, J.C.; Morrot, A.; Fonseca, L.M.d.; Freire-de-Lima, L.; Decote-Ricardo, D.; et al. Increased Trypanosoma cruzi Growth during Infection of Macrophages Cultured on Collagen I Matrix. Life 2023, 13, 1063. https://doi.org/10.3390/life13041063
Logullo J, Diniz-Lima I, Rocha JDB, Cortê-Real S, Silva-Júnior EBd, Guimarães-de-Oliveira JC, Morrot A, Fonseca LMd, Freire-de-Lima L, Decote-Ricardo D, et al. Increased Trypanosoma cruzi Growth during Infection of Macrophages Cultured on Collagen I Matrix. Life. 2023; 13(4):1063. https://doi.org/10.3390/life13041063
Chicago/Turabian StyleLogullo, Jorgete, Israel Diniz-Lima, Juliana Dutra B. Rocha, Suzana Cortê-Real, Elias Barbosa da Silva-Júnior, Joyce Cristina Guimarães-de-Oliveira, Alexandre Morrot, Leonardo Marques da Fonseca, Leonardo Freire-de-Lima, Debora Decote-Ricardo, and et al. 2023. "Increased Trypanosoma cruzi Growth during Infection of Macrophages Cultured on Collagen I Matrix" Life 13, no. 4: 1063. https://doi.org/10.3390/life13041063