

Fermented Vegetables and Legumes vs. Lifestyle Diseases: Microbiota and More

Abstract
:1. Introduction
2. Methods
3. Probiotic and Prebiotic Potential
3.1. Microbiome and Microbiome-Related Lifestyle Diseases
3.2. Fermented Vegetables and Gut Microbiota
4. Epigenetic Changes
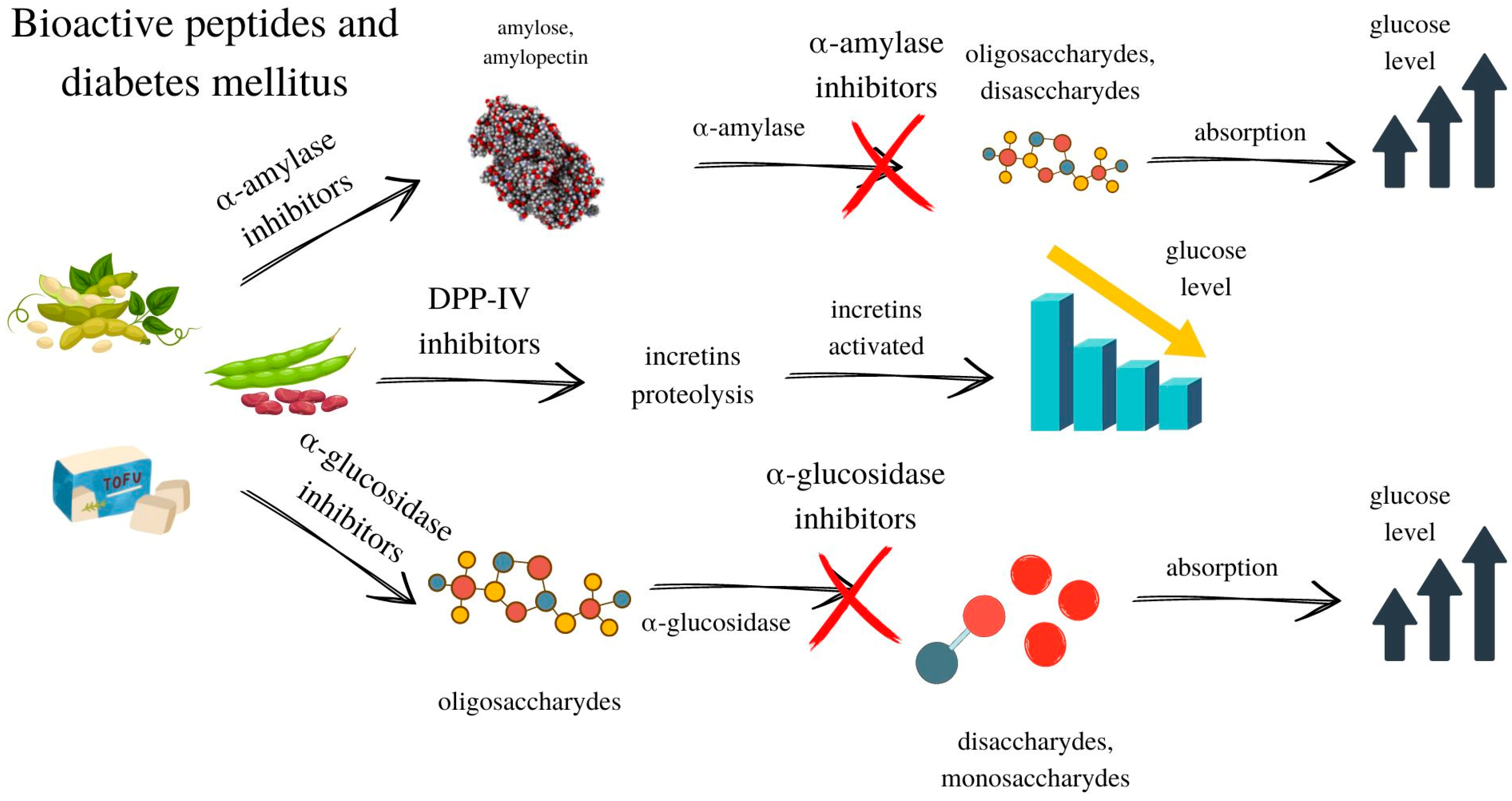
5. Bioactive Peptides
5.1. Inhibitors of Angiotensin I-Converting Enzyme
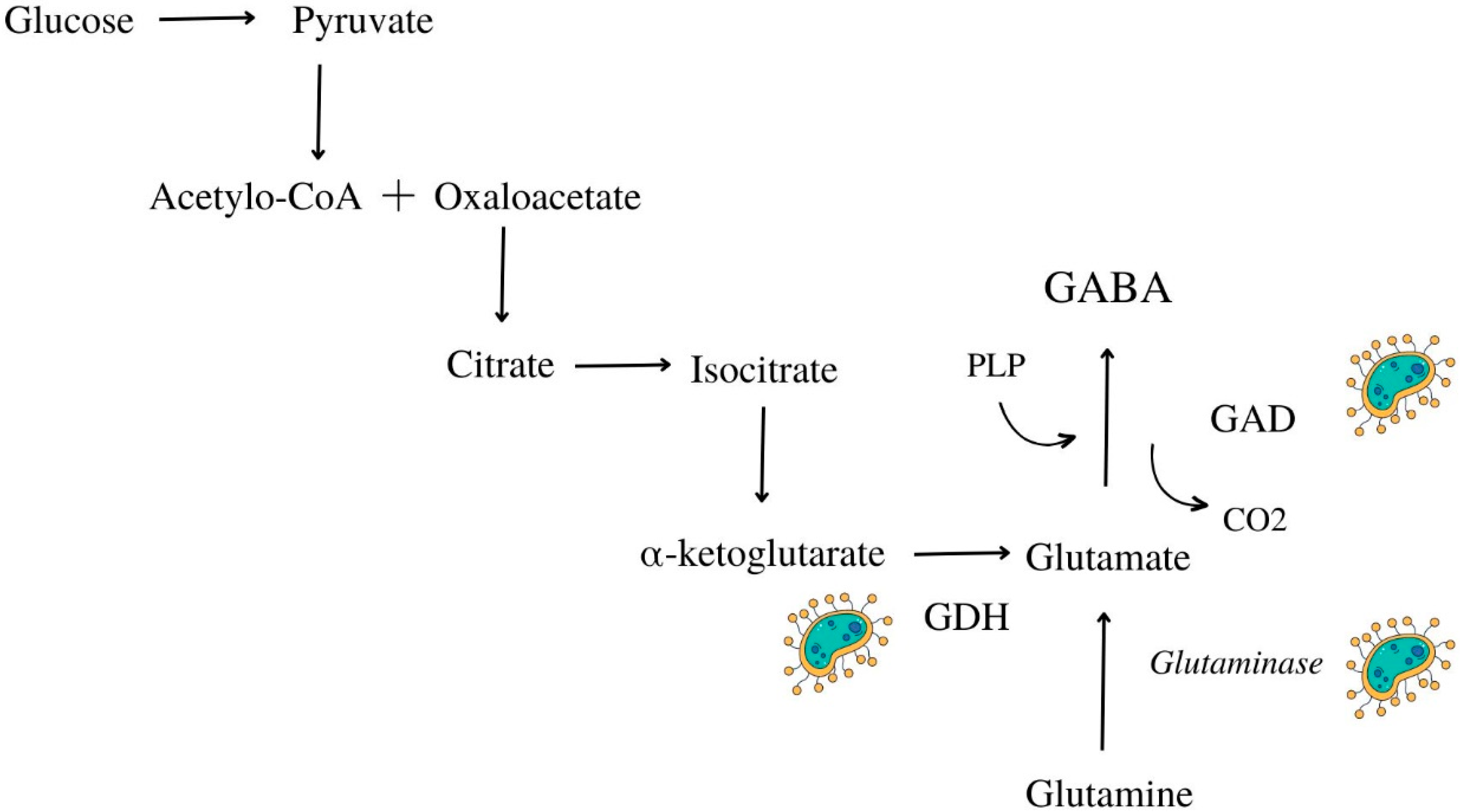
5.2. Γ-Aminobutyric Acid
5.3. Other Food-Derived Bioactive Peptides
6. Antioxidant Potential
Antioxidants Content in Fermented Vegetables

{kind=link}
{kind=link}
| Fermented Vegetable | Used Method | Identified Antioxidant Compound in Fermented Product | Influence of Fermentation on Antioxidant Compounds | Reference |
|---|---|---|---|---|
| Pigeon pea African yam bean Kidney bean | Spectrophotometric method | Polyphenols | Fermentation could increase the free soluble phenolic content and consequently enhance the antioxidant activities | [107] |
| Spinach | Spectrophotometric method | Polyphenols | Fermentation increases the phenolic content | [108] |
| Spinach Broccoli | Spectrophotometric method, LC-MS | Folic acid, 5-methyl tetrahydrofolate | Fermentation increases folic acid content | [109] |
| Chinese cabbage | Spectrophotometric method | Polyphenols, flavonoids | Fermentation increases phenolic content of the methanol extract and reduces flavonoid content in the water extract | [110] |
| Red cabbage | HPLC-DAD-MS/MS | Anthocyanins and derivatives of cyanidin (cyanidin-3-diglucoside-5-glucoside, cyanidin-3-(p-coumaroyl)-diglucoside-5-glucoside, cyanidin-3-(feruloyl)-diglucoside-5-glucoside, cyanidin-3-(sinapoyl)-diglucoside5-glucoside, cyanidin-3-(feruloyl)(feruloyl)-diglucoside-5-glucoside, cyanidin-3-(feruloyl)(sinapoyl)-diglucoside-5-glucoside, and cyanidin-3-(sinapoyl)(sinapoyl)-diglucoside-5-glucoside] | The diminution of anthocyanins content for the fermented product compared to the fresh product | [102] |
| Red cabbage | HPLC-MS/MS | Nonacylated and acylated anthocyanins with the main structure of cyanidin triglucoside | The diminution of anthocyanins content for the fermented product compared to the fresh product | [111] |
| Red beetroot | micro-HPLC-TOF-MS/MS | Betalains (betanin, isobetanin, betanidin and vulgaxanthin) | The fermentation of red beet reduced the content of betalains | [27] |
| Red beetroot | HPLC-MS/MS | Phenolic acids and flavonoids (isoferulic acid, protocatechuic acid, epicatechin, and apigenin | The fermentation process caused an increase in the content of free phenolic acids and reduced the content of conjugated phenolic acids The fermentation process caused a reduction in the content of free flavonoids and an increase in the content of conjugated flavonoids The fermentation process resulted in a reduction in the total content of phenolics (phenolic acids and flavonoids) | [112] |
7. Salt
7.1. Salt and Salt Substitutes vs. Lifestyle Diseases
7.2. Salinity of Fermented Vegetables
7.3. Salt, Fermented Vegetables, and Lifestyle-Diseases
8. Fermented Vegetables and Legumes vs. Lifestyle Diseases
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. The Updated Appendix 3 of the WHO Global NCD Action Plan 2013–2020; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Malesza, I.J.; Malesza, M.; Walkowiak, J.; Mussin, N.; Walkowiak, D.; Aringazina, R.; Bartkowiak-Wieczorek, J.; Mądry, E. High-Fat, Western-Style Diet, Systemic Inflammation, and Gut Microbiota: A Narrative Review. Cells 2021, 10, 3164. [Google Scholar] [CrossRef] [PubMed]
- Shively, C.A.; Appt, S.E.; Vitolins, M.Z.; Uberseder, B.; Michalson, K.T.; Silverstein-Metzler, M.G.; Register, T.C. Mediterranean vs. Western Diet Effects on Caloric Intake, Obesity, Metabolism, and Hepatosteatosis in Nonhuman Primates. Obesity 2019, 27, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Christ, A.; Lauterbach, M.; Latz, E. Western Diet and the Immune System: An Inflammatory Connection. Immunity 2019, 51, 794–811. [Google Scholar] [CrossRef]
- Widmer, R.J.; Flammer, A.J.; Lerman, L.O.; Lerman, A. The Mediterranean Diet, Its Components, and Cardiovascular Disease. Am. J. Med. 2015, 128, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.; Bryan, J.; Hodgson, J.; Murphy, K. Definition of the Mediterranean Diet; a Literature Review. Nutrients 2015, 7, 9139–9153. [Google Scholar] [CrossRef] [PubMed]
- Gille, D.; Schmid, A.; Walther, B.; Vergères, G. Fermented Food and Non-Communicable Chronic Diseases: A Review. Nutrients 2018, 10, 448. [Google Scholar] [CrossRef] [PubMed]
- Şanlier, N.; Gökcen, B.B.; Sezgin, A.C. Health Benefits of Fermented Foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef]
- Mathur, H.; Beresford, T.P.; Cotter, P.D. Health Benefits of Lactic Acid Bacteria (LAB) Fermentates. Nutrients 2020, 12, 1679. [Google Scholar] [CrossRef]
- Chen, X.; Lu, Y.; Zhao, A.; Wu, Y.; Zhang, Y.; Yang, X. Quantitative Analyses for Several Nutrients and Volatile Components during Fermentation of Soybean by Bacillus Subtilis Natto. Food Chem. 2022, 374, 131725. [Google Scholar] [CrossRef]
- Ahnan-Winarno, A.D.; Cordeiro, L.; Winarno, F.G.; Gibbons, J.; Xiao, H. Tempeh: A Semicentennial Review on Its Health Benefits, Fermentation, Safety, Processing, Sustainability, and Affordability. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1717–1767. [Google Scholar] [CrossRef]
- Dey, A.; Rasane, P.; Kaur, S.; Singh, J.; Luwang, M.D. Tofu: Technological and Nutritional Potential. Tech. Artic. 2017, 36, 8–24. [Google Scholar]
- Devanthi, P.V.P.; Gkatzionis, K. Soy Sauce Fermentation: Microorganisms, Aroma Formation, and Process Modification. Food Res. Int. 2019, 120, 364–374. [Google Scholar] [CrossRef]
- Liu, L.; Chen, X.; Hao, L.; Zhang, G.; Jin, Z.; Li, C.; Yang, Y.; Rao, J.; Chen, B. Traditional Fermented Soybean Products: Processing, Flavor Formation, Nutritional and Biological Activities. Crit. Rev. Food Sci. Nutr. 2020, 62, 1971–1989. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, Y.B.; Lee, C.H.; Park, I. Effects of the Addition of Herbs on the Properties of Doenjang. Foods 2021, 10, 1307. [Google Scholar] [CrossRef]
- Yang, H.J.; Kim, H.J.; Kim, M.J.; Kang, S.; Kim, D.S.; Daily, J.W.; Jeong, D.Y.; Kwon, D.Y.; Park, S. Standardized Chungkookjang, Short-Term Fermented Soybeans with Bacillus Lichemiformis, Improves Glucose Homeostasis as Much as Traditionally Made Chungkookjang in Diabetic Rats. J. Clin. Biochem. Nutr. 2013, 52, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Verni, M.; De Mastro, G.; De Cillis, F.; Gobbetti, M.; Rizzello, C.G. Lactic Acid Bacteria Fermentation to Exploit the Nutritional Potential of Mediterranean Faba Bean Local Biotypes. Food Res. Int. 2019, 125, 108571. [Google Scholar] [CrossRef]
- Torino, M.I.; Limón, R.I.; Martínez-Villaluenga, C.; Mäkinen, S.; Pihlanto, A.; Vidal-Valverde, C.; Frias, J. Antioxidant and Antihypertensive Properties of Liquid and Solid State Fermented Lentils. Food Chem. 2013, 136, 1030–1037. [Google Scholar] [CrossRef]
- Zhai, Y.; Pérez-Díaz, I.M. Contribution of Leuconostocaceae to CO2-Mediated Bloater Defect in Cucumber Fermentation. Food Microbiol. 2020, 91, 103536. [Google Scholar] [CrossRef]
- Franco, W.; Pérez-Díaz, I.M. Microbial Interactions Associated with Secondary Cucumber Fermentation. J. Appl. Microbiol. 2013, 114, 161–172. [Google Scholar] [CrossRef]
- Jung, S.J.; Kim, M.J.; Chae, S.W. Quality and Functional Characteristics of Kimchi Made with Organically Cultivated Young Chinese Cabbage (Olgari-Baechu). J. Eth. Foods 2016, 3, 150–158. [Google Scholar] [CrossRef]
- Yang, X.; Hu, W.; Xiu, Z.; Jiang, A.; Yang, X.; Sarengaowa; Ji, Y.; Guan, Y.; Feng, K. Comparison of Northeast Sauerkraut Fermentation between Single Lactic Acid Bacteria Strains and Traditional Fermentation. Food Res. Int. 2020, 137, 109553. [Google Scholar] [CrossRef] [PubMed]
- Šalić, A.; Šamec, D. Changes in the Content of Glucosinolates, Polyphenols and Carotenoids during Lactic-Acid Fermentation of Cruciferous Vegetables: A Mini Review. Food Chem. 2022, 16, 100457. [Google Scholar] [CrossRef] [PubMed]
- Ricci, A.; Marrella, M.; Hadj Saadoun, J.; Bernini, V.; Godani, F.; Dameno, F.; Neviani, E.; Lazzi, C. Development of Lactic Acid-Fermented Tomato Products. Microorganisms 2020, 8, 1192. [Google Scholar] [CrossRef] [PubMed]
- Aryee, A.N.A.; Owusu-Kwarteng, J.; Senwo, Z.; Alvarez, M.N. Characterizing Fermented Habanero Pepper (Capsicum chinense L). Food Chem. Adv. 2022, 1, 100137. [Google Scholar] [CrossRef]
- Gupta, S.; Ravishankar, S. A Comparison of the Antimicrobial Activity of Garlic, Ginger, Carrot, and Turmeric Pastes against Escherichia Coli O157:H7 in Laboratory Buffer and Ground Beef. Foodborne Pathog. Dis. 2005, 2, 330–340. [Google Scholar] [CrossRef]
- Sawicki, T.; Wiczkowski, W. The Effects of Boiling and Fermentation on Betalain Profiles and Antioxidant Capacities of Red Beetroot Products. Food Chem. 2018, 259, 292–303. [Google Scholar] [CrossRef]
- Martel, J.; Chang, S.H.; Ko, Y.F.; Hwang, T.L.; Young, J.D.; Ojcius, D.M. Gut Barrier Disruption and Chronic Disease. Trends Endocrinol. Metab. 2022, 33, 247–265. [Google Scholar] [CrossRef]
- Amabebe, E.; Robert, F.O.; Agbalalah, T.; Orubu, E.S.F. Microbial Dysbiosis-Induced Obesity: Role of Gut Microbiota in Homoeostasis of Energy Metabolism. Br. J. Nutr. 2020, 123, 1127–1137. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef]
- Vazquez-Marroquin, G.; Ochoa-Précoma, R.; Porchia, L.M.; Pérez-Fuentes, R.; Nicolás-Toledo, L.; Rodríguez-Antolín, J.; Gonzalez-Mejia, M.E. The Effect of Microbiome Therapies on Waist Circumference, a Measure of Central Obesity, in Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Acad. Nutr. Diet. 2023. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-Chain Fatty Acids in Control of Body Weight and Insulin Sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Rahat-Rozenbloom, S.; Fernandes, J.; Gloor, G.B.; Wolever, T.M.S. Evidence for Greater Production of Colonic Short-Chain Fatty Acids in Overweight than Lean Humans. Int. J. Obes. 2014, 38, 1525–1531. [Google Scholar] [CrossRef] [PubMed]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in Lean and Overweight Healthy Subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Mithieux, G.; Gautier-Stein, A. Intestinal Glucose Metabolism Revisited. Diabetes Res. Clin. Pract. 2014, 105, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Meex, R.C.R.; Venema, K.; Blaak, E.E. Gut Microbial Metabolites in Obesity, NAFLD and T2DM. Nat. Rev. Endocrinol. 2019, 15, 261–273. [Google Scholar] [CrossRef]
- Barczyńska, R.; Litwin, M.; Sliżewska, K.; Szalecki, M.; Berdowska, A.; Bandurska, K.; Libudzisz, Z.; Kapuśniak, J. Bacterial Microbiota and Fatty Acids in the Faeces of Overweight and Obese Children. Pol. J. Microbiol. 2018, 67, 339–345. [Google Scholar] [CrossRef]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment Dominates over Host Genetics in Shaping Human Gut Microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, J.; Warmbrunn, M.V.; Nieuwdorp, M.; Clément, K. Metabolism and Metabolic Disorders and the Microbiome: The Intestinal Microbiota Associated With Obesity, Lipid Metabolism, and Metabolic Health-Pathophysiology and Therapeutic Strategies. Gastroenterology 2021, 160, 573–599. [Google Scholar] [CrossRef]
- Chen, J.; Vitetta, L. Gut Microbiota Metabolites in NAFLD Pathogenesis and Therapeutic Implications. Int. J. Mol. Sci. 2020, 21, 5214. [Google Scholar] [CrossRef]
- Hajer, G.R.; van Haeften, T.W.; Visseren, F.L.J. Adipose Tissue Dysfunction in Obesity, Diabetes, and Vascular Diseases. Eur. Heart J. 2008, 29, 2959–2971. [Google Scholar] [CrossRef]
- Soderborg, T.K.; Clark, S.E.; Mulligan, C.E.; Janssen, R.C.; Babcock, L.; Ir, D.; Lemas, D.J.; Johnson, L.K.; Weir, T.; Lenz, L.L.; et al. The Gut Microbiota in Infants of Obese Mothers Increases Inflammation and Susceptibility to NAFLD. Nat. Commun. 2018, 9, 4462. [Google Scholar] [CrossRef] [PubMed]
- Marco, M.L.; Sanders, M.E.; Gänzle, M.; Arrieta, M.C.; Cotter, P.D.; de Vuyst, L.; Hill, C.; Holzapfel, W.; Lebeer, S.; Merenstein, D.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on Fermented Foods. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 196. [Google Scholar] [CrossRef] [PubMed]
- FAO; WHO. Probiotics in Food Health and Nutritional Properties and Guidelines for Evaluation; FAO Food and Nutrition Paper; FAO: Rome, Italy; WHO: Geneva, Switzerland, 2006. [Google Scholar]
- Cerk, K.; Aguilera-Gómez, M. Microbiota Analysis for Risk Assessment: Evaluation of Hazardous Dietary Substances and Its Potential Role on the Gut Microbiome Variability and Dysbiosis. EFSA J. 2022, 20, 200404. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, M.K.; Giri, S.K. Probiotic Functional Foods: Survival of Probiotics during Processing and Storage. J. Funct. Foods 2014, 9, 225–241. [Google Scholar] [CrossRef]
- Barengolts, E. Gut Microbiota, Prebiotics, Probiotics, and Synbiotics in Management of Obesity and Prediabetes: Review of Randomized Controlled Trials. Endocr. Pract. 2016, 22, 1224–1234. [Google Scholar] [CrossRef] [PubMed]
- Cassani, L.; Gomez-Zavaglia, A.; Simal-Gandara, J. Technological Strategies Ensuring the Safe Arrival of Beneficial Microorganisms to the Gut: From Food Processing and Storage to Their Passage through the Gastrointestinal Tract. Food Res. Int. 2020, 129, 108852. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, S.H.; Jeon, C.O. Kimchi Microflora: History, Current Status, and Perspectives for Industrial Kimchi Production. Appl. Microbiol. Biotechnol. 2014, 98, 2385–2393. [Google Scholar] [CrossRef]
- Kwak, S.-H.; Cho, Y.-M.; Noh, G.-M.; Om, A.-S. Cancer Preventive Potential of Kimchi Lactic Acid Bacteria (Weissella cibaria, Lactobacillus plantarum). J. Cancer Prev. 2014, 19, 253–258. [Google Scholar] [CrossRef]
- Kim, D.W.; Kim, B.M.; Lee, H.J.; Jang, G.J.; Song, S.H.; Lee, J.I.; Lee, S.B.; Shim, J.M.; Lee, K.W.; Kim, J.H.; et al. Effects of Different Salt Treatments on the Fermentation Metabolites and Bacterial Profiles of Kimchi. J. Food Sci. 2017, 82, 1124–1131. [Google Scholar] [CrossRef]
- Xiong, T.; Li, X.; Guan, Q.; Peng, F.; Xie, M. Starter Culture Fermentation of Chinese Sauerkraut: Growth, Acidification and Metabolic Analyses. Food Control 2014, 41, 122–127. [Google Scholar] [CrossRef]
- Nielsen, E.S.; Garnås, E.; Jensen, K.J.; Hansen, L.H.; Olsen, P.S.; Ritz, C.; Krych, L.; Nielsen, D.S. Lacto-Fermented Sauerkraut Improves Symptoms in IBS Patients Independent of Product Pasteurisation—A Pilot Study. Food Funct. 2018, 9, 5323–5335. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.F.; DuVivier, R.; Johanningsmeier, S.D. Changes in the Free Amino Acid Profile of Pickling Cucumber during Lactic Acid Fermentation. J. Food Sci. 2022, 87, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, N.; Nozaki, N.; Ishihara, K.; Ishikawa, A.; Tsuji, H. Analysis of Isoflavone Content in Tempeh, a Fermented Soybean, and Preparation of a New Isoflavone-Enriched Tempeh. J. Biosci. Bioeng. 2005, 100, 685–687. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.; Li, J.; Liang, F.; Wang, Y.; Guan, Q.; Xie, M. Effects of Salt Concentration on Chinese Sauerkraut Fermentation. LWT—Food Sci. Technol. 2016, 69, 169–174. [Google Scholar] [CrossRef]
- Lorn, D.; Nguyen, T.K.C.; Ho, P.H.; Tan, R.; Licandro, H.; Waché, Y. Screening of Lactic Acid Bacteria for Their Potential Use as Aromatic Starters in Fermented Vegetables. Int. J. Food Microbiol. 2021, 350, 109242. [Google Scholar] [CrossRef]
- Park, K.Y.; Jeong, J.K.; Lee, Y.E.; Daily, J.W. Health Benefits of Kimchi (Korean Fermented Vegetables) as a Probiotic Food. J. Med. Food 2014, 17, 6–20. [Google Scholar] [CrossRef]
- Long, R.T.; Zeng, W.S.; Chen, L.Y.; Guo, J.; Lin, Y.Z.; Huang, Q.S.; Luo, S.Q. Bifidobacterium as an Oral Delivery Carrier of Oxyntomodulin for Obesity Therapy: Inhibitory Effects on Food Intake and Body Weight in Overweight Mice. Int. J. Obesity 2010, 34, 712–719. [Google Scholar] [CrossRef]
- An, H.M.; Park, S.Y.; Lee, D.K.; Kim, J.R.; Kyeong Cha, M.; Lee, S.W.; Lim, H.T.; Kim, K.J.; Ha, N.J. Antiobesity and Lipid-Lowering Effects of Bifidobacterium Spp. in High Fat Diet-Induced Obese Rats. Lipids Health Dis. 2011, 10, 116. [Google Scholar] [CrossRef]
- Chen, J.J.; Wang, R.; Li, X.F.; Wang, R.L. Bifidobacterium Longum Supplementation Improved High-Fat-Fed-Induced Metabolic Syndrome and Promoted Intestinal Reg I Gene Expression. Exp. Biol. Med. 2011, 236, 823–831. [Google Scholar] [CrossRef]
- Salazar, N.; Dewulf, E.M.; Neyrinck, A.M.; Bindels, L.B.; Cani, P.D.; Mahillon, J.; de Vos, W.M.; Thissen, J.P.; Gueimonde, M.; de los Reyes-Gavilán, C.G.; et al. Inulin-Type Fructans Modulate Intestinal Bifidobacterium Species Populations and Decrease Fecal Short-Chain Fatty Acids in Obese Women. Clin. Nutr. 2015, 34, 501–507. [Google Scholar] [CrossRef]
- Han, K.; Bose, S.; Wang, J.H.; Kim, B.S.; Kim, M.J.; Kim, E.J.; Kim, H. Contrasting Effects of Fresh and Fermented Kimchi Consumption on Gut Microbiota Composition and Gene Expression Related to Metabolic Syndrome in Obese Korean Women. Mol. Nutr. Food Res. 2015, 59, 1004–1008. [Google Scholar] [CrossRef] [PubMed]
- Mi, T.; Wang, D.; Yao, S.; Yang, H.; Che, Y.; Wu, C. Effects of Salt Concentration on the Quality and Microbial Diversity of Spontaneously Fermented Radish Paocai. Food Res. Int. 2022, 160, 111622. [Google Scholar] [CrossRef]
- Stoll, D.A.; Müller, A.; Meinhardt, A.K.; Dötsch, A.; Greiner, R.; Kulling, S.E.; Huch, M. Influence of Salt Concentration and Iodized Table Salt on the Microbiota of Fermented Cucumbers. Food Microbiol. 2020, 92, 103552. [Google Scholar] [CrossRef]
- Oyedeji, A.B.; Green, E.; Jeff-Agboola, Y.A.; Olanbiwoninu, A.A.; Areo, E.; Martins, I.E.; El-Imam, A.M.A.; Adebo, O.A. Presence of Pathogenic Microorganisms in Fermented Foods. In Indigenous Fermented Foods for the Tropics, 1st ed.; Adebo, O.A., Chinma, C.E., Obedina, A.O., Soares, A.G., Panda, S.K., Gan, R.-Y., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 519–537. [Google Scholar] [CrossRef]
- Weger, B.D.; Gobet, C.; Yeung, J.; Martin, E.; Jimenez, S.; Betrisey, B.; Foata, F.; Berger, B.; Balvay, A.; Foussier, A.; et al. The Mouse Microbiome Is Required for Sex-Specific Diurnal Rhythms of Gene Expression and Metabolism. Cell Metab. 2019, 29, 362–382. [Google Scholar] [CrossRef] [PubMed]
- Hieu, D.Q.; Hang, B.T.B.; Lokesh, J.; Garigliany, M.M.; Huong, D.T.T.; Yen, D.T.; Liem, P.T.; Tam, B.M.; Hai, D.M.; Son, V.N.; et al. Salinity Significantly Affects Intestinal Microbiota and Gene Expression in Striped Catfish Juveniles. Appl. Microbiol. Biotechnol. 2022, 106, 3245–3264. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; van de Wiele, T.; de Bodt, J.; Marzorati, M.; Gewirtz, A.T. Dietary Emulsifiers Directly Alter Human Microbiota Composition and Gene Expression Ex Vivo Potentiating Intestinal Inflammation. Gut 2017, 66, 1414. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-Sierra, A.; Ramos-Lopez, O.; Riezu-Boj, J.I.; Milagro, F.I.; Martinez, J.A. Diet, Gut Microbiota, and Obesity: Links with Host Genetics and Epigenetics and Potential Applications. Adv. Nutr. 2019, 10, 17–30. [Google Scholar] [CrossRef]
- Karaki, S.I.; Mitsui, R.; Hayashi, H.; Kato, I.; Sugiya, H.; Iwanaga, T.; Furness, J.B.; Kuwahara, A. Short-Chain Fatty Acid Receptor, GPR43, Is Expressed by Enteroendocrine Cells and Mucosal Mast Cells in Rat Intestine. Cell Tissue Res. 2006, 324, 353–360. [Google Scholar] [CrossRef]
- Karaki, S.I.; Tazoe, H.; Hayashi, H.; Kashiwabara, H.; Tooyama, K.; Suzuki, Y.; Kuwahara, A. Expression of the Short-Chain Fatty Acid Receptor, GPR43, in the Human Colon. J. Mol. Histol. 2008, 39, 135–142. [Google Scholar] [CrossRef]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-Chain Fatty Acids Stimulate Glucagon-like Peptide-1 Secretion via the G-Protein-Coupled Receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef]
- Yun, C.L.; Zierath, J.R. AMP-Activated Protein Kinase Signaling in Metabolic Regulation. J. Clin. Investig. 2006, 116, 1776. [Google Scholar] [CrossRef]
- Toldrá, F.; Reig, M.; Aristoy, M.C.; Mora, L. Generation of Bioactive Peptides during Food Processing. Food Chem. 2018, 267, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Chai, K.F.; Voo, A.Y.H.; Chen, W.N. Bioactive Peptides from Food Fermentation: A Comprehensive Review of Their Sources, Bioactivities, Applications, and Future Development. Compr. Rev. Food Sci. Food. Saf. 2020, 19, 3825–3885. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Lönnerdal, B. Bioactive Peptides Derived from Human Milk Proteins: An Update. Curr. Opin. Clin. Nutr. Metab. Care 2020, 23, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, C.; Gleddie, S.; Xiao, C.W. Soybean Bioactive Peptides and Their Functional Properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef]
- Guo, Q.; Chen, P.; Chen, X. Bioactive Peptides Derived from Fermented Foods: Preparation and Biological Activities. J. Funct. Foods 2023, 101, 105422. [Google Scholar] [CrossRef]
- Piepho, R.W. Overview of the Angiotensin-Converting-Enzyme Inhibitors. Am. J. Health Syst. Pharm. 2000, 57 (Suppl. 1), 3–7. [Google Scholar] [CrossRef]
- Taniguchi, M.; Aida, R.; Saito, K.; Ochiai, A.; Takesono, S.; Saitoh, E.; Tanaka, T. Identification and Characterization of Multifunctional Cationic Peptides from Traditional Japanese Fermented Soybean Natto Extracts. J. Biosci. Bioeng. 2019, 127, 472–478. [Google Scholar] [CrossRef]
- Ademiluyi, A.O.; Oboh, G. Angiotensin I-Converting Enzyme Inhibitory Activity and Hypocholesterolemic Effect of Some Fermented Tropical Legumes in Streptozotocin-Induced Diabetic Rats. Int. J. Diabetes Dev. Ctries. 2015, 35, 493–500. [Google Scholar] [CrossRef]
- Xiao, Y.; Sun, M.; Zhang, Q.; Chen, Y.; Miao, J.; Rui, X.; Dong, M. Effects of Cordyceps militaris (L.) Fr. Fermentation on the Nutritional, Physicochemical, Functional Properties and Angiotensin I Converting Enzyme Inhibitory Activity of Red Bean (Phaseolus Angularis [Willd.] W.F. Wight.) Flour. J. Food Sci. Technol. 2018, 55, 1244–1255. [Google Scholar] [CrossRef]
- Usinger, L.; Ibsen, H.; Linneberg, A.; Azizi, M.; Flambard, B.; Jensen, L.T. Human in Vivo Study of the Renin-Angiotensin-Aldosterone System and the Sympathetic Activity after 8 Weeks Daily Intake of Fermented Milk. Clin. Physiol. Funct. Imaging 2010, 30, 162–168. [Google Scholar] [CrossRef]
- Rashmi, D.; Zanan, R.; John, S.; Khandagale, K.; Nadaf, A. γ-Aminobutyric Acid (GABA): Biosynthesis, Role, Commercial Production, and Applications. Stud. Nat. Prod. Chem. 2018, 57, 413–452. [Google Scholar] [CrossRef]
- Li, H.; Cao, Y. Lactic Acid Bacterial Cell Factories for Gamma-Aminobutyric Acid. Amino Acids 2010, 39, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Oketch-Rabah, H.A.; Madden, E.F.; Roe, A.L.; Betz, J.M. United States Pharmacopeia (USP) Safety Review of Gamma-Aminobutyric Acid (GABA). Nutrients 2021, 13, 2742. [Google Scholar] [CrossRef] [PubMed]
- de Bie, T.H.; Balvers, M.G.J.; de Vos, R.C.H.; Witkamp, R.F.; Jongsma, M.A. The Influence of a Tomato Food Matrix on the Bioavailability and Plasma Kinetics of Oral Gamma-Aminobutyric Acid (GABA) and Its Precursor Glutamate in Healthy Men. Food Funct. 2022, 13, 8399–8410. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Dasagrandhi, C.; Park, S.K.; Eom, S.H.; Huh, M.K.; Mok, J.S.; Kim, Y.M. Optimization of Gamma-Aminobutyric Acid Production Using Sea Tangle Extract by Lactic Acid Bacterial Fermentation. LWT 2018, 90, 636–642. [Google Scholar] [CrossRef]
- Limón, R.I.; Peñas, E.; Torino, M.I.; Martínez-Villaluenga, C.; Dueñas, M.; Frias, J. Fermentation Enhances the Content of Bioactive Compounds in Kidney Bean Extracts. Food Chem. 2015, 172, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Shah, N.P. Restoration of GABA Production Machinery in Lactobacillus Brevis by Accessible Carbohydrates, Anaerobiosis and Early Acidification. Food Microbiol. 2018, 69, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.S.; Cha, I.T.; Roh, S.W.; Shin, H.H.; Seo, M.J. Enhanced Production of Gamma-Aminobutyric Acid by Optimizing Culture Conditions of Lactobacillus Brevis HYE1 Isolated from Kimchi, a Korean Fermented Food. J. Microbiol. Biotechnol. 2017, 27, 450–459. [Google Scholar] [CrossRef]
- Yoo, D.; Park, Y. Association between the Intake of Fermented Soy Products and Hypertension Risk in Postmenopausal Women and Men Aged 50 Years or Older: The Korea National Health and Nutrition Examination Survey 2013–2018. Nutrients 2020, 12, 3621. [Google Scholar] [CrossRef]
- Venâncio, T.M.; Oliveira, A.E.A.; Silva, L.B.; Machado, O.L.T.; Fernandes, K.V.S.; Xavier-Filho, J. A Protein with Amino Acid Sequence Homology to Bovine Insulin Is Present in the Legume Vigna Unguiculata (Cowpea). Braz J. Med. Biol. Res. 2003, 36, 1167–1173. [Google Scholar] [CrossRef]
- Kehinde, B.A.; Sharma, P. Recently Isolated Antidiabetic Hydrolysates and Peptides from Multiple Food Sources: A Review. Crit. Rev. Food Sci. Nutr. 2018, 60, 322–340. [Google Scholar] [CrossRef]
- Patil, P.; Mandal, S.; Tomar, S.K.; Anand, S. Food Protein-Derived Bioactive Peptides in Management of Type 2 Diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef]
- Tonucci, L.B.; Olbrich dos Santos, K.M.; Licursi de Oliveira, L.; Rocha Ribeiro, S.M.; Duarte Martino, H.S. Clinical Application of Probiotics in Type 2 Diabetes Mellitus: A Randomized, Double-Blind, Placebo-Controlled Study. Clin. Nutr. 2017, 36, 85–92. [Google Scholar] [CrossRef]
- Zhao, D.; Shah, N.P. Changes in Antioxidant Capacity, Isoflavone Profile, Phenolic and Vitamin Contents in Soymilk during Extended Fermentation. LWT 2014, 58, 454–462. [Google Scholar] [CrossRef]
- Ciska, E.; Honke, J.; Drabińska, N. Changes in Glucosinolates and Their Breakdown Products during the Fermentation of Cabbage and Prolonged Storage of Sauerkraut: Focus on Sauerkraut Juice. Food Chem. 2021, 365, 130498. [Google Scholar] [CrossRef]
- Cai, Y.X.; Wang, J.H.; McAuley, C.; Augustin, M.A.; Terefe, N.S. Fermentation for Enhancing the Bioconversion of Glucoraphanin into Sulforaphane and Improve the Functional Attributes of Broccoli Puree. J. Funct. Foods 2019, 61, 103461. [Google Scholar] [CrossRef]
- Odongo, G.A.; Schlotz, N.; Herz, C.; Hanschen, F.S.; Baldermann, S.; Neugart, S.; Trierweiler, B.; Frommherz, L.; Franz, C.M.A.P.; Ngwene, B.; et al. The Role of Plant Processing for the Cancer Preventive Potential of Ethiopian Kale (Brassica carinata). Food Nutr. Res. 2017, 61, 1271527. [Google Scholar] [CrossRef]
- Wiczkowski, W.; Szawara-Nowak, D.; Topolska, J. Changes in the Content and Composition of Anthocyanins in Red Cabbage and Its Antioxidant Capacity during Fermentation, Storage and Stewing. Food Chem. 2015, 167, 115–123. [Google Scholar] [CrossRef]
- Tlais, A.Z.A.; Lemos Junior, W.J.F.; Filannino, P.; Campanaro, S.; Gobbetti, M.; di Cagno, R. How Microbiome Composition Correlates with Biochemical Changes during Sauerkraut Fermentation: A Focus on Neglected Bacterial Players and Functionalities. Microbiol. Spectr. 2022, 10, e00168-22. [Google Scholar] [CrossRef]
- Xiao, F.; Xu, T.; Lu, B.; Liu, R. Guidelines for Antioxidant Assays for Food Components. Food Front. 2020, 1, 60–69. [Google Scholar] [CrossRef]
- Peñas, E.; Frias, J.; Sidro, B.; Vidal-Valverde, C. Chemical Evaluation and Sensory Quality of Sauerkrauts Obtained by Natural and Induced Fermentations at Different NaCl Levels from Brassica Oleracea Var. Capitata Cv. Bronco Grown in Eastern Spain. Effect of Storage. J. Agric. Food Chem. 2010, 58, 3549–3557. [Google Scholar] [CrossRef] [PubMed]
- Ifesan, B.O.T.; Egbewole, O.O.; Ifesan, B.T. Effect of Fermentation on Nutritional Composition of Selected Commonly Consumed Green Leafy Vegetables in Nigeria. Int. J. Appl. Sci. Biotechnol. 2014, 2, 291–297. [Google Scholar] [CrossRef]
- Oboh, G.; Ademiluyi, A.O.; Akindahunsi, A.A. Changes in Polyphenols Distribution and Antioxidant Activity during Fermentation of Some Underutilized Legumes. Food Sci. Technol. Int. 2009, 15, 41–46. [Google Scholar] [CrossRef]
- Aspiyanto; Susilowati, A.; Iskandar, J.M.; Melanie, H.; Maryati, Y.; Lotulung, P.D. Characteristic of Fermented Spinach (Amaranthus spp.) Polyphenol by Kombucha Culture for Antioxidant Compound. AIP Conf. Proc. 2017, 1803, 020018. [Google Scholar] [CrossRef]
- Melanie, H.; Susilowati, A.; Maryati, Y.; Lotulung, P.D. Fermentation of Spinach (Amaranthus Spp.) and Broccoli (Brassica oleracea L.) Using Kombucha Culture as Natural Source of Folic Acid for Functional Food. IPTEK J. Proceed. Ser. 2017, 39, 269–275. [Google Scholar] [CrossRef]
- Sun, Y.P.; Chou, C.C.; Yu, R.C. Antioxidant Activity of Lactic-Fermented Chinese Cabbage. Food Chem. 2009, 115, 912–917. [Google Scholar] [CrossRef]
- Mane, S.; Bremner, D.H.; Tziboula-Clarke, A.; Lemos, M.A. Effect of Ultrasound on the Extraction of Total Anthocyanins from Purple Majesty Potato. Ultrason. Sonochem. 2015, 27, 509–514. [Google Scholar] [CrossRef]
- Płatosz, N.; Sawicki, T.; Wiczkowski, W. Profile of Phenolic Acids and Flavonoids of Red Beet and Its Fermentation Products. Does Long-Term Consumption of Fermented Beetroot Juice Affect Phenolics Profile in Human Blood Plasma and Urine? Pol. J. Food Nutr. Sci. 2020, 70, 55–65. [Google Scholar] [CrossRef]
- Database—Eurostat. Available online: https://ec.europa.eu/eurostat/data/database (accessed on 25 February 2023).
- Moosavian, S.P.; Haghighatdoost, F.; Surkan, P.J.; Azadbakht, L. Salt and Obesity: A Systematic Review and Meta-Analysis of Observational Studies. Int. J. Food Sci. Nutr. 2017, 68, 265–277. [Google Scholar] [CrossRef]
- Allison, S.J. Metabolism: High Salt Intake as a Driver of Obesity. Nat. Rev. Nephrol. 2018, 14, 285. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; He, F.J.; Macgregor, G.A. High Salt Intake: Independent Risk Factor for Obesity? Hypertension 2015, 66, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Ishimoto, T.; Lanaspa, M.A.; Le, M.P.T.; Garcia, G.E.; Diggle, C.P.; MacLean, P.S.; Jackman, M.R.; Asipu, A.; Roncal-Jimenez, C.A.; Kosugia, T.; et al. Opposing Effects of Fructokinase C and A Isoforms on Fructose-Induced Metabolic Syndrome in Mice. Proc. Natl. Acad. Sci. USA 2012, 109, 4320–4325. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.X.; Arcand, J.A.; Campbell, N.R.C.; Johnson, C.; Malta, D.; Petersen, K.; Rae, S.; Santos, J.A.; Sivakumar, B.; Thout, S.R.; et al. The World Hypertension League Science of Salt: A Regularly Updated Systematic Review of Salt and Health Outcomes Studies (Sept 2019 to Dec 2020). J. Hum. Hypertens. 2022, 36, 1048–1058. [Google Scholar] [CrossRef]
- He, F.J.; Tan, M.; Ma, Y.; MacGregor, G.A. Salt Reduction to Prevent Hypertension and Cardiovascular Disease: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2020, 75, 632–647. [Google Scholar] [CrossRef]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut Microbiota Dysbiosis Contributes to the Development of Hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef]
- Richards, E.M.; Pepine, C.J.; Raizada, M.K.; Kim, S. The Gut, Its Microbiome, and Hypertension. Curr. Hypertens. Rep. 2017, 19, 36. [Google Scholar] [CrossRef]
- Henry, M.E.; Appel, L.J. Potassium-Enriched Salt Substitutes: Benefits, Risks, and a “Trolley Problem” in Public Health. Am. J. Clin. Nutr. 2021, 114, 12–13. [Google Scholar] [CrossRef]
- Greer, R.C.; Marklund, M.; Anderson, C.A.M.; Cobb, L.K.; Dalcin, A.T.; Henry, M.; Appel, L.J. Potassium-Enriched Salt Substitutes as a Means to Lower Blood Pressure: Benefits and Risks. Hypertension 2020, 75, 266–274. [Google Scholar] [CrossRef]
- Dupree, D.E.; Price, R.E.; Burgess, B.A.; Andress, E.L.; Breidt, F. Effects of Sodium Chloride or Calcium Chloride Concentration on the Growth and Survival of Escherichia Coli O157:H7 in Model Vegetable Fermentations. J. Food Prot. 2019, 82, 570–578. [Google Scholar] [CrossRef]
- Guan, Q.; Zheng, W.; Huang, T.; Xiao, Y.; Liu, Z.; Peng, Z.; Gong, D.; Xie, M.; Xiong, T. Comparison of Microbial Communities and Physiochemical Characteristics of Two Traditionally Fermented Vegetables. Food Res. Int. 2020, 128, 108755. [Google Scholar] [CrossRef]
- Agirman, B.; Settanni, L.; Erten, H. Effect of Different Mineral Salt Mixtures and Dough Extraction Procedure on the Physical, Chemical and Microbiological Composition of Şalgam: A Black Carrot Fermented Beverage. Food Chem. 2021, 344, 128618. [Google Scholar] [CrossRef]
- Yang, X.; Hu, W.; Jiang, A.; Xiu, Z.; Ji, Y.; Guan, Y.; Sarengaowa; Yang, X. Effect of Salt Concentration on Quality of Chinese Northeast Sauerkraut Fermented by Leuconostoc Mesenteroides and Lactobacillus Plantarum. Food Biosci. 2019, 30, 100421. [Google Scholar] [CrossRef]
- Müller, A.; Rösch, N.; Cho, G.S.; Meinhardt, A.K.; Kabisch, J.; Habermann, D.; Böhnlein, C.; Brinks, E.; Greiner, R.; Franz, C.M.A.P. Influence of Iodized Table Salt on Fermentation Characteristics and Bacterial Diversity during Sauerkraut Fermentation. Food Microbiol. 2018, 76, 473–480. [Google Scholar] [CrossRef]
- Bidlas, E.; Lambert, R.J.W. Comparing the Antimicrobial Effectiveness of NaCl and KCl with a View to Salt/Sodium Replacement. Int. J. Food Microbiol. 2008, 124, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Beuchat, L.R.; Perkins, L.M.; Nakayama, T. Fermentation and Sensory Characteristics of Kimchi Containing Potassium Chloride as a Partial Replacement for Sodium Chloride. Int. J. Food Microbiol. 1994, 21, 335–340. [Google Scholar] [CrossRef]
- Song, H.J.; Lee, H.-J. Consumption of Kimchi, a Salt Fermented Vegetable, Is Not Associated with Hypertension Prevalence. J. Ethn. Foods 2014, 1, 8–12. [Google Scholar] [CrossRef]
- An, S.Y.; Lee, M.S.; Jeon, J.Y.; Ha, E.S.; Kim, T.H.; Yoon, J.Y.; Ok, C.O.; Lee, H.K.; Hwang, W.S.; Choe, S.J.; et al. Beneficial Effects of Fresh and Fermented Kimchi in Prediabetic Individuals. Ann. Nutr. Metab. 2013, 63, 111–119. [Google Scholar] [CrossRef]
- Fan, X.; Yu, L.; Shi, Z.; Li, C.; Zeng, X.; Wu, Z.; Pan, D. Characterization of a Novel Flavored Yogurt Enriched in γ-Aminobutyric Acid Fermented by Levilactobacillus Brevis CGMCC1.5954. J. Dairy Sci. 2023, 106, 852–867. [Google Scholar] [CrossRef]
- Back, H.I.; Kim, S.R.; Yang, J.A.; Kim, M.G.; Chae, S.W.; Cha, Y.S. Effects of Chungkookjang Supplementation on Obesity and Atherosclerotic Indices in Overweight/Obese Subjects: A 12-Week, Randomized, Double-Blind, Placebo-Controlled Clinical Trial. J. Med. Food 2011, 14, 532–537. [Google Scholar] [CrossRef]
- Byun, M.S.; Yu, O.K.; Cha, Y.S.; Park, T.S. Korean Traditional Chungkookjang Improves Body Composition, Lipid Profiles and Atherogenic Indices in Overweight/Obese Subjects: A Double-Blind, Randomized, Crossover, Placebo-Controlled Clinical Trial. Eur. J. Clin. Nutr. 2016, 70, 1116–1122. [Google Scholar] [CrossRef]
- Cavallini, D.C.U.; Manzoni, M.S.J.; Bedani, R.; Roselino, M.N.; Celiberto, L.S.; Vendramini, R.C.; de Valdez, G.F.; Abdalla, D.S.P.; Pinto, R.A.; Rosetto, D.; et al. Probiotic Soy Product Supplemented with Isoflavones Improves the Lipid Profile of Moderately Hypercholesterolemic Men: A Randomized Controlled Trial. Nutrients 2016, 8, 52. [Google Scholar] [CrossRef]
- Choi, I.H.; Noh, J.S.; Han, J.S.; Kim, H.J.; Han, E.S.; Song, Y.O. Kimchi, a Fermented Vegetable, Improves Serum Lipid Profiles in Healthy Young Adults: Randomized Clinical Trial. J. Med. Food 2013, 16, 223. [Google Scholar] [CrossRef]
- Cha, Y.S.; Park, Y.; Lee, M.; Chae, S.W.; Park, K.; Kim, Y.; Lee, H.S. Doenjang, a Korean Fermented Soy Food, Exerts Antiobesity and Antioxidative Activities in Overweight Subjects with the PPAR-Γ2 C1431T Polymorphism: 12-Week, Double-Blind Randomized Clinical Trial. J. Med. Food 2014, 17, 119–127. [Google Scholar] [CrossRef]


| Food Product | Basic Material | Main Additives Different than Salt and Specific Fermentative Microorganisms | Source |
|---|---|---|---|
| Natto | Soybeans | Bacillus subtilis natto | [10] |
| Tempeh | Soybeans | Rhizopus spp. | [11] |
| Sufu (fermented tofu) | Soybeans | Actinomucor elegans, Mucorracemosus, Mucor sufu, Mucor dispersus, Mucorracemosus, Mucor wutuongkiao, and Aspergillus spp. | [12] |
| Soy sauce | Soybeans | Wheat flour/Aspergillus oryaze, Tetragenococcus halophilus, and Zygosaccharomyces rouxii | [13,14] |
| Doenjang | Soybeans | Bacillus subtilis, Bacillus. licheniformis, Bacillus megaterium, Aspergillus spp., Mucor spp., and Rhizopus spp. | [15] |
| Chungkookjang | Soybeans | Bacillus spp. | [16] |
| Fermented legumes different than soya | Chickpeas, beans, peas, lentils, fava beans | LAB | [17,18] |
| Fermented cucumber | Cucumber | LAB | [19,20] |
| Kimchi | Chinese cabbage | Red pepper, garlic, ginger, leek, and glutinous rice flour/LAB | [21] |
| Sauerkraut | White cabbage | LAB | [22] |
| Fermented Cruciferous vegetables other than cabbage | Broccoli, cauliflower, brussels sprouts, kale | LAB | [23] |
| Fermented solanaceous vegetables | Tomatoes, eggplants, peppers (all kind) | LAB | [24,25] |
| Fermented roots | Carrots, radishes, beetroot, celery root, turnips | LAB | [26,27] |
| Fermented Product | Investigated Group | Effect | Source |
|---|---|---|---|
| Chungkookjang | Women and men with obesity and metabolic syndrome (n = 60), 12 weeks | Improvement in apolipoprotein B, which suggests potential antiatherosclerotic effect Decreased visceral fat (from 8.073 mm2 to 7.167 mm2) Decrease in LDL-C from 115.90 mg/100 mL to 111.21 mg/100 mL | [134] |
| Chungkookjang | Women and men with overweight/obesity (n = 166), 12 weeks | No significant difference in metabolic parameters between placebo and study group | [135] |
| Fermented soy product | Men with overweight and moderately increased cholesterol (n = 34), 12 weeks | Reduction of CVD risk markers in moderately hypercholesterolemic men by lipid profile improvement | [136] |
| Kimchi | Women and men with overweight (n = 21), 8 weeks | Decreased appetite, total energy intake, and body weight (2.1 kg) Decrease in systolic (from 126.5 mm Hg to 119.9 mm Hg) and diastolic (from 78.9 mm Hg to 74.9 mm Hg) blood pressure | [132] |
| Kimchi | Healthy volunteers (n = 100), 7 days | Decrease in systolic (from 120.3 mm Hg to 118.3 mm Hg) and diastolic (from 70.3 mm Hg to 69.3 mm Hg) blood pressure Decrease in TG (from 75.2 mg/dL to 67.7 mg/dL), TC (from 174.4 mg/dL to 165.5 mg/dL), and LDL-C (from 98.5 mg/dL to 91.7 mg/dL) Decrease in HDL-C (from 63.3 mg/dL to 60.7 mg/dL) | [137] |
| Doenjang | Overweight and obese patients (n = 83), 12 weeks | Decreased visceral fat Doenjang exhibited antiobesity and antioxidative effect due to improved PPAR-γ expression The catalase activity was increased | [138] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knez, E.; Kadac-Czapska, K.; Grembecka, M. Fermented Vegetables and Legumes vs. Lifestyle Diseases: Microbiota and More. Life 2023, 13, 1044. https://doi.org/10.3390/life13041044
Knez E, Kadac-Czapska K, Grembecka M. Fermented Vegetables and Legumes vs. Lifestyle Diseases: Microbiota and More. Life. 2023; 13(4):1044. https://doi.org/10.3390/life13041044
Chicago/Turabian StyleKnez, Eliza, Kornelia Kadac-Czapska, and Małgorzata Grembecka. 2023. "Fermented Vegetables and Legumes vs. Lifestyle Diseases: Microbiota and More" Life 13, no. 4: 1044. https://doi.org/10.3390/life13041044