
Impact of Light and Dark Treatment on Phenylpropanoid Pathway Genes, Primary and Secondary Metabolites in Agastache rugosa Transgenic Hairy Root Cultures by Overexpressing Arabidopsis Transcription Factor AtMYB12
 , , , and
, , , and 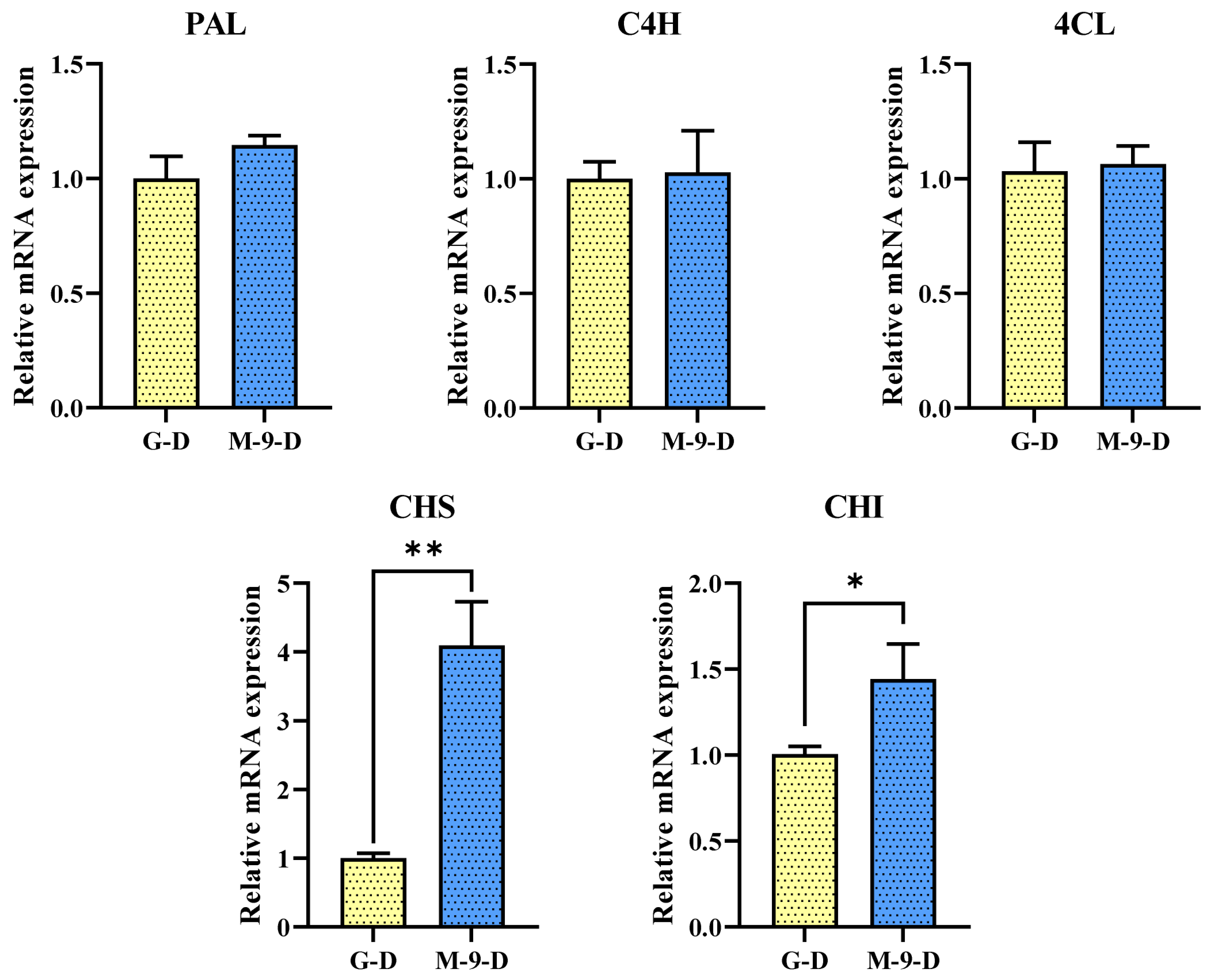{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. AtMYB12-Overexpression Level in Transgenic Hairy Roots
2.2. Total Phenolic Compounds in AtMYB12 Transgenic Hairy Roots
2.3. Impact of Dark Treatment on Phenylpropanoid Pathway Genes Expression in Control and Transgenic Hairy Roots of Korean Mint
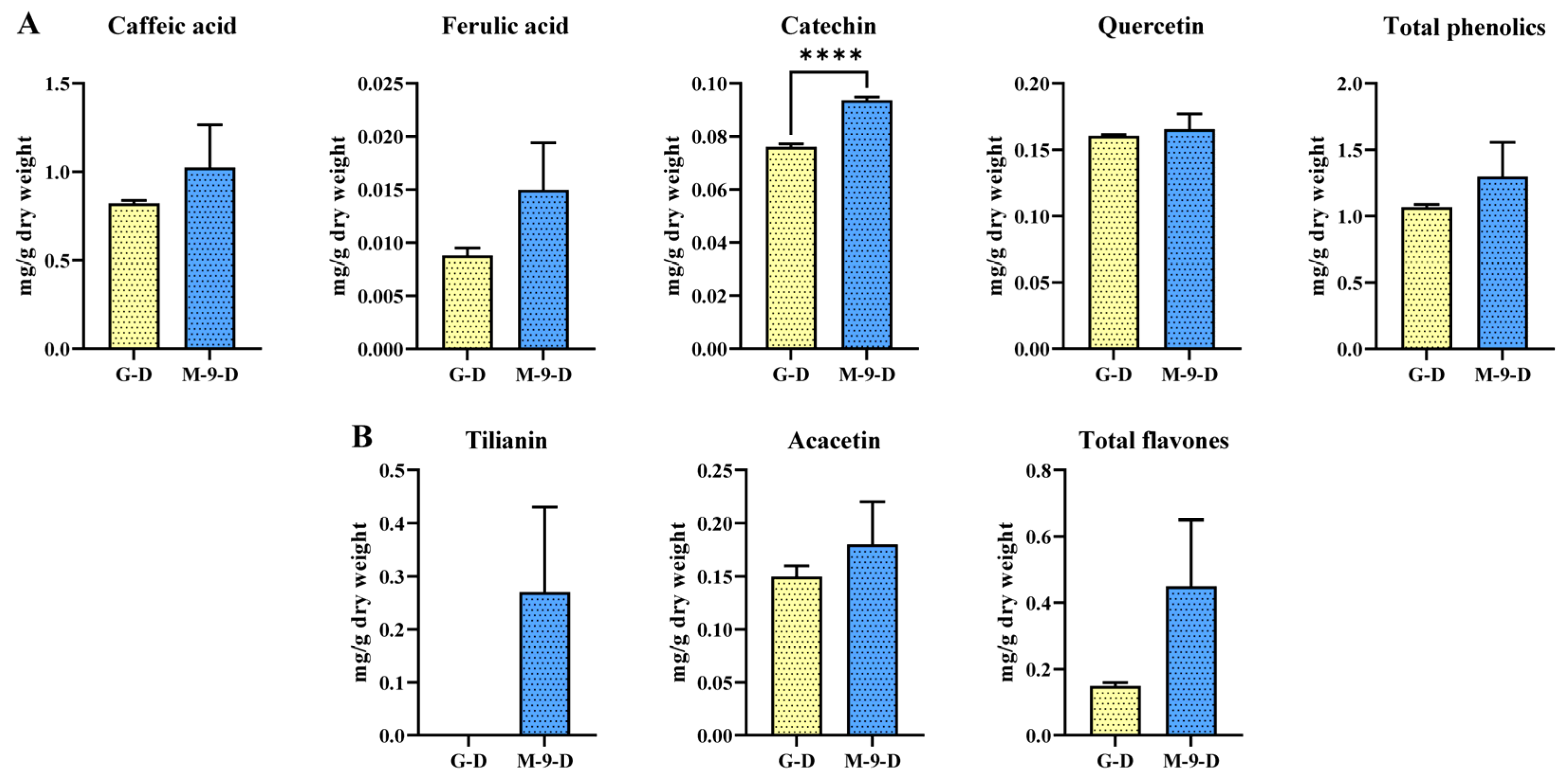
2.4. Impact of Dark Treatment on Phenolic and Flavones Content in Control and Transgenic Hairy Roots of Korean Mint
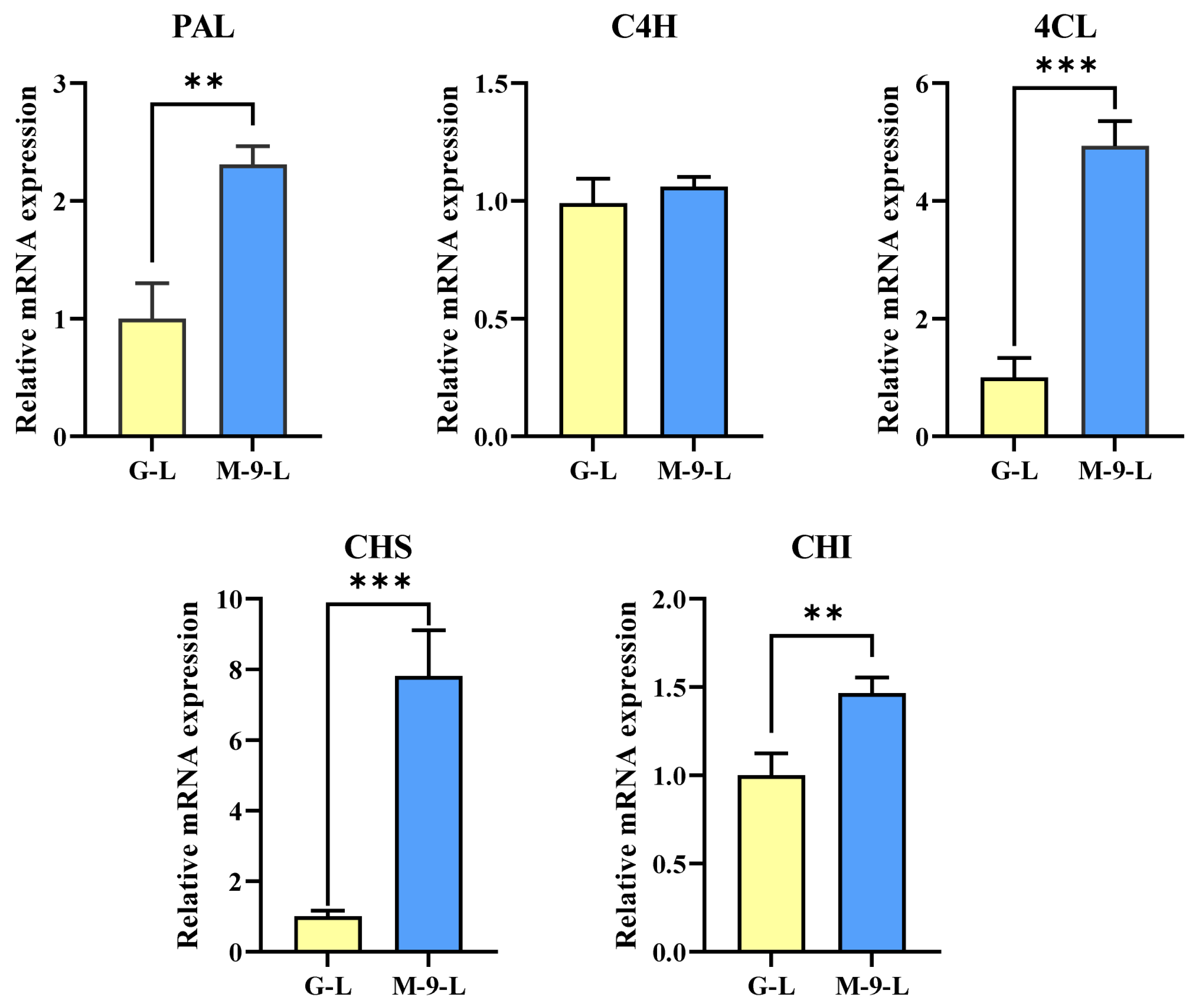
2.5. Impact of Light Treatment on Phenylpropanoid Pathway Genes Expression in Control and Transgenic Hairy Roots of Korean Mint
2.6. Impact of Light Treatment on Phenolic and Flavones Content in Control and Transgenic Hairy Roots of Korean Mint
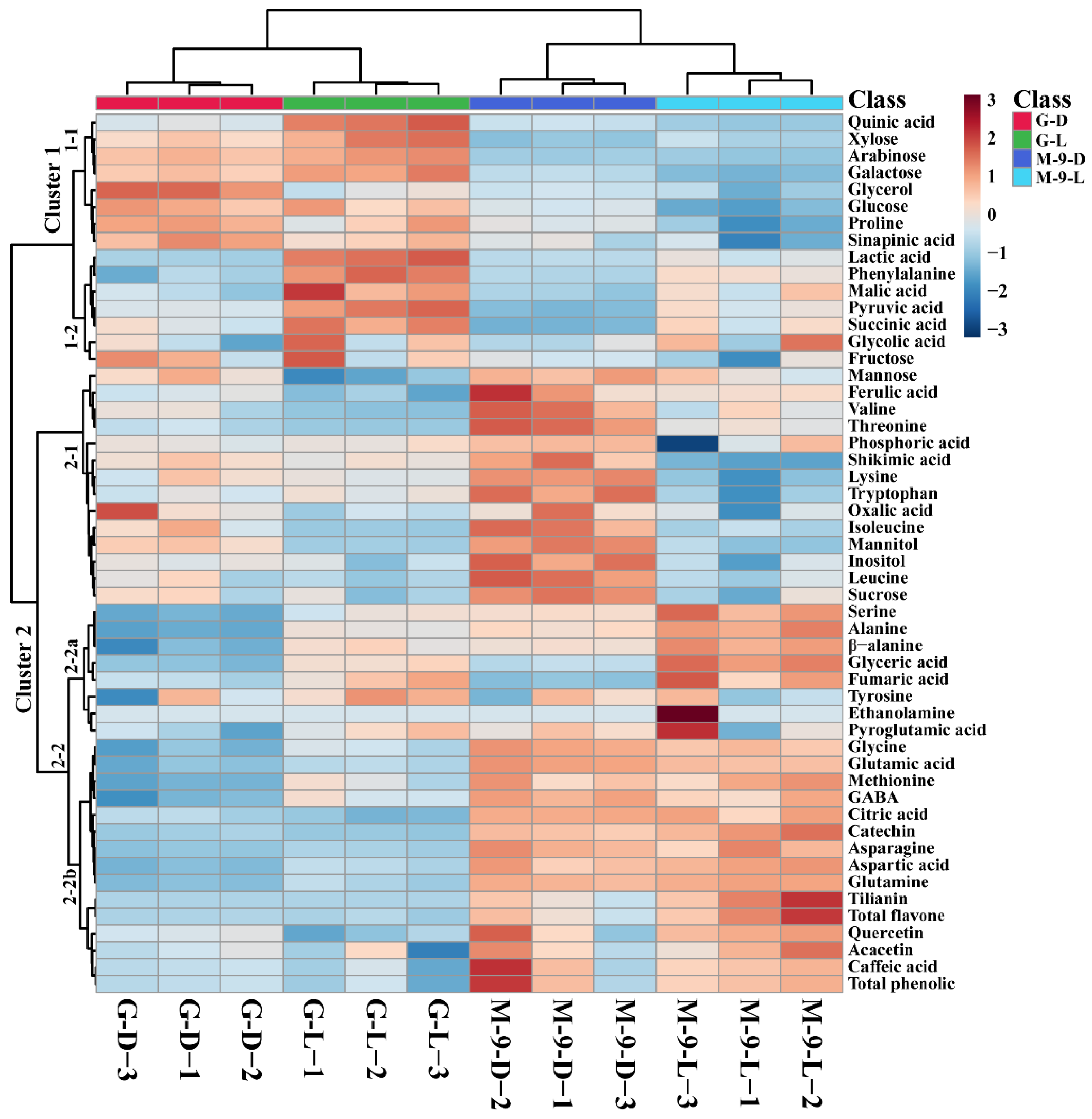
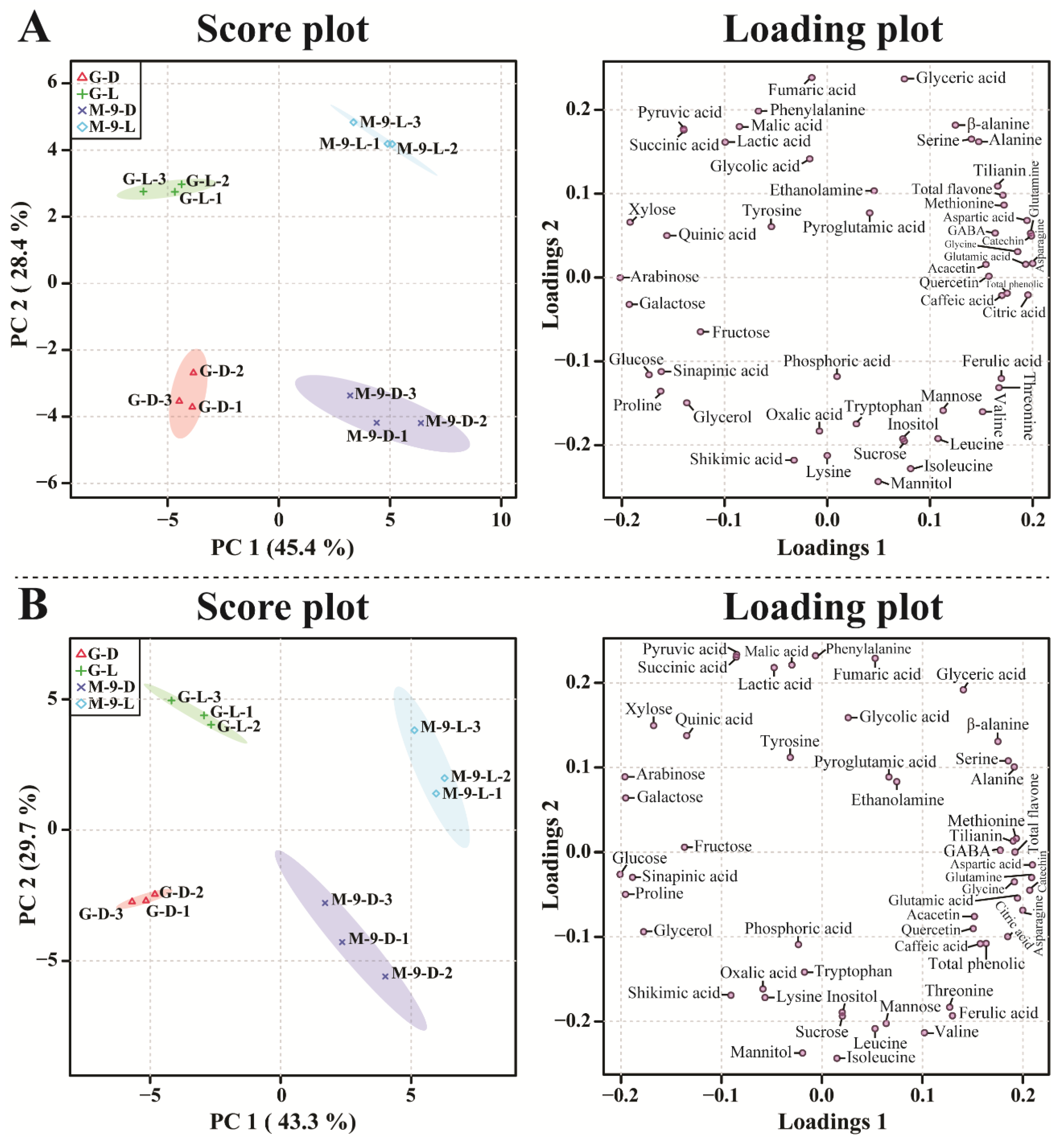
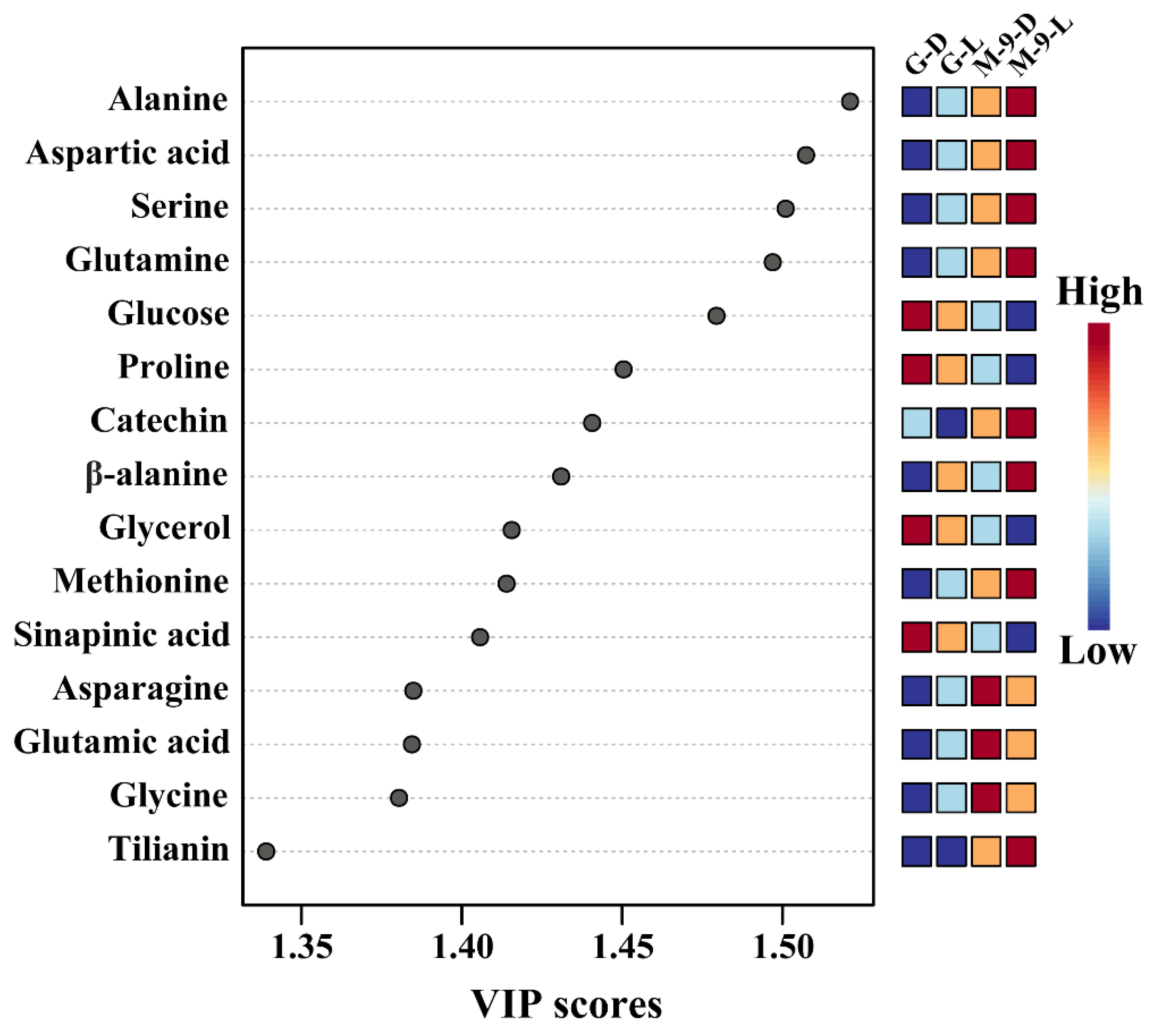
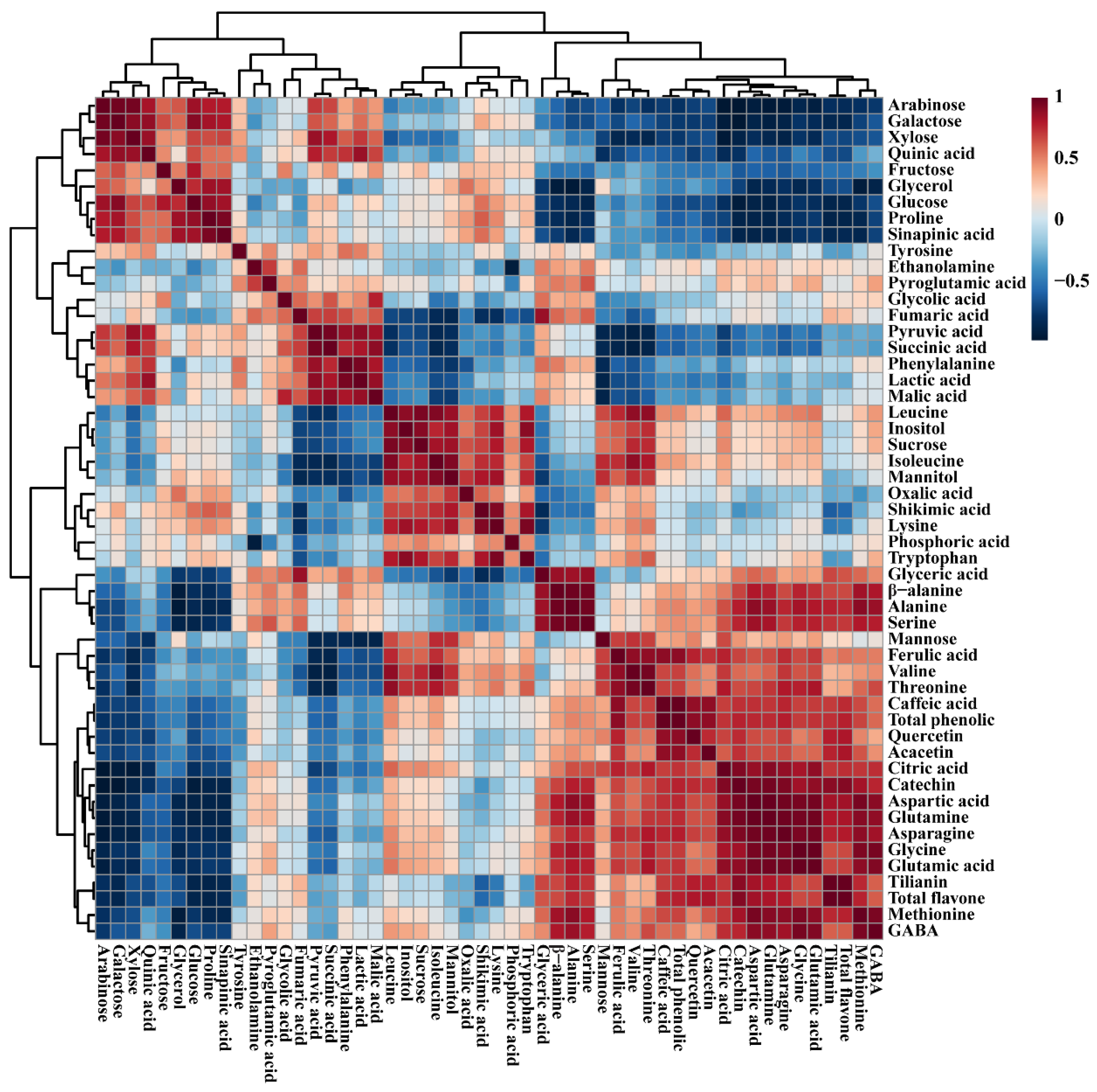
2.7. Metabolic Profiling of Identified Metabolites from Control and Transgenic Hairy Root Lines
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Seed Germination
4.2. Hairy Root Induction
4.3. RNA Extraction and cDNA Synthesis
4.4. Quantitative Real-Time PCR Analysis
4.5. Extraction and HPLC Analysis of Phenolic and Flavones Compounds
4.6. GC-TOFMS Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kumar, S.; Sumner, B.W.; Sumner, L.W. 7.08—Modern plant metabolomics for the discovery and characterization of natural products and their biosynthetic genes. In Comprehensive Natural Products III; Liu, H.-W.B., Begley, T.P., Eds.; Elsevier: Oxford, UK, 2020; pp. 156–188. [Google Scholar]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant Pathol. 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Korkina, L.G. Phenylpropanoids as naturally occurring antioxidants: From plant defense to human health. Cell Mol. Biol. 2007, 53, 15–25. [Google Scholar] [PubMed]
- Carvalho, A.A.; Andrade, L.N.; de Sousa, É.B.V.; de Sousa, D.P. Antitumor phenylpropanoids found in essential oils. BioMed Res. Int. 2015, 2015, 392674. [Google Scholar] [CrossRef] [PubMed]
- Didry, N.; Seidel, V.; Dubreuil, L.; Tillequin, F.; Bailleul, F. Isolation and antibacterial activity of phenylpropanoid derivatives from Ballota nigra. J. Ethnopharmacol. 1999, 67, 197–202. [Google Scholar] [CrossRef]
- Rigano, D.; Formisano, C.; Basile, A.; Lavitola, A.; Senatore, F.; Rosselli, S.; Bruno, M. Antibacterial activity of flavonoids and phenylpropanoids from Marrubium globosum ssp. libanoticum. Phytother. Res. PTR 2007, 21, 395–397. [Google Scholar] [CrossRef]
- Chang, C.-L.; Zhang, L.-J.; Chen, R.Y.; Kuo, L.-M.Y.; Huang, J.-P.; Huang, H.-C.; Lee, K.-H.; Wu, Y.-C.; Kuo, Y.-H. Antioxidant and anti-inflammatory phenylpropanoid derivatives from Calamus quiquesetinervius. J. Nat. Prod. 2010, 73, 1482–1488. [Google Scholar] [CrossRef]
- Piazzon, A.; Vrhovsek, U.; Masuero, D.; Mattivi, F.; Mandoj, F.; Nardini, M. Antioxidant activity of phenolic acids and their metabolites: Synthesis and antioxidant properties of the sulfate derivatives of ferulic and caffeic acids and of the acyl glucuronide of ferulic acid. J. Agric. Food Chem. 2012, 60, 12312–12323. [Google Scholar] [CrossRef]
- Kernan, M.R.; Amarquaye, A.; Chen, J.L.; Chan, J.; Sesin, D.F.; Parkinson, N.; Ye, Z.; Barrett, M.; Bales, C.; Stoddart, C.A.; et al. Antiviral phenylpropanoid glycosides from the medicinal plant Markhamia lutea. J. Nat. Prod. 1998, 61, 564–570. [Google Scholar] [CrossRef]
- Lee, Y.; Lim, H.-W.; Ryu, I.W.; Huang, Y.-H.; Park, M.; Chi, Y.M.; Lim, C.-J. anti-inflammatory, barrier-protective, and antiwrinkle properties of Agastache rugosa Kuntze in human epidermal keratinocytes. BioMed Res. Int. 2020, 2020, 1759067. [Google Scholar] [CrossRef]
- Wróbel-Biedrawa, D.; Grabowska, K.; Galanty, A.; Sobolewska, D.; Podolak, I. A flavonoid on the brain: Quercetin as a potential therapeutic agent in central nervous system disorders. Life 2022, 12, 591. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Baskar, T.B.; Park, Y.E.; Park, J.S.; Lee, S.Y.; Park, S.U. In vitro antioxidant and antimicrobial properties of flower, leaf, and stem extracts of Korean Mint. Antioxidants 2019, 8, 75. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-H.; Kim, M.-J.; Kwon, H.-D.; Park, I.-H. Antimicrobial activity and components of extracts from Agastache rugosa during growth period. Prev. Nutr. Food Sci. 2001, 6, 10–15. [Google Scholar]
- Shin, S.; Kang, C.-A. Antifungal activity of the essential oil of Agastache rugosa Kuntze and its synergism with ketoconazole. Lett. Appl. Microbiol. 2003, 36, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Lee, H.-K.; Shin, C.-G.; Huh, H. HIV integrase inhibitory activity of Agastache rugosa. Arch. Pharm. Res. 1999, 22, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.-H.; Kim, J.S.; Lee, J.; Seo, Y.H.; Kim, H.S.; Ryu, S.M.; Choi, G.; Moon, B.C.; Lee, A.Y. Pharmacological effects of Agastache rugosa against gastritis using a network pharmacology approach. Biomolecules 2020, 10, 1298. [Google Scholar] [CrossRef]
- Jang, S.-A.; Hwang, Y.-H.; Kim, T.; Yang, H.; Lee, J.; Seo, Y.H.; Park, J.-I.; Ha, H. Water extract of Agastache rugosa prevents ovariectomy-induced bone loss by inhibiting osteoclastogenesis. Foods 2020, 9, 1181. [Google Scholar] [CrossRef]
- Zielińska, S.; Matkowski, A. Phytochemistry and bioactivity of aromatic and medicinal plants from the genus Agastache (Lamiaceae). Phytochem. Rev. 2014, 13, 391–416. [Google Scholar] [CrossRef]
- Ambawat, S.; Sharma, P.; Yadav, N.R.; Yadav, R.C. MYB transcription factor genes as regulators for plant responses: An overview. Physiol. Mol. Biol. Plants 2013, 19, 307–321. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, Y.; Wang, Y.; Liu, Z.; Gao, C. Comprehensive analysis of MYB gene family and their expressions under abiotic stresses and hormone treatments in Tamarix hispida. Front. Plant Sci. 2018, 9, 1303. [Google Scholar] [CrossRef]
- Dubos, C.; Le Gourrierec, J.; Baudry, A.; Huep, G.; Lanet, E.; Debeaujon, I.; Routaboul, J.-M.; Alboresi, A.; Weisshaar, B.; Lepiniec, L. MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2008, 55, 940–953. [Google Scholar] [CrossRef]
- Jackson, D.; Culianez-Macia, F.; Prescott, A.G.; Roberts, K.; Martin, C. Expression patterns of myb genes from Antirrhinum flowers. Plant Cell 1991, 3, 115–125. [Google Scholar] [PubMed]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. Cell Mol. Biol. 2008, 53, 814–827. [Google Scholar] [CrossRef]
- Mehrtens, F.; Kranz, H.; Bednarek, P.; Weisshaar, B. The Arabidopsis transcription factor MYB12 is a flavonol-specific regulator of phenylpropanoid biosynthesis. Plant Physiol. 2005, 138, 1083–1096. [Google Scholar] [CrossRef] [PubMed]
- Schilbert, H.M.; Glover, B.J. Analysis of flavonol regulator evolution in the Brassicaceae reveals MYB12, MYB111 and MYB21 duplications associated with MYB11 and MYB24 gene loss. BMC Genom. 2022, 23, 604. [Google Scholar] [CrossRef] [PubMed]
- Stracke, R.; Ishihara, H.; Huep, G.; Barsch, A.; Mehrtens, F.; Niehaus, K.; Weisshaar, B. Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling. Plant J. 2007, 50, 660–677. [Google Scholar] [CrossRef]
- Tian, J.; Zhang, J.; Han, Z.-y.; Song, T.-t.; Li, J.-y.; Wang, Y.-r.; Yao, Y.-c. McMYB12 Transcription factors co-regulate proanthocyanidin and anthocyanin biosynthesis in Malus Crabapple. Sci. Rep. 2017, 7, 43715. [Google Scholar] [CrossRef]
- Wang, N.; Xu, H.; Jiang, S.; Zhang, Z.; Lu, N.; Qiu, H.; Qu, C.; Wang, Y.; Wu, S.; Chen, X. MYB12 and MYB22 play essential roles in proanthocyanidin and flavonol synthesis in red-fleshed apple (Malus sieversii f. niedzwetzkyana). Plant J. 2017, 90, 276–292. [Google Scholar] [CrossRef]
- Shi, M.; Liao, P.; Nile, S.H.; Georgiev, M.I.; Kai, G. Biotechnological exploration of transformed root culture for value-added products. Trends Biotechnol. 2021, 39, 137–149. [Google Scholar] [CrossRef]
- Park, N.I.; Li, X.; Thwe, A.A.; Lee, S.Y.; Kim, S.G.; Wu, Q.; Park, S.U. Enhancement of rutin in Fagopyrum esculentum hairy root cultures by the Arabidopsis transcription factor AtMYB12. Biotechnol. Lett. 2012, 34, 577–583. [Google Scholar] [CrossRef]
- Li, X.H.; Kim, J.K.; Park, S.U. Heterologous expression of three transcription factors differently regulated astragalosides metabolic biosynthesis in Astragalus membranaceus hairy roots. Plants 2022, 11, 1897. [Google Scholar] [CrossRef]
- Wang, F.; Kong, W.; Wong, G.; Fu, L.; Peng, R.; Li, Z.; Yao, Q. AtMYB12 regulates flavonoids accumulation and abiotic stress tolerance in transgenic Arabidopsis thaliana. Mol. Genet. Genom. 2016, 291, 1545–1559. [Google Scholar] [CrossRef] [PubMed]
- Park, W.T.; Yeo, S.K.; Sathasivam, R.; Park, J.S.; Kim, J.K.; Park, S.U. Influence of light-emitting diodes on phenylpropanoid biosynthetic gene expression and phenylpropanoid accumulation in Agastache rugosa. Appl. Biol. Chem. 2020, 63, 25. [Google Scholar] [CrossRef]
- Liu, C.-z.; Guo, C.; Wang, Y.-c.; Ouyang, F. Effect of light irradiation on hairy root growth and artemisinin biosynthesis of Artemisia annua L. Process Biochem. 2002, 38, 581–585. [Google Scholar] [CrossRef]
- Nourozi, E.; Hosseini, B.; Hassani, A. Influences of various factors on hairy root induction in Agastache foeniculum (Pursh) Kuntze. Acta Agric. Slov. 2016, 107, 45. [Google Scholar] [CrossRef]
- Mukherjee, S.; Kutty, N.N.; Bera, P.; Mitra, A. Impact of light and sucrose supplementation on cellular differentiation, metabolic shift and modulation of gene expression in hairy roots of Daucus carota. Plant Cell Tissue Organ Cult. 2019, 136, 383–397. [Google Scholar] [CrossRef]
- Thwe, A.A.; Kim, Y.; Li, X.; Kim, Y.B.; Park, N.-I.; Kim, H.H.; Kim, S.-J.; Park, S.U. Accumulation of phenylpropanoids and correlated gene expression in hairy roots of tartary buckwheat under light and dark conditions. Appl. Biochem. Biotechnol. 2014, 174, 2537–2547. [Google Scholar] [CrossRef]
- Park, C.H.; Park, Y.E.; Yeo, H.J.; Park, N.I.; Park, S.U. Effect of light and dark on the phenolic compound accumulation in tartary buckwheat hairy roots overexpressing ZmLC. Int. J. Mol. Sci. 2021, 22, 4702. [Google Scholar] [CrossRef]
- Zhang, D.; Jiang, C.; Huang, C.; Wen, D.; Lu, J.; Chen, S.; Zhang, T.; Shi, Y.; Xue, J.; Ma, W.; et al. The light-induced transcription factor FtMYB116 promotes accumulation of rutin in Fagopyrum tataricum. Plant Cell Environ. 2019, 42, 1340–1351. [Google Scholar] [CrossRef]
- Pandey, A.; Misra, P.; Bhambhani, S.; Bhatia, C.; Trivedi, P.K. Expression of Arabidopsis MYB transcription factor, AtMYB111, in tobacco requires light to modulate flavonol content. Sci. Rep. 2014, 4, 5018. [Google Scholar] [CrossRef]
- Gutierrez-Valdes, N.; Häkkinen, S.T.; Lemasson, C.; Guillet, M.; Oksman-Caldentey, K.-M.; Ritala, A.; Cardon, F. Hairy Root Cultures—A Versatile Tool with Multiple Applications. Front. Plant Sci. 2020, 11, 33. [Google Scholar] [CrossRef]
- Hu, Z.-B.; Du, M. Hairy root and its application in plant genetic engineering. J. Integr. Plant Biol. 2006, 48, 121–127. [Google Scholar] [CrossRef]
- Du, H.; Zhang, L.; Liu, L.; Tang, X.-F.; Yang, W.-J.; Wu, Y.-M.; Huang, Y.-B.; Tang, Y.-X. Biochemical and molecular characterization of plant MYB transcription factor family. Biochemistry 2009, 74, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Osbourn, A.; Ma, P. MYB transcription factors as regulators of phenylpropanoid metabolism in plants. Mol. Plant 2015, 8, 689–708. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Butelli, E.; Hill, L.; Parr, A.; Niggeweg, R.; Bailey, P.; Weisshaar, B.; Martin, C. AtMYB12 regulates caffeoyl quinic acid and flavonol synthesis in tomato: Expression in fruit results in very high levels of both types of polyphenol. Plant J. 2008, 56, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Misra, P.; Choudhary, D.; Yadav, R.; Goel, R.; Bhambhani, S.; Sanyal, I.; Trivedi, R.; Kumar Trivedi, P. AtMYB12 expression in tomato leads to large scale differential modulation in transcriptome and flavonoid content in leaf and fruit tissues. Sci. Rep. 2015, 5, 12412. [Google Scholar] [CrossRef]
- Li, X.; Sathasivam, R.; Park, N.I.; Wu, Q.; Park, S.U. Enhancement of phenylpropanoid accumulation in tartary buckwheat hairy roots by overexpression of MYB transcription factors. Ind. Crops Prod. 2020, 156, 112887. [Google Scholar] [CrossRef]
- Wang, W.; Hu, S.; Yang, J.; Zhang, C.; Zhang, T.; Wang, D.; Cao, X.; Wang, Z. A novel R2R3-MYB transcription factor SbMYB12 positively regulates baicalin biosynthesis in Scutellaria baicalensis Georgi. Int. J. Mol. Sci. 2022, 23, 15452. [Google Scholar] [CrossRef]
- Yang, L.; Li, W.-C.; Fu, F.-l.; Qu, J.; Sun, F.; Yu, H.; Zhang, J. Characterization of phenylalanine ammonia-lyase genes facilitating flavonoid biosynthesis from two species of medicinal plant Anoectochilus. PeerJ 2022, 10, e13614. [Google Scholar] [CrossRef]
- Singh, K.; Kumar, S.; Rani, A.; Gulati, A.; Ahuja, P.S. Phenylalanine ammonia-lyase (PAL) and cinnamate 4-hydroxylase (C4H) and catechins (flavan-3-ols) accumulation in tea. Funct. Integr. Genom. 2009, 9, 125–134. [Google Scholar] [CrossRef]
- Han, M.; Yang, C.; Zhou, J.; Zhu, J.; Meng, J.; Shen, T.; Xin, Z.; Li, H. Analysis of flavonoids and anthocyanin biosynthesis-related genes expression reveals the mechanism of petal color fading of Malus hupehensis (Rosaceae). Braz. J. Bot. 2020, 43, 81–89. [Google Scholar] [CrossRef]
- Jenkins, G.I.; Long, J.C.; Wade, H.K.; Shenton, M.R.; Bibikova, T.N. UV and blue light signalling: Pathways regulating chalcone synthase gene expression in Arabidopsis. New Phytol. 2001, 151, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Hemm, M.R.; Rider, S.D.; Ogas, J.; Murry, D.J.; Chapple, C. Light induces phenylpropanoid metabolism in Arabidopsis roots. Plant J. 2004, 38, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Cerovic, Z.G.; Pinelli, P.; Tattini, M. Light-induced accumulation of ortho-dihydroxylated flavonoids as non-destructively monitored by chlorophyll fluorescence excitation techniques. Environ. Exp. Bot. 2011, 73, 3–9. [Google Scholar] [CrossRef]
- Shih, C.H.; Chu, H.; Tang, L.K.; Sakamoto, W.; Maekawa, M.; Chu, I.K.; Wang, M.; Lo, C. Functional characterization of key structural genes in rice flavonoid biosynthesis. Planta 2008, 228, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Nam, T.-G.; Lim, Y.J.; Eom, S.H. Flavonoid accumulation in common buckwheat (Fagopyrum esculentum) sprout tissues in response to light. Hortic. Environ. Biotechnol. 2018, 59, 19–27. [Google Scholar] [CrossRef]
- Albert, N.W.; Lewis, D.H.; Zhang, H.; Schwinn, K.E.; Jameson, P.E.; Davies, K.M. Members of an R2R3-MYB transcription factor family in Petunia are developmentally and environmentally regulated to control complex floral and vegetative pigmentation patterning. Plant J. 2011, 65, 771–784. [Google Scholar] [CrossRef]
- Takos, A.M.; Jaffé, F.W.; Jacob, S.R.; Bogs, J.; Robinson, S.P.; Walker, A.R. Light-induced expression of a MYB gene regulates anthocyanin biosynthesis in red apples. Plant Physiol. 2006, 142, 1216–1232. [Google Scholar] [CrossRef]
- Yao, P.; Huang, Y.; Dong, Q.; Wan, M.; Wang, A.; Chen, Y.; Li, C.; Wu, Q.; Chen, H.; Zhao, H. FtMYB6, a light-induced SG7 R2R3-MYB transcription factor, promotes flavonol biosynthesis in tartary buckwheat (Fagopyrum tataricum). J. Agric. Food Chem. 2020, 68, 13685–13696. [Google Scholar] [CrossRef]
- Pandey, A.; Misra, P.; Chandrashekar, K.; Trivedi, P.K. Development of AtMYB12-expressing transgenic tobacco callus culture for production of rutin with biopesticidal potential. Plant Cell Rep. 2012, 31, 1867–1876. [Google Scholar] [CrossRef]
- Pineda-Hidalgo, K.V.; Flores-Leyva, B.; Salazar-Salas, N.Y.; Chávez-Ontiveros, J.; Garzon-Tiznado, J.A.; Sánchez-López, J.; Delgado-Vargas, F.; López-Valenzuela, J.A. Expression of MYB transcription factors and target genes and its association with phenolic content and antioxidant activity of selected Solanum lycopersicum var. cerasiforme accessions from Mexico. CyTA-J. Food 2022, 20, 374–384. [Google Scholar] [CrossRef]
- Stracke, R.; Favory, J.-J.; Gruber, H.; Bartelniewoehner, L.; Bartels, S.; Binkert, M.; Funk, M.; Weisshaar, B.; Ulm, R. The Arabidopsis bZIP transcription factor HY5 regulates expression of the PFG1/MYB12 gene in response to light and ultraviolet-B radiation. Plant Cell Environ. 2010, 33, 88–103. [Google Scholar] [PubMed]
- Xiao, Y.; Chu, L.; Zhang, Y.; Bian, Y.; Xiao, J.; Xu, D. HY5: A pivotal regulator of light-dependent development in higher plants. Front. Plant Sci. 2022, 12, 800989. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Yeo, H.J.; Park, Y.E.; Baek, S.-A.; Kim, J.K.; Park, S.U. Transcriptome analysis and metabolic profiling of Lycoris radiata. Biology 2019, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, E.; Robbertse, P.; Niederwieser, J. Plant carbohydrate partitioning of Lachenalia cv. Ronina during bulb production. Sci. Hortic. 2004, 102, 433–440. [Google Scholar] [CrossRef]
- Smith, C. Carbohydrate biochemistry. In Plant Biochemistry and Molecular Biology, 2nd ed.; Lea, P.J., Leegood, R.C., Eds.; Wiley: Chickester, UK, 1999; pp. 1–384. [Google Scholar]
- Yu, S.; Kwok, K.H.; Doran, P.M. Effect of sucrose, exogenous product concentration, and other culture conditions on growth and steroidal alkaloid production by Solanum aviculare hairy roots. Enzym. Microb. Technol. 1996, 18, 238–243. [Google Scholar] [CrossRef]
- Solfanelli, C.; Poggi, A.; Loreti, E.; Alpi, A.; Perata, P. Sucrose-specific induction of the anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiol. 2006, 140, 637–646. [Google Scholar] [CrossRef]
- Baque, M.A.; Elgirban, A.; Lee, E.-J.; Paek, K.-Y. Sucrose regulated enhanced induction of anthraquinone, phenolics, flavonoids biosynthesis and activities of antioxidant enzymes in adventitious root suspension cultures of Morinda citrifolia (L.). Acta Physiol. Plant 2012, 34, 405–415. [Google Scholar] [CrossRef]
- Weathers, P.; DeJesus-Gonzalez, L.; Kim, Y.; Souret, F.; Towler, M. Alteration of biomass and artemisinin production in Artemisia annua hairy roots by media sterilization method and sugars. Plant Cell Rep. 2004, 23, 414–418. [Google Scholar] [CrossRef]
- Yoon, J.-Y.; Chung, I.-M.; Thiruvengadam, M. Evaluation of phenolic compounds, antioxidant and antimicrobial activities from transgenic hairy root cultures of gherkin (Cucumis anguria L.). S. Afr. J. Bot. 2015, 100, 80–86. [Google Scholar] [CrossRef]
- Ghimire, B.; Thiruvengadam, M.; Chung, I.-M. Identification of elicitors enhances the polyphenolic compounds and pharmacological potential in hairy root cultures of Aster scaber. S. Afr. J. Bot. 2019, 125, 92–101. [Google Scholar] [CrossRef]
- Kim, Y.K.; Li, X.; Xu, H.; Il Park, N.; Uddin, M.R.; Pyon, J.Y.; Park, S.U. Production of phenolic compounds in hairy root culture of tartary buckwheat (Fagopyrum tataricum Gaertn). J. Crop Sci. Biotechnol. 2009, 12, 53–57. [Google Scholar] [CrossRef]
- Sivanandhan, G.; Arun, M.; Mayavan, S.; Rajesh, M.; Mariashibu, T.; Manickavasagam, M.; Selvaraj, N.; Ganapathi, A. Chitosan enhances withanolides production in adventitious root cultures of Withania somnifera (L.) Dunal. Ind. Crops Prod. 2012, 37, 124–129. [Google Scholar] [CrossRef]
- Park, C.H.; Kim, Y.S.; Li, X.; Kim, H.H.; Arasu, M.V.; Al-Dhabi, N.A.; Lee, S.Y.; Park, S.U. Influence of different carbohydrates on flavonoid accumulation in hairy root cultures of Scutellaria baicalensis. Nat. Prod. Commun. 2016, 11, 1934578X1601100625. [Google Scholar] [CrossRef]
- Luo, J.; He, G.-Y. Optimization of elicitors and precursors for paclitaxel production in cell suspension culture of Taxus chinensis in the presence of nutrient feeding. Process Biochem. 2004, 39, 1073–1079. [Google Scholar] [CrossRef]
- Kundu, S.; Salma, U.; Ali, M.N.; Hazra, A.K.; Mandal, N. Development of transgenic hairy roots and augmentation of secondary metabolites by precursor feeding in Sphagneticola calendulacea (L.) Pruski. Ind. Crops Prod. 2018, 121, 206–215. [Google Scholar] [CrossRef]
- Chung, I.-M.; Rekha, K.; Rajakumar, G.; Thiruvengadam, M. Production of glucosinolates, phenolic compounds and associated gene expression profiles of hairy root cultures in turnip (Brassica rapa ssp. rapa). 3 Biotech 2016, 6, 175. [Google Scholar] [CrossRef] [PubMed]
- Weremczuk-Jeżyna, I.; Grzegorczyk-Karolak, I.; Frydrych, B.; Królicka, A.; Wysokińska, H. Hairy roots of Dracocephalum moldavica: Rosmarinic acid content and antioxidant potential. Acta Physiol. Plant 2013, 35, 2095–2103. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Praveen, N.; Maria John, K.; Yang, Y.-S.; Kim, S.-H.; Chung, I.-M. Establishment of Momordica charantia hairy root cultures for the production of phenolic compounds and determination of their biological activities. Plant Cell Tissue Organ Cult. 2014, 118, 545–557. [Google Scholar] [CrossRef]
- Zakhleniuk, O.V.; Raines, C.A.; Lloyd, J.C. pho3: A phosphorus-deficient mutant of Arabidopsis thaliana (L.) Heynh. Planta 2001, 212, 529–534. [Google Scholar] [CrossRef]
- Goyal, S.; Lambert, C.; Cluzet, S.; Mérillon, J.; Ramawat, K.G. Secondary Metabolites and Plant Defence. In Plant Defence: Biological Control; Springer International Publishing: Berlin/Heidelberg, Germany, 2011; pp. 109–138. [Google Scholar]
- Tuan, P.A.; Park, W.T.; Xu, H.; Park, N.I.; Park, S.U. Accumulation of tilianin and rosmarinic acid and expression of phenylpropanoid biosynthetic genes in Agastache rugosa. J. Agric. Food Chem. 2012, 60, 5945–5951. [Google Scholar] [CrossRef]
- Park, C.H.; Sathasivam, R.; Kim, T.J.; Park, B.B.; Kim, J.K.; Park, S.U. Metabolic profiling and secondary metabolite accumulation during fruit development of Cornus officinalis Sieb. et Zucc. Ind. Crops Prod. 2022, 189, 115779. [Google Scholar] [CrossRef]
- Sathasivam, R.; Choi, M.; Radhakrishnan, R.; Kwon, H.; Yoon, J.; Yang, S.H.; Kim, J.K.; Chung, Y.S.; Park, S.U. Effects of various Agrobacterium rhizogenes strains on hairy root induction and analyses of primary and secondary metabolites in Ocimum basilicum. Front. Plant Sci. 2022, 13, 983776. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Do, T.M.H.; Choi, M.; Kim, J.K.; Kim, Y.J.; Park, C.; Park, C.H.; Park, N.I.; Kim, C.; Sathasivam, R.; Park, S.U. Impact of Light and Dark Treatment on Phenylpropanoid Pathway Genes, Primary and Secondary Metabolites in Agastache rugosa Transgenic Hairy Root Cultures by Overexpressing Arabidopsis Transcription Factor AtMYB12. Life 2023, 13, 1042. https://doi.org/10.3390/life13041042
Do TMH, Choi M, Kim JK, Kim YJ, Park C, Park CH, Park NI, Kim C, Sathasivam R, Park SU. Impact of Light and Dark Treatment on Phenylpropanoid Pathway Genes, Primary and Secondary Metabolites in Agastache rugosa Transgenic Hairy Root Cultures by Overexpressing Arabidopsis Transcription Factor AtMYB12. Life. 2023; 13(4):1042. https://doi.org/10.3390/life13041042
Chicago/Turabian StyleDo, Thi Minh Hanh, Minsol Choi, Jae Kwang Kim, Ye Jin Kim, Chanung Park, Chang Ha Park, Nam Il Park, Changsoo Kim, Ramaraj Sathasivam, and Sang Un Park. 2023. "Impact of Light and Dark Treatment on Phenylpropanoid Pathway Genes, Primary and Secondary Metabolites in Agastache rugosa Transgenic Hairy Root Cultures by Overexpressing Arabidopsis Transcription Factor AtMYB12" Life 13, no. 4: 1042. https://doi.org/10.3390/life13041042