
Acai Berry (Euterpe sp.) Extracts Are Neuroprotective against L-Glutamate-Induced Toxicity by Limiting Mitochondrial Dysfunction and Cellular Redox Stress

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture and Treatments
2.2.1. Human Neuroblastoma SH-SY5Y Cell Line Culture
2.2.2. Human Rhabdomyosarcoma Cell Line TE671 Culture
2.2.3. Human Cortical Neuronal Progenitor Cell Culture (ReNcell CX)
2.2.4. Cell Treatments with L-Glutamic Acid (L-Glu) and Aqueous and Ethanolic Extracts of Acai Berries
2.3. (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide (MTT) Cell Viability Assays
2.4. Lactate Dehydrogenase (LDH) Assay
2.5. Adenosine 5′-Triphosphate (ATP) Bioluminescent Assay
2.6. Measurements of Mitochondrial Membrane Potential (MMP) Assay
2.7. 2,7-Dichlorodihydrofluorescein Diacetate (DCFDA) Assay
2.8. Whole-Cell Patch-Clamp Assay
2.9. Liquid-Chromatography Mass-Spectrometry (LC-MS)
2.10. Statistical Analysis
3. Results
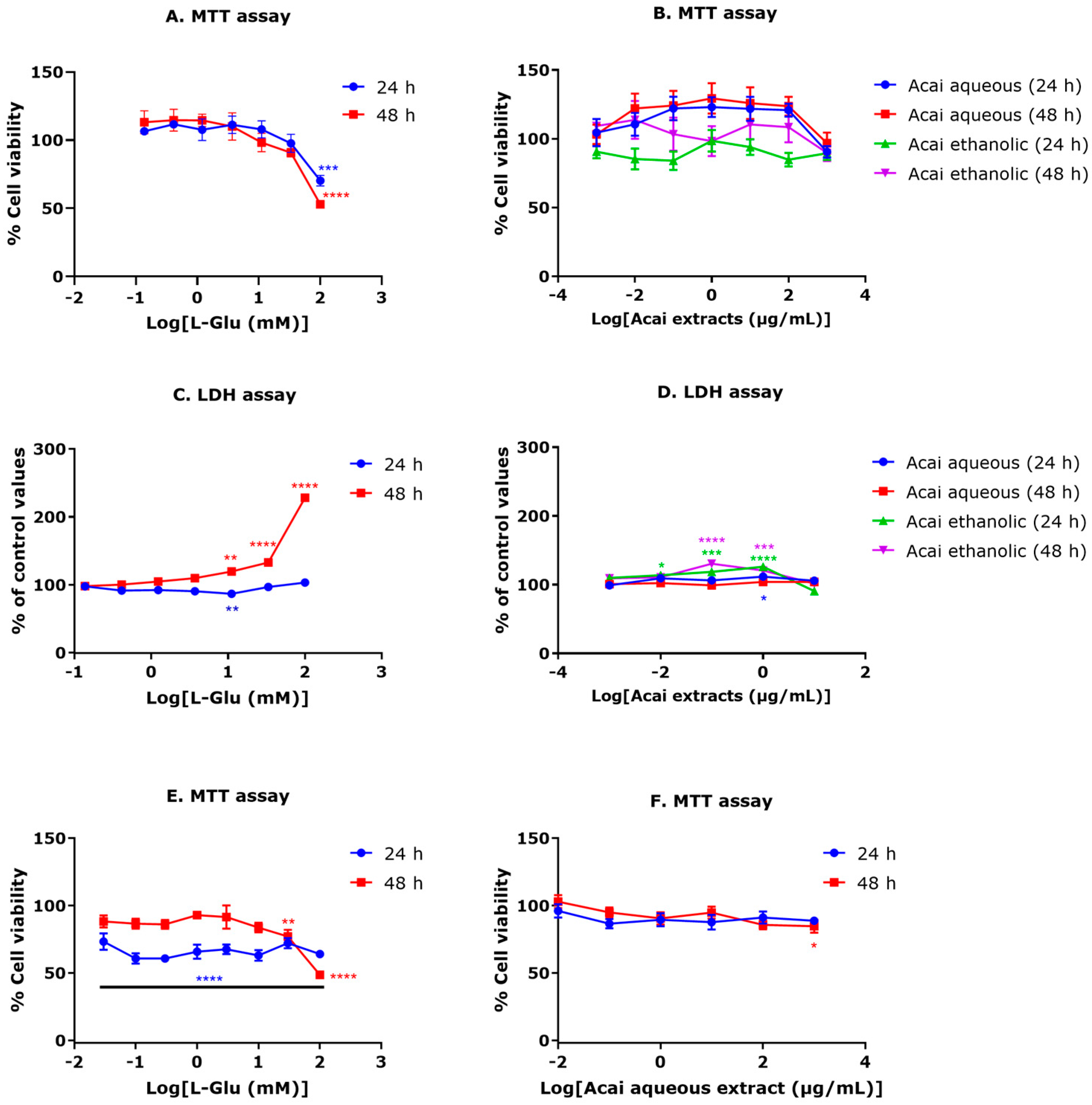
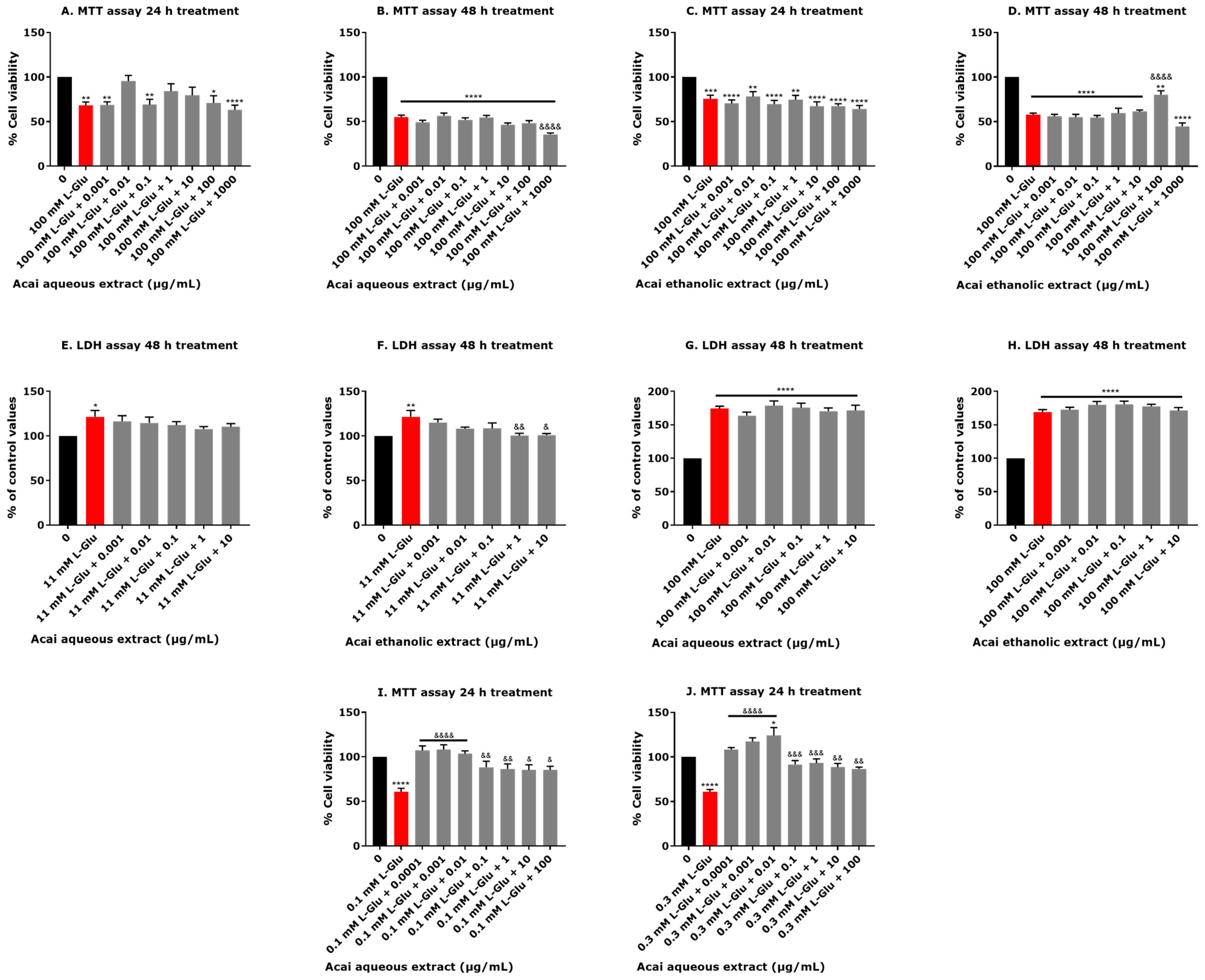
3.1. Exposure of Neuronal Cells to L-Glu Reduces Cell Viability
3.2. Certain Concentrations of Acai Berry Extracts Provide Neuroprotection against L-Glu-Induced Decreased Cell Viability
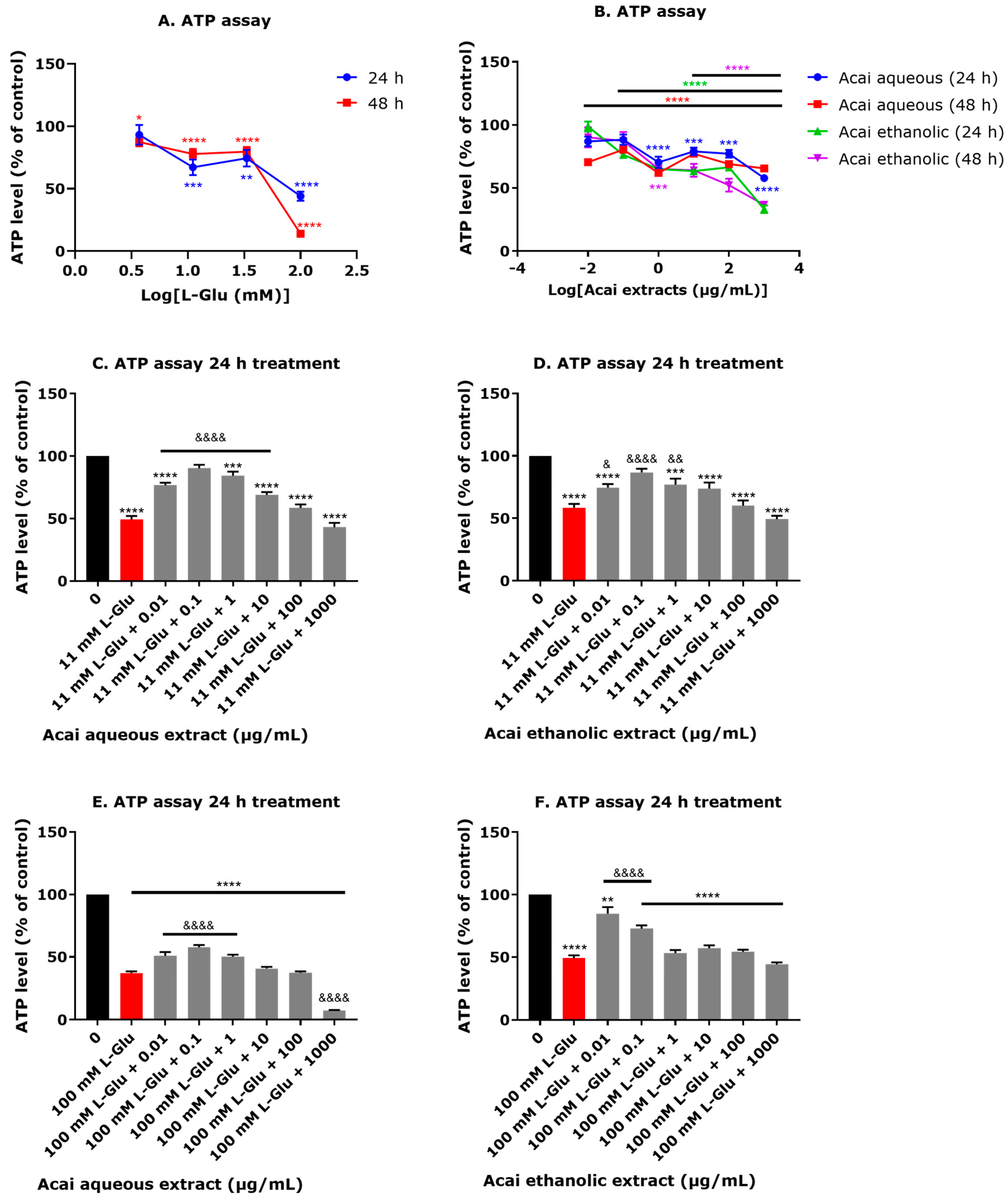
3.3. Acai Berry Extracts Preserve ATP Levels That Were Diminished after L-Glu Treatment
3.4. Acai Berry Extracts Restored the MMP Level That Was Reduced after L-Glu Treatment
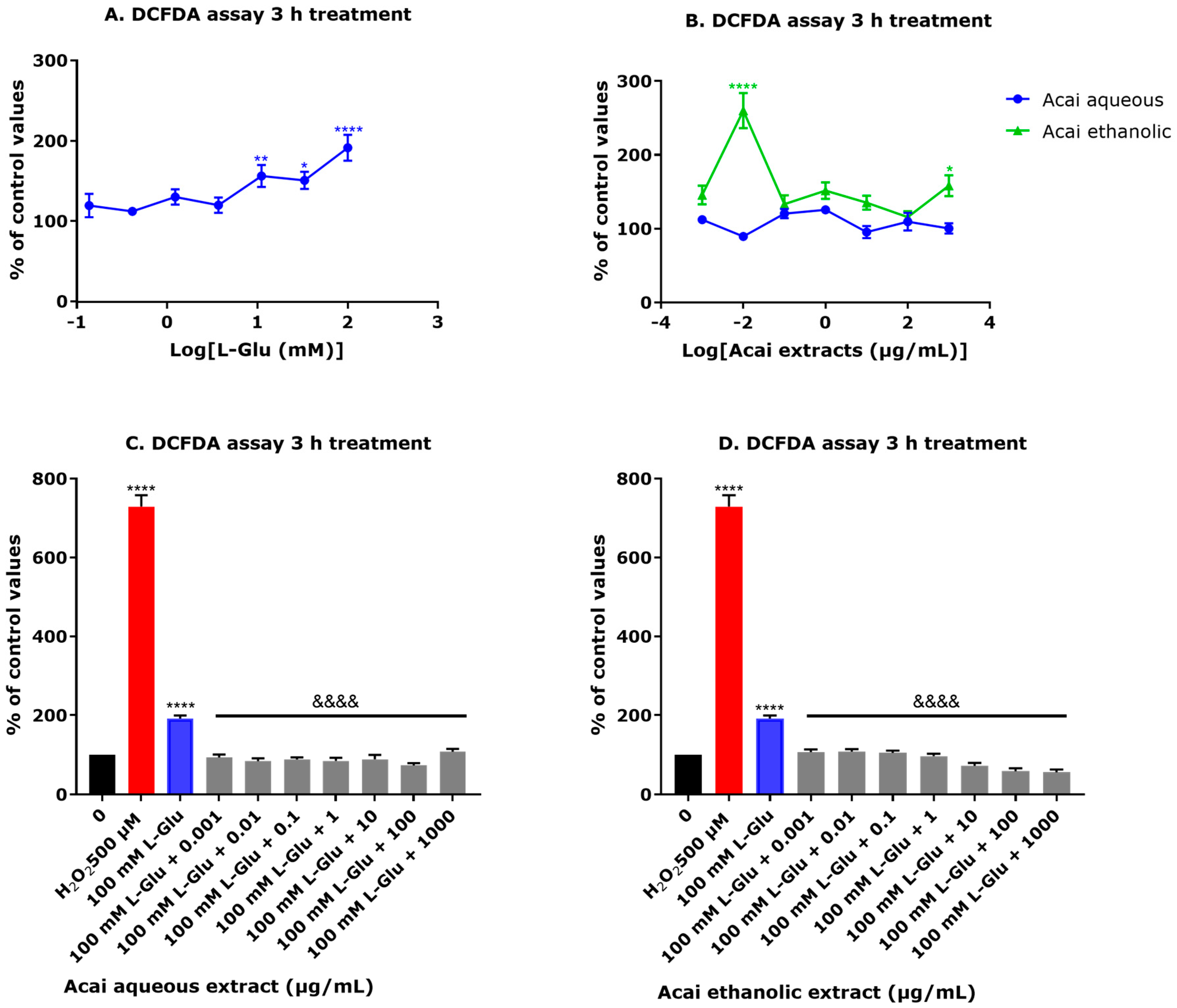
3.5. Acai Berry Extracts Significantly Reduced ROS Production Induced by L-Glu
3.6. Whole-Cell Patch-Clamp Assay
3.7. Liquid Chromatography-Mass Spectrometry (LC-MS)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moldovan, O.-L.; Rusu, A.; Tanase, C.; Vari, C.-E. Glutamate—A multifaceted molecule: Endogenous neurotransmitter, controversial food additive, design compound for anti-cancer drugs. A critical appraisal. Food Chem. Toxicol. 2021, 153, 112290. [Google Scholar] [CrossRef]
- Willard, S.; Koochekpour, S. Glutamate, glutamate receptors, and downstream signaling pathways. Int. J. Biol. Sci. 2013, 9, 948–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Danbolt, N. Glutamate as a neurotransmitter in the healthy brain. J. Neural Transm. 2014, 121, 799–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, J.; Markussen, K.; Jakobsen, E.; Schousboe, A.; Waagepetersen, H.; Rosenberg, P.; Aldana, B. Glutamate metabolism and recycling at the excitatory synapse in health and neurodegeneration. Neuropharmacology 2021, 196, 108719. [Google Scholar] [CrossRef]
- Mehta, A.; Prabhakar, M.; Kumar, P.; Deshmukh, R.; Sharma, P. Excitotoxicity: Bridge to various triggers in neurodegenerative disorders. Eur. J. Pharmacol. 2013, 698, 6–18. [Google Scholar] [CrossRef]
- Hawkins, R.; Viña, J. How glutamate is managed by the blood-brain barrier. Biology 2016, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Iovino, L.; Tremblay, M.; Civiero, L. Glutamate-induced excitotoxicity in Parkinson’s disease: The role of glial cells. J. Pharmacol. Sci. 2020, 144, 151–164. [Google Scholar] [CrossRef]
- Moussawi, K.; Riegel, A.; Nair, S.; Kalivas, P. Extracellular glutamate: Functional compartments operate in different concentration ranges. Front. Syst. Neurosci. 2011, 5, 94. [Google Scholar] [CrossRef] [Green Version]
- Zaragozá, R. Transport of amino acids across the blood-brain barrier. Front. Physiol. 2020, 11, 973. [Google Scholar] [CrossRef]
- Suzuki, H.; Kawakita, F.; Asada, R.; Nakano, F.; Nishikawa, H.; Fujimoto, M. Old but still hot target, glutamate-mediated neurotoxicity in stroke. Transl. Stroke Res. 2022, 13, 216–217. [Google Scholar] [CrossRef]
- Verma, M.; Lizama, B.; Chu, C. Excitotoxicity, calcium and mitochondria: A triad in synaptic neurodegeneration. Transl. Neurodegener. 2022, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Reddy, P. Role of glutamate and NMDA receptors in Alzheimer’s disease. J. Alzheimers Dis. 2017, 57, 1041–1048. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, F.; Mai, D.; Qu, S. Molecular mechanisms of glutamate toxicity in Parkinson’s disease. Front. Neurosci. 2020, 14, 585584. [Google Scholar] [CrossRef] [PubMed]
- Rajda, C.; Pukoli, D.; Bende, Z.; Majláth, Z.; Vécsei, L. Excitotoxins, mitochondrial and redox disturbances in multiple sclerosis. Int. J. Mol. Sci. 2017, 18, 353. [Google Scholar] [CrossRef] [Green Version]
- Van Damme, P.; Dewil, M.; Robberecht, W.; Van Den Bosch, L. Excitotoxicity and amyotrophic lateral sclerosis. Neurodegener. Dis. 2005, 2, 147–159. [Google Scholar] [CrossRef]
- Estrada Sánchez, A.; Mejía-Toiber, J.; Massieu, L. Excitotoxic neuronal death and the pathogenesis of Huntington’s disease. Arch. Med. Res. 2008, 39, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Zeron, M.; Chen, N.; Moshaver, A.; Lee, A.; Wellington, C.; Hayden, M.; Raymond, L. Mutant huntingtin enhances excitotoxic cell death. Mol. Cell. Neurosci. 2001, 17, 41–53. [Google Scholar] [CrossRef]
- Madeira, C.; Vargas-Lopes, C.; Brandão, C.; Reis, T.; Laks, J.; Panizzutti, R.; Ferreira, S. Elevated glutamate and glutamine levels in the cerebrospinal fluid of patients with probable Alzheimer’s disease and depression. Front. Psychiatry 2018, 9, 561. [Google Scholar] [CrossRef] [Green Version]
- ALNasser, M.; Mellor, I.; Carter, W. Is L-Glutamate toxic to neurons and thereby contributes to neuronal loss and neurodegeneration? A systematic review. Brain Sci. 2022, 12, 577. [Google Scholar] [CrossRef] [PubMed]
- Ankarcrona, M.; Dypbukt, J.; Bonfoco, E.; Zhivotovsky, B.; Orrenius, S.; Lipton, S.; Nicotera, P. Glutamate-induced neuronal death: A succession of necrosis or apoptosis depending on mitochondrial function. Neuron 1995, 15, 961–973. [Google Scholar] [CrossRef] [Green Version]
- Olney, J.; Wozniak, D.; Farber, N. Excitotoxic neurodegeneration in Alzheimer Disease: New hypothesis and new therapeutic strategies. Arch. Neurol. 1997, 54, 1234–1240. [Google Scholar] [CrossRef]
- Lewerenz, J.; Hewett, S.; Huang, Y.; Lambros, M.; Gout, P.; Kalivas, P.; Massie, A.; Smolders, I.; Methner, A.; Pergande, M.; et al. The cystine/glutamate antiporter system xc− in health and disease: From molecular mechanisms to novel therapeutic opportunities. Antioxid. Redox Signal. 2013, 18, 522–555. [Google Scholar] [CrossRef] [Green Version]
- Mattson, M. Excitotoxic and excitoprotective mechanisms. Neuromolecular Med. 2003, 3, 65–94. [Google Scholar] [CrossRef] [Green Version]
- Schubert, D.; Piasecki, D. Oxidative glutamate toxicity can be a component of the excitotoxicity cascade. J. Neurosci. 2001, 21, 7455–7462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, D.; Maulucci-Gedde, M.; Kriegstein, A. Glutamate neurotoxicity in cortical cell culture. J. Neurosci. 1987, 7, 357–368. [Google Scholar] [CrossRef]
- Kato, K.; Puttfarcken, P.; Lyons, E.; Coyle, J. Developmental time course and ionic dependence of kainate-mediated toxicity in rat cerebellar granule cell cultures. J. Pharmacol. Exp. Ther. 1991, 256, 402–411. [Google Scholar] [PubMed]
- Rothman, S.; Olney, J. Glutamate and the pathophysiology of hypoxic–ischemic brain damage. Ann. Neurol. 1986, 19, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M. Calcium and neurodegeneration. Aging cell 2007, 6, 337–350. [Google Scholar] [CrossRef]
- Atlante, A.; Calissano, P.; Bobba, A.; Giannattasio, S.; Marra, E.; Passarella, S. Glutamate neurotoxicity, oxidative stress and mitochondria. FEBS Lett. 2001, 497, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Duchen, M. Mitochondria and calcium: From cell signalling to cell death. J. Physiol. 2000, 529, 57–68. [Google Scholar] [CrossRef]
- Norenberg, M.; Rao, K. The mitochondrial permeability transition in neurologic disease. Neurochem. Int. 2007, 50, 983–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, C.; Oliveira, C. Oxidative glutamate toxicity involves mitochondrial dysfunction and perturbation of intracellular Ca2+ homeostasis. Neurosci. Res. 2000, 37, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewerenz, J.; Klein, M.; Methner, A. Cooperative action of glutamate transporters and cystine/glutamate antiporter system Xc– protects from oxidative glutamate toxicity. J. Neurochem. 2006, 98, 916–925. [Google Scholar] [CrossRef]
- Kuriakose, D.; Xiao, Z. Pathophysiology and treatment of stroke: Present status and future perspectives. Int. J. Mol. Sci. 2020, 21, 7609. [Google Scholar] [CrossRef]
- Prentice, H.; Modi, J.; Wu, J.-Y. Mechanisms of neuronal protection against excitotoxicity, endoplasmic reticulum stress, and mitochondrial dysfunction in stroke and neurodegenerative diseases. Oxid. Med. Cell. Longev. 2015, 2015, 964518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulose, S.; Fisher, D.; Larson, J.; Bielinski, D.; Rimando, A.; Carey, A.; Schauss, A.; Shukitt-Hale, B. Anthocyanin-rich açai (Euterpe oleracea Mart.) fruit pulp fractions attenuate inflammatory stress signaling in mouse brain BV-2 microglial cells. J. Agric. Food Chem. 2012, 60, 1084–1093. [Google Scholar] [CrossRef]
- ALNasser, M.; Mellor, I. Neuroprotective activities of acai berries (Euterpe sp.): A review. J. Herbmed Pharmacol. 2022, 11, 166–181. [Google Scholar] [CrossRef]
- Benatrehina, A.; Pan, L.; Naman, B.; Li, J.; Kinghorn, D. Usage, biological activity, and safety of selected botanical dietary supplements consumed in the United States. J. Tradit. Complement. Med. 2018, 8, 267–277. [Google Scholar] [CrossRef]
- Jensen, G.; Wu, X.; Patterson, K.; Barnes, J.; Carter, S.; Scherwitz, L.; Beaman, R.; Endres, J.; Schauss, A. In vitro and in vivo antioxidant and anti-inflammatory capacities of an antioxidant-rich fruit and berry juice blend. results of a pilot and randomized, double-blinded, placebo-controlled, crossover study. J. Agric. Food Chem. 2008, 56, 8326–8333. [Google Scholar] [CrossRef] [Green Version]
- Spada, P.; Dani, C.; Bortolini, G.; Funchal, C.; Henriques, J.; Salvador, M. Frozen fruit pulp of Euterpe oleraceae Mart. (acai) prevents hydrogen peroxide-induced damage in the cerebral cortex, cerebellum, and hippocampus of rats. J. Med. Food 2009, 12, 1084–1088. [Google Scholar] [CrossRef]
- De Oliveira, N.; Almeida, M.; Pontes, F.; Barcelos, M.; de Paula da Silva, C.; Rosa, J.; Cruz, R.; da Silva Hage-Melim, L. Antioxidant effect of flavonoids present in Euterpe oleracea Martius and neurodegenerative diseases: A literature review. Cent. Nerv. Syst. Agents Med. Chem. 2019, 19, 75–99. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.; Andreazza, A.; da Silva, T.; Boligon, A.; do Nascimento, V.; Scola, G.; Duong, A.; Cadona, F.; Ribeiro, E.; da Cruz, I. Neuroprotective effects of acai (Euterpe oleracea Mart.) against rotenone in vitro exposure. Oxid. Med. Cell. Longev. 2016, 2016, 8940850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalevich, J.; Langford, D. Considerations for the use of SH-SY5Y neuroblastoma cells in neurobiology. Methods Mol. Biol. 2013, 1078, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Suarez, L.; Awabdh, S.; Coumoul, X.; Chauvet, C. The SH-SY5Y human neuroblastoma cell line, a relevant in vitro cell model for investigating neurotoxicology in human: Focus on organic pollutants. Neurotoxicology 2022, 92, 131–155. [Google Scholar] [CrossRef] [PubMed]
- Donato, R.; Miljan, E.; Hines, S.; Aouabdi, S.; Pollock, K.; Patel, S.; Edwards, F.; Sinden, J. Differential development of neuronal physiological responsiveness in two human neural stem cell lines. BMC Neurosci. 2007, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Brocke, K.; Staufner, C.; Luksch, H.; Geiger, K.; Stepulak, A.; Marzahn, J.; Schackert, G.; Temme, A.; Ikonomidou, C. Glutamate receptors in pediatric tumors of the central nervous system. Cancer Biol. Ther. 2010, 9, 455–468. [Google Scholar] [CrossRef] [Green Version]
- Luksch, H.; Uckermann, O.; Stepulak, A.; Hendruschk, S.; Marzahn, J.; Bastian, S.; Staufner, C.; Temme, A.; Ikonomidou, C. Silencing of selected glutamate receptor subunits modulates cancer growth. Anticancer. Res. 2011, 31, 3181–3192. [Google Scholar]
- Stepulak, A.; Luksch, H.; Gebhardt, C.; Uckermann, O.; Marzahn, J.; Sifringer, M.; Rzeski, W.; Staufner, C.; Brocke, K.; Turski, L.; et al. Expression of glutamate receptor subunits in human cancers. Histochem. Cell. Biol. 2009, 132, 435–445. [Google Scholar] [CrossRef]
- Wojciech, R.; Turski, L.; Ikonomidou, C. Glutamate antagonists limit tumor growth. Proc. Natl. Acad. Sci. USA 2001, 98, 6372–6377. [Google Scholar] [CrossRef] [Green Version]
- ALNasser, M.; Mellor, I.; Carter, W. A preliminary assessment of the nutraceutical potential of acai berry (Euterpe sp.) as a potential natural treatment for Alzheimer’s disease. Molecules 2022, 27, 4891. [Google Scholar] [CrossRef]
- Nwidu, L.; Elmorsy, E.; Aprioku, J.; Siminialayi, I.; Carter, W. In vitro anti-cholinesterase and antioxidant activity of extracts of Moringa oleifera plants from Rivers State, Niger Delta, Nigeria. Medicines 2018, 5, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, D.; Musgrave, I.; Harvey, B.; Smid, S. Açaí (Euterpe oleraceae Mart.) berry extract exerts neuroprotective effects against β-amyloid exposure in vitro. Neurosci. Lett. 2013, 556, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Amat-Ur-Rasool, H.; Ahmed, M.; Hasnain, S.; Carter, W. Anti-cholinesterase combination drug therapy as a potential treatment for Alzheimer’s disease. Brain Sci. 2021, 11, 184. [Google Scholar] [CrossRef]
- de Medeiros, L.; De Bastiani, M.; Rico, E.; Schonhofen, P.; Pfaffenseller, B.; Wollenhaupt-Aguiar, B.; Grun, L.; Barbé-Tuana, F.; Zimmer, E.; Castro, M.; et al. Cholinergic differentiation of human neuroblastoma SH-SY5Y cell line and its potential use as an in vitro model for Alzheimer’s disease studies. Mol. Neurobiol. 2019, 56, 7355–7367. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, J.; Dewilde, A.; Pal, A.; Bello, D.; Therrien, J.; Braunhut, S.; Marx, K. Understanding and correcting for carbon nanotube interferences with a commercial LDH cytotoxicity assay. Toxicology 2012, 299, 99–111. [Google Scholar] [CrossRef]
- Lee, M.-S.; Park, W.-S.; Kim, Y.; Ahn, W.; Kwon, S.-H.; Her, S. Intracellular ATP assay of live cells using PTD-conjugated luciferase. Sensors 2012, 12, 15628–15637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Ghafari, A.; Elmorsy, E.; Fikry, E.; Alrowaili, M.; Carter, W. The heavy metals lead and cadmium are cytotoxic to human bone osteoblasts via induction of redox stress. PLoS ONE 2019, 14, e0225341. [Google Scholar] [CrossRef] [Green Version]
- Abdelrazig, S.; Safo, L.; Rance, G.; Fay, M.; Theodosiou, E.; Topham, P.; Kim, D.-H.; Fernández-Castané, A. Metabolic characterisation of Magnetospirillum gryphiswaldense MSR-1 using LC-MS-based metabolite profiling. RSC Adv. 2020, 10, 32548–32560. [Google Scholar] [CrossRef] [PubMed]
- Babiaka, S.; Nia, R.; Abuga, K.; Mbah, J.; Nziko, V.; Paper, D.; Ntie-Kang, F. Antioxidant potential of flavonoid glycosides from Manniophyton fulvum Müll. (Euphorbiaceae): Identification and molecular modeling. Sci. Afr. 2020, 8, e00423. [Google Scholar] [CrossRef]
- Bolouri-Moghaddam, M.; Le Roy, K.; Xiang, L.; Rolland, F.; Van den Ende, W. Sugar signalling and antioxidant network connections in plant cells. FEBS J. 2010, 277, 2022–2037. [Google Scholar] [CrossRef]
- Ercan, L.; Dogru, M. Antioxidant and antimicrobial capacity of quinic acid. BEU J. Sci. Technol. 2022, 11, 1018–1025. [Google Scholar] [CrossRef]
- Xu, J.-G.; Hu, Q.-P.; Liu, Y. Antioxidant and DNA-protective activities of chlorogenic acid isomers. J. Agric. Food Chem. 2012, 60, 11625–11630. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, J.; Ferreres, F.; Gil-Izquierdo, Á.; Valentao, P.; Andrade, P. Medicinal species as MTDLs: Turnera diffusa Willd. Ex Schult inhibits CNS enzymes and delays glutamate excitotoxicity in SH-SY5Y cells via oxidative damage. Food Chem. Toxicol. 2017, 106, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Brizi, C.; Santulli, C.; Micucci, M.; Budriesi, R.; Chiarini, A.; Aldinucci, C.; Frosini, M. Neuroprotective effects of Castanea sativa Mill. bark extract in human neuroblastoma cells subjected to oxidative stress. J. Cell. Biochem. 2016, 117, 510–520. [Google Scholar] [CrossRef]
- de Oliveira, M.; Duarte, A.; Chenet, A.; de Almeida, F.; Andrade, C. Carnosic acid pretreatment attenuates mitochondrial dysfunction in SH-SY5Y Cells in an experimental model of glutamate-induced excitotoxicity. Neurotox. Res. 2019, 36, 551–562. [Google Scholar] [CrossRef]
- Lee, H.; Spandidos, D.; Tsatsakis, A.; Margina, D.; Izotov, B.; Yang, S. Neuroprotective effects of Scrophularia buergeriana extract against glutamate-induced toxicity in SH-SY5Y cells. Int. J. Mol. Med. 2019, 43, 2144–2152. [Google Scholar] [CrossRef]
- Li, H.; Han, W.; Wang, H.; Ding, F.; Xiao, L.; Shi, R.; Ai, L.; Huang, Z. Tanshinone IIA inhibits glutamate-induced oxidative toxicity through prevention of mitochondrial dysfunction and suppression of MAPK activation in SH-SY5Y human neuroblastoma cells. Oxid. Med. Cell. Longev. 2017, 2017, 4517486. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.-W.; Zhang, L.; Zhu, S.-J.; Chen, W.-C.; Mei, B. Excitotoxicity effects of glutamate on human neuroblastoma SH-SY5Y cells via oxidative damage. Neurosci. Bull. 2010, 26, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Xin, H.; Cui, Y.; An, Z.; Yang, Q.; Zou, X.; Yu, N. Attenuated glutamate induced ROS production by antioxidative compounds in neural cell lines. RSC Adv. 2019, 9, 34735–34743. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Han, A.; Kim, E.; Yang, J.; Ahn, J.; Na, J.; Cho, S. KHG21834 attenuates glutamate-induced mitochondrial damage, apoptosis, and NLRP3 inflammasome activation in SH-SY5Y human neuroblastoma cells. Eur. J. Pharmacol. 2019, 856, 172412. [Google Scholar] [CrossRef]
- Yuksel, T.; Yayla, M.; Halici, Z.; Cadirci, E.; Polat, B.; Kose, D. Protective effect of 5-HT7 receptor activation against glutamate-induced neurotoxicity in human neuroblastoma SH-SY5Y cells via antioxidative and antiapoptotic pathways. Neurotoxicol Teratol. 2019, 72, 22–28. [Google Scholar] [CrossRef]
- Cadoná, F.; de Souza, D.; Fontana, T.; Bodenstein, D.; Ramos, A.; Sagrillo, M.; Salvador, M.; Mota, K.; Davidson, C.; Ribeiro, E.; et al. Açaí (Euterpe oleracea Mart.) as a potential anti-neuroinflammatory agent: NLRP3 priming and activating signal pathway modulation. Mol. Neurobiol. 2021, 58, 4460–4476. [Google Scholar] [CrossRef]
- Arrifano, G.; Lichtenstein, M.; Souza-Monteiro, J.; Farina, M.; Rogez, H.; Carvalho, J.; Suñol, C.; Crespo-López, M. Clarified açaí (Euterpe oleracea) juice as an anticonvulsant agent: In vitro mechanistic study of GABAergic targets. Oxid. Med. Cell. Longev. 2018, 2018, 2678089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza, D.; Pappis, L.; Bandeira, T.; Sangoi, G.; Fontana, T.; Rissi, V.; Sagrillo, M.; Duarte, M.; Duarte, T.; Bodenstein, D.; et al. Açaí (Euterpe oleracea Mart.) presents anti-neuroinflammatory capacity in LPS-activated microglia cells. Nutr. Neurosci. 2020, 25, 1188–1199. [Google Scholar] [CrossRef] [PubMed]
- Torma, P.; Brasil, A.; Carvalho, A.; Jablonski, A.; Rabelo, T.; Moreira, J.; Gelain, D.; Flôres, S.; Augusti, P.; Rios, A. Hydroethanolic extracts from different genotypes of açaí (Euterpe oleracea) presented antioxidant potential and protected human neuron-like cells (SH-SY5Y). Food Chem. 2017, 222, 94–104. [Google Scholar] [CrossRef]
- Sun, X.; Shi, X.; Lu, L.; Jiang, Y.; Liu, B. Stimulus-dependent neuronal cell responses in SH-SY5Y neuroblastoma cells. Mol. Med. Rep. 2016, 13, 2215–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Wang, K.; Zhang, K.; Lin, X.; Zhu, L.; Zhou, F. Puerarin protects human neuroblastoma SH-SY5Y cells against glutamate-induced oxidative stress and mitochondrial dysfunction. J. Biochem. Mol. Toxicol. 2016, 30, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, P.; Sulejczak, D.; Kleczkowska, P.; Bukowska-Ośko, I.; Kucia, M.; Popiel, M.; Wietrak, E.; Kramkowski, K.; Wrzosek, K.; Kaczyńska, K. Mitochondrial oxidative stress-A causative factor and therapeutic target in many diseases. Int. J. Mol. Sci. 2021, 22, 13384. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Chen, M.; Jiang, J. Mitochondrial dysfunction in neurodegenerative diseases and drug targets via apoptotic signaling. Mitochondrion 2019, 49, 35–45. [Google Scholar] [CrossRef]
- De Almeida Magalhães, T.; De Oliveira Macedo, P.; Converti, A.; Neves de Lima, A. The use of Euterpe oleracea Mart. as a new perspective for disease treatment and prevention. Biomolecules 2020, 10, 813. [Google Scholar] [CrossRef]
- Laurindo, L.; Barbalho, S.; Araújo, A.; Guiguer, E.; Mondal, A.; Bachtel, G.; Bishayee, A. Açaí (Euterpe oleracea Mart.) in health and disease: A critical review. Nutrients 2023, 15, 989. [Google Scholar] [CrossRef]
- Altemimi, A.; Lakhssassi, N.; Baharlouei, A.; Watson, D.; Lightfoot, D. Phytochemicals: Extraction, isolation, and identification of bioactive compounds from plant extracts. Plants 2017, 6, 42. [Google Scholar] [CrossRef]
- Chung, H.-J. Physiological activity of acai berry (Euterpe oleracea Mart.) extracted with different solvents. J. Korean Soc. Food Cult. 2012, 27, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Jantas, D.; Pytel, M.; Mozrzymas, J.; Leskiewicz, M.; Regulska, M.; Antkiewicz-Michaluk, L.; Lason, W. The attenuating effect of memantine on staurosporine-, salsolinol- and doxorubicin-induced apoptosis in human neuroblastoma SH-SY5Y cells. Neurochem. Int. 2008, 52, 864–877. [Google Scholar] [CrossRef]
- Bebitoglu, B.; Elif, O.; Acet, N.; Hodxiz, A.; Temel, F.; Saniye, A.; Kilickap, A. The neuroprotective effect of lamotrigine against glutamate excitotoxicity in SH-SY5Y human neuroblastoma cells. Marmara Med. J. 2020, 33, 146–152. [Google Scholar] [CrossRef]
- Naarala, J.; Nykvist, P.; Tuomala, M.; Savolainen, K. Excitatory amino acid-induced slow biphasic responses of free intracellular calcium in human neuroblastoma cells. FEBS Lett. 1993, 330, 222–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naarala, J.; Tervo, P.; Loikkanen, J.; Savolainen, K. Blocking of carbachol-induced calcium mobilization by glutamate receptor antagonists. Neurosci. Res. Commun. 2002, 30, 1–6. [Google Scholar] [CrossRef]
- Nair, V.; Niznik, H.; Mishra, R. Interaction of NMDA and dopamine D2L receptors in human neuroblastoma SH-SY5Y cells. J. Neurochem. 1996, 66, 2390–2393. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Murali, S. Stimulation of Na+-K+-2Cl− cotransporter in neuronal cells by excitatory neurotransmitter glutamate. Am. J. Physiol. 1998, 275, C772–C779. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, G.; Szeto, S.; Chong, C.; Quan, Q.; Huang, C.; Cui, W.; Guo, B.; Wang, Y.; Han, Y.; et al. Examining the neuroprotective effects of protocatechuic acid and chrysin on in vitro and in vivo models of Parkinson disease. Free. Radic. Biol. Med. 2015, 84, 331–343. [Google Scholar] [CrossRef]
- Szwajgier, D.; Borowiec, K.; Pustelniak, K. The Neuroprotective effects of phenolic acids: Molecular mechanism of action. Nutrients 2017, 9, 477. [Google Scholar] [CrossRef] [Green Version]
- Ogut, E.; Armagan, K.; Gül, Z. The role of syringic acid as a neuroprotective agent for neurodegenerative disorders and future expectations. Metab. Brain Dis. 2022, 37, 859–880. [Google Scholar] [CrossRef] [PubMed]
- Ullah, R.; Ikram, M.; Park, T.; Ahmad, R.; Saeed, K.; Alam, S.; Rehman, I.; Khan, A.; Khan, I.; Jo, M.; et al. Vanillic acid, a bioactive phenolic compound, counteracts LPS-Induced neurotoxicity by regulating c-Jun N-terminal kinase in mouse brain. Int. J. Mol. Sci. 2020, 22, 361. [Google Scholar] [CrossRef] [PubMed]
- Winter, A.; Brenner, M.; Punessen, N.; Snodgrass, M.; Byars, C.; Arora, Y.; Linseman, D. Comparison of the neuroprotective and anti-Inflammatory effects of the anthocyanin metabolites, protocatechuic acid and 4-hydroxybenzoic acid. Oxid. Med. Cell. Longev. 2017, 2017, 6297080. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Jahan, S.; Imtiyaz, Z.; Alshahrani, S.; Antar Makeen, H.; Mohammed Alshehri, B.; Kumar, A.; Arafah, A.; Rehman, M. Neuroprotection: Targeting multiple pathways by naturally occurring phytochemicals. Biomedicines 2020, 8, 284. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Saito, S.; Tanaka, M.; Yamakage, H.; Kusakabe, T.; Shimatsu, A.; Ihara, M.; Satoh-Asahara, N. Pleiotropic neuroprotective effects of taxifolin in cerebral amyloid angiopathy. Proc. Natl. Acad. Sci. USA 2019, 116, 10031–10038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhao, Y.; Tan, X.; Liu, J.; Zhi, Y.; Yi, L.; Bai, S.; Du, Q.; Li, Q.X.; Dong, Y. Isoorientin inhibits inflammation in macrophages and endotoxemia mice by regulating glycogen synthase kinase 3β. Mediat. Inflamm. 2020, 2020, 8704146. [Google Scholar] [CrossRef]
- Ahmad, I.; Kumar, D.; Patel, H. Computational investigation of phytochemicals from Withania somnifera (Indian ginseng/ashwagandha) as plausible inhibitors of GluN2B-containing NMDA receptors. J. Biomol. Struct. Dyn. 2022, 40, 7991–8003. [Google Scholar] [CrossRef]
- David, T.; Omotuyi, O.; Agboola, O.; Okonkwo, D.; Adelakun, N. Identification of Gly/NMDAR antagonist from Chromolaena odorata’s derived phytoconstituents using induced fit docking approach. bioRxiv 2019. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Compound | Formula | Exact Mass | Acai Berry Extracts | Antioxidant Activity | |
|---|---|---|---|---|---|
| PBS | Ethanol | ||||
| Phenolic compounds and phenolic acids: | |||||
| Protocatechuic acid | C7H6O4 | 154.0268 | Detected | Detected | ✓ |
| Syringic acid | C9H10O5 | 198.0530 | Detected | Detected | ✓ |
| Vanillic acid | C8H8O4 | 168.0425 | Detected | Detected | ✓ |
| Gallic acid | C7H6O5 | 170.0217 | Detected | Not Detected | ✓ |
| 4-Hydroxybenzoic acid | C7H6O3 | 138.0319 | Detected | Detected | ✓ |
| Benzoic acid | C7H6O2 | 122.0371 | Detected | Not Detected | ✓ |
| 2,5-Dihydroxybenzoic acid | C7H6O4 | 154.0268 | Detected | Detected | ✓ |
| Chlorogenic acid | C16H18O9 | 354.0951 | Detected | Detected | ✓ |
| Flavonoids: | |||||
| Dihydrokaempferol | C15H12O6 | 288.0634 | Detected | Detected | ✓ |
| Luteolin | C15H10O6 | 286.0478 | Not Detected | Detected | ✓ |
| Quercetin | C15H10O7 | 302.0426 | Not Detected | Detected | ✓ |
| Taxifolin deoxyhexose or Taxifolin | C15H12O7 | 304.0584 | Detected | Detected | ✓ |
| Quercetin-3-O-rutinoside (rutin) | C27H30O16 | 610.1519 | Not Detected | Detected | ✓ |
| Quercetin 3-O-glucoside (Isoquercitrin) | C21H20O12 | 464.0950 | Not Detected | Detected | ✓ |
| Kaempferol rhamnoside | C21H20O10 | 432.1048 | Detected | Not Detected | ✓ |
| Isoorientin | C21H20O11 | 448.0996 | Not Detected | Detected | ✓ |
| Lignans: | |||||
| (+)-Isolariciresinol | C20H24O6 | 360.1575 | Detected | Detected | ✓ |
| (+)-lariciresinol | C20H24O6 | 360.1575 | Detected | Detected | ✓ |
| Dihydroconiferyl alcohol | C10H14O3 | 182.0945 | Detected | Detected | - |
| Proanthocyanidins: | |||||
| (+)-Catechin | C15H14O6 | 290.0785 | Not Detected | Detected | ✓ |
| Monoterpenoids: | |||||
| (+)-menthiafolic acid | C10H16O3 | 184.1100 | Detected | Not Detected | - |
| (E,Z)-2,6-dimethyl-2,6-octadiene-1,8-diol | C10H18O2 | 170.1309 | Not Detected | Detected | - |
| Norisoprenoids: | |||||
| (-)-loliolide | C11H16O3 | 196.1099 | Not Detected | Detected | ✓ |
| Major fatty acids: | |||||
| Monounsaturated fatty acids | |||||
| Oleic acid | C18H34O2 | 282.2558 | Detected | Detected | ✓ |
| Palmitoleic acid | C16H30O2 | 254.2247 | Not Detected | Detected | - |
| Polyunsaturated fatty acids | |||||
| Linoleic acid | C18H32O2 | 280.2401 | Not Detected | Detected | ✓ |
| Linolenic acid | C18H30O2 | 278.2245 | Not Detected | Detected | ✓ |
| Saturated fatty acids | |||||
| Palmitic acid | C16H32O2 | 256.2403 | Not Detected | Detected | - |
| Stearic acid | C18H36O2 | 284.2714 | Not Detected | Detected | - |
| Amino acids: | |||||
| Alanine | C3H7NO2 | 89.0477 | Detected | Detected | - |
| Lysine | C6H14N2O2 | 146.1055 | Detected | Not Detected | ✓ |
| Arginine | C6H14N4O2 | 174.1116 | Not Detected | Detected | ✓ |
| Methionine | C5H11NO2S | 149.0511 | Detected | Not Detected | ✓ |
| Phenylalanine | C9H11NO2 | 165.0790 | Detected | Detected | - |
| Proline | C5H9NO2 | 115.0632 | Detected | Detected | - |
| Glutamic acid | C5H9NO4 | 147.0532 | Detected | Detected | - |
| Serine | C3H7NO3 | 105.0426 | Detected | Detected | - |
| Glycine | C2H5NO2 | 75.0320 | Detected | Detected | - |
| Threonine | C4H9NO3 | 119.0582 | Detected | Detected | - |
| Histidine | C6H9N3O2 | 155.0694 | Detected | Detected | ✓ |
| Tryptophan | C11H12N2O2 | 204.0901 | Detected | Detected | ✓ |
| Tyrosine | C9H11NO3 | 181.0741 | Detected | Detected | ✓ |
| Isoleucine | C6H13NO2 | 131.0946 | Detected | Detected | - |
| Valine | C5H11NO2 | 117.0788 | Detected | Detected | - |
| Leucine | C6H13NO2 | 131.0948 | Not Detected | Detected | - |
| Other compounds: | |||||
| Cellotetraose | C24H42O21 | 666.2225 | Not Detected | Detected | - |
| Sucrose | C12H22O11 | 342.1155 | Detected | Detected | ✓ |
| Quinic acid isomer 1 | C7H12O6 | 192.0635 | Detected | Detected | ✓ |
| Ascorbic acid (Vitamin C) | C6 H8 O6 | 176.0322 | Detected | Detected | ✓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
ALNasser, M.N.; AlSaadi, A.M.; Whitby, A.; Kim, D.-H.; Mellor, I.R.; Carter, W.G. Acai Berry (Euterpe sp.) Extracts Are Neuroprotective against L-Glutamate-Induced Toxicity by Limiting Mitochondrial Dysfunction and Cellular Redox Stress. Life 2023, 13, 1019. https://doi.org/10.3390/life13041019
ALNasser MN, AlSaadi AM, Whitby A, Kim D-H, Mellor IR, Carter WG. Acai Berry (Euterpe sp.) Extracts Are Neuroprotective against L-Glutamate-Induced Toxicity by Limiting Mitochondrial Dysfunction and Cellular Redox Stress. Life. 2023; 13(4):1019. https://doi.org/10.3390/life13041019
Chicago/Turabian StyleALNasser, Maryam N., Ayman M. AlSaadi, Alison Whitby, Dong-Hyun Kim, Ian R. Mellor, and Wayne G. Carter. 2023. "Acai Berry (Euterpe sp.) Extracts Are Neuroprotective against L-Glutamate-Induced Toxicity by Limiting Mitochondrial Dysfunction and Cellular Redox Stress" Life 13, no. 4: 1019. https://doi.org/10.3390/life13041019