
Autonomic Nervous System Adaptation and Circadian Rhythm Disturbances of the Cardiovascular System in a Ground-Based Murine Model of Spaceflight
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals
2.3. Surgery
2.4. Hindlimb Unloading
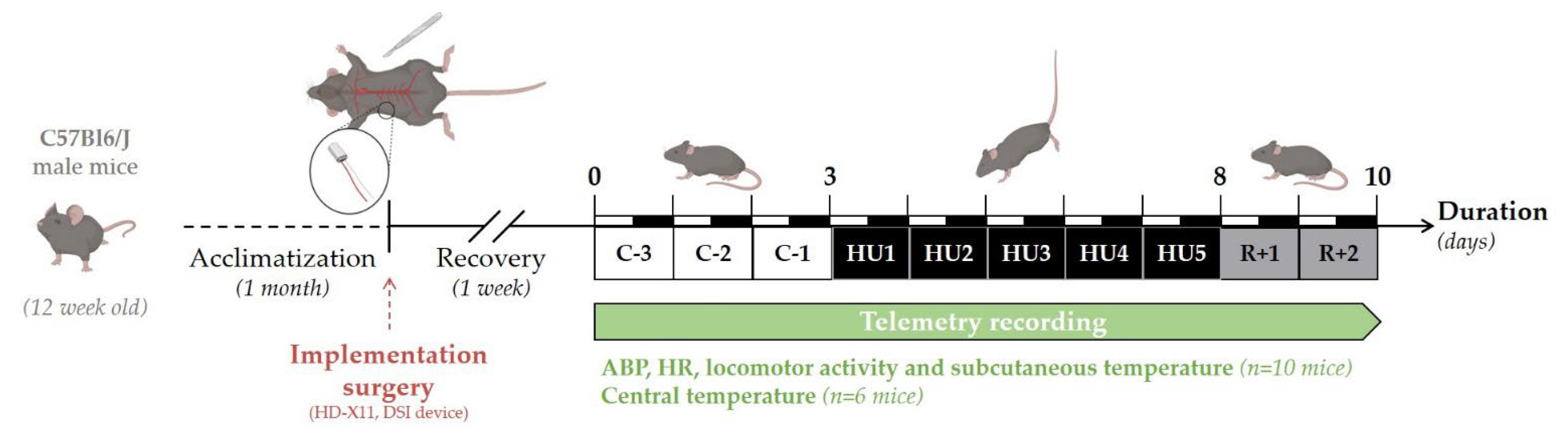
2.5. Experimental Design
2.6. Telemetric Data Collection and Analysis
2.7. Heart Rate Variability Analysis
2.8. Baroreflex Sensitivity Analysis
2.9. Statistical Analysis
3. Results
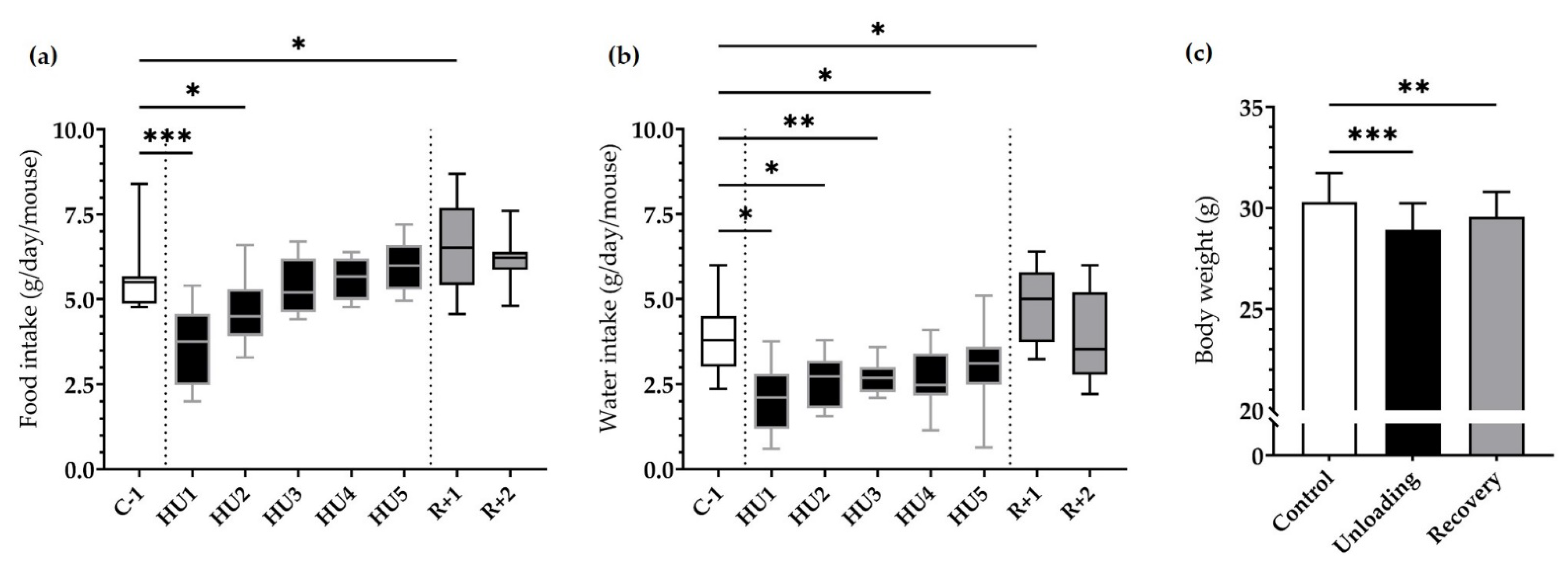
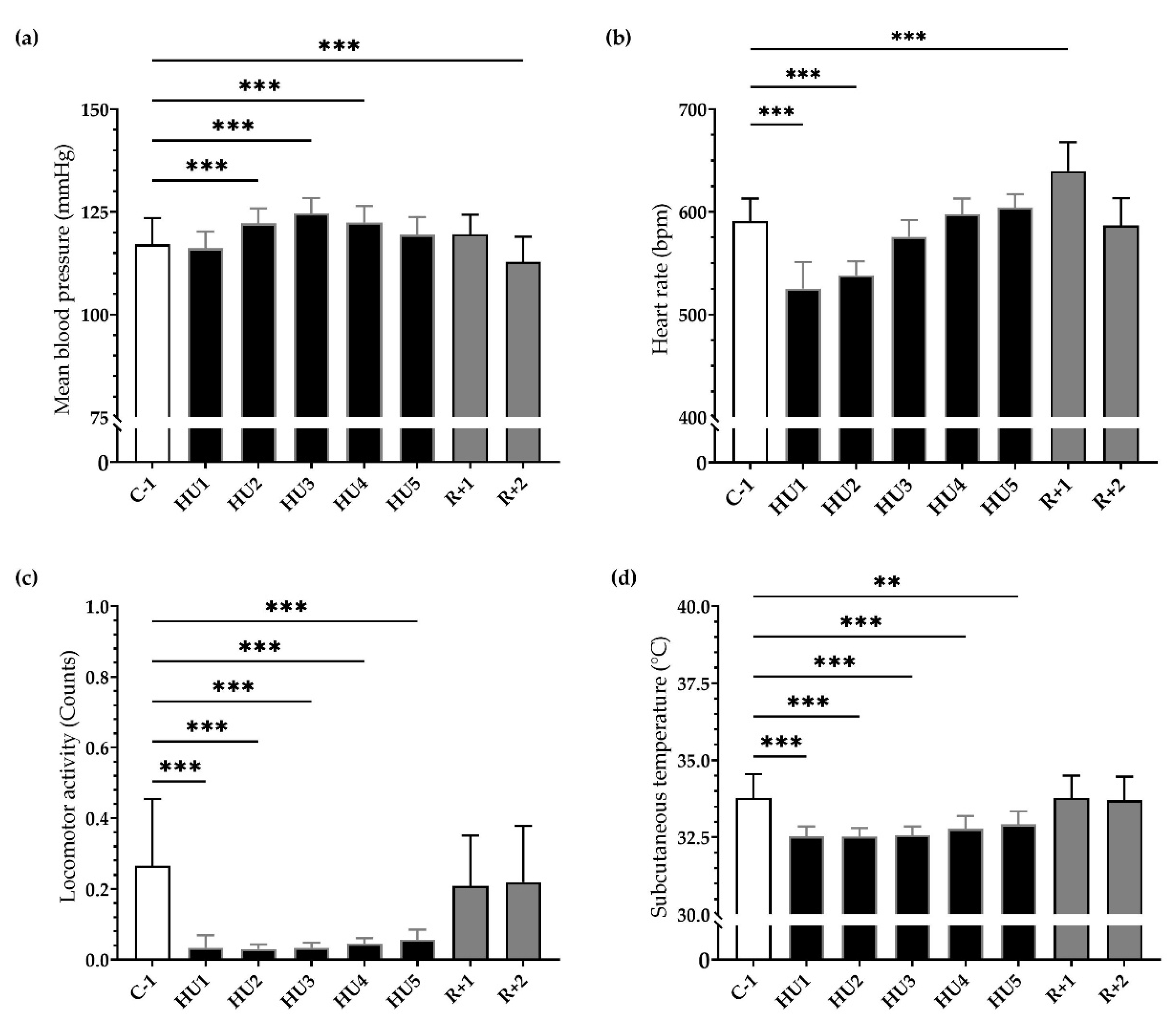
3.1. Physiological Phenotyping
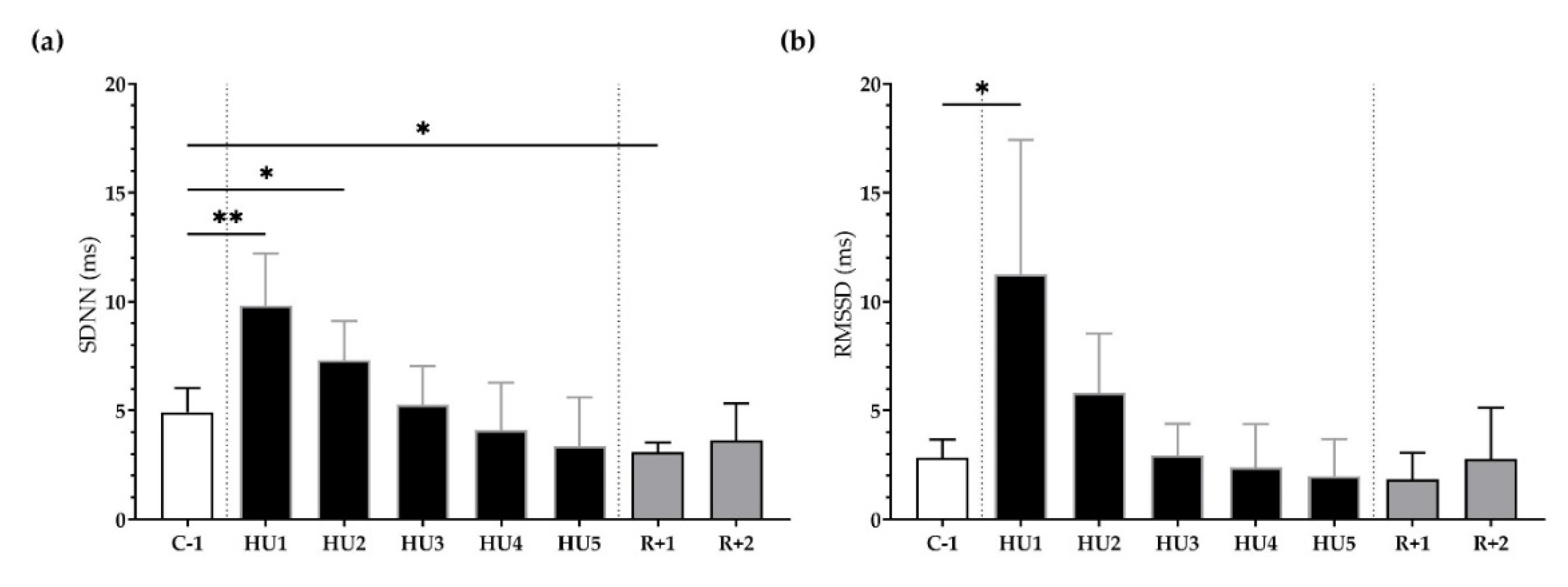
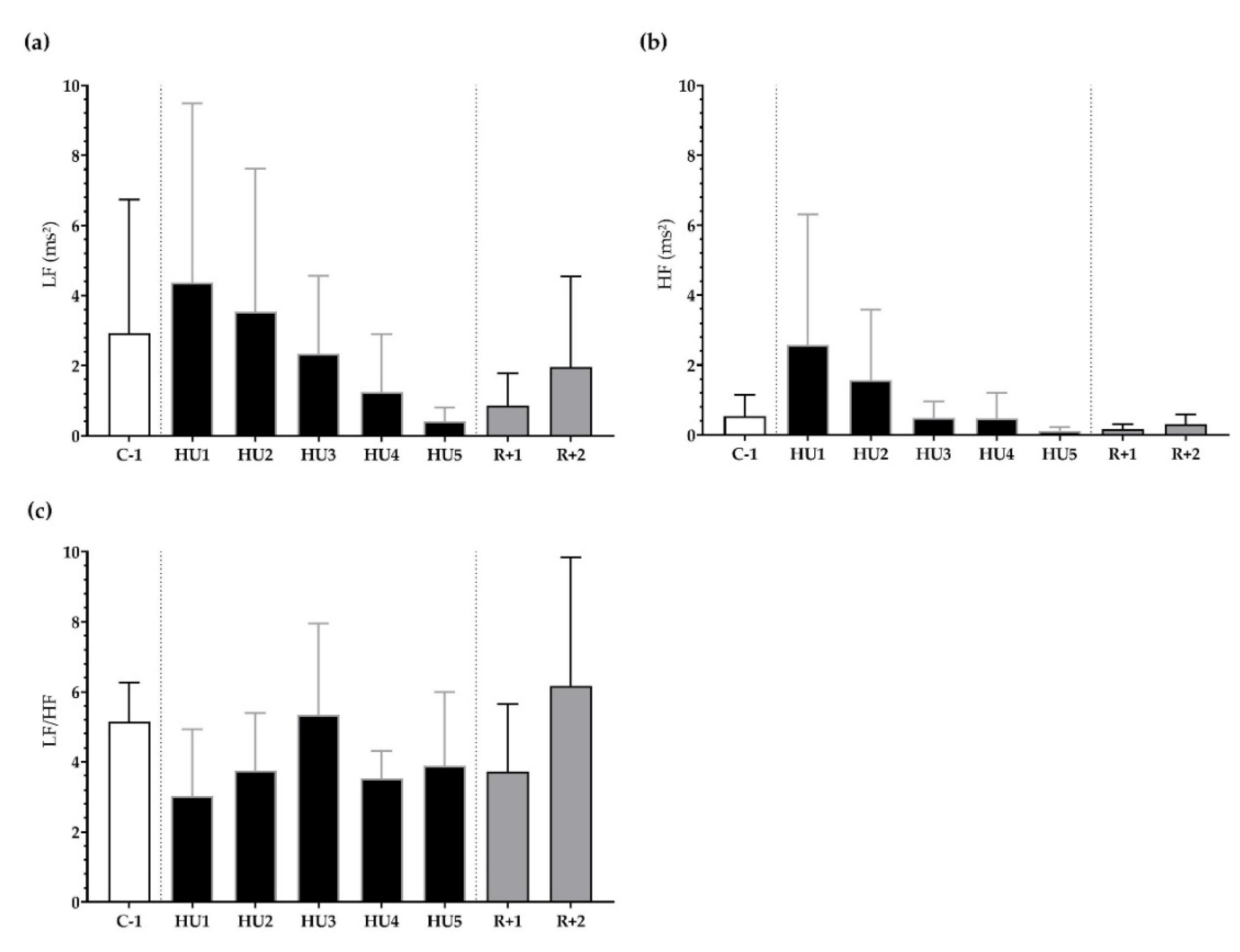
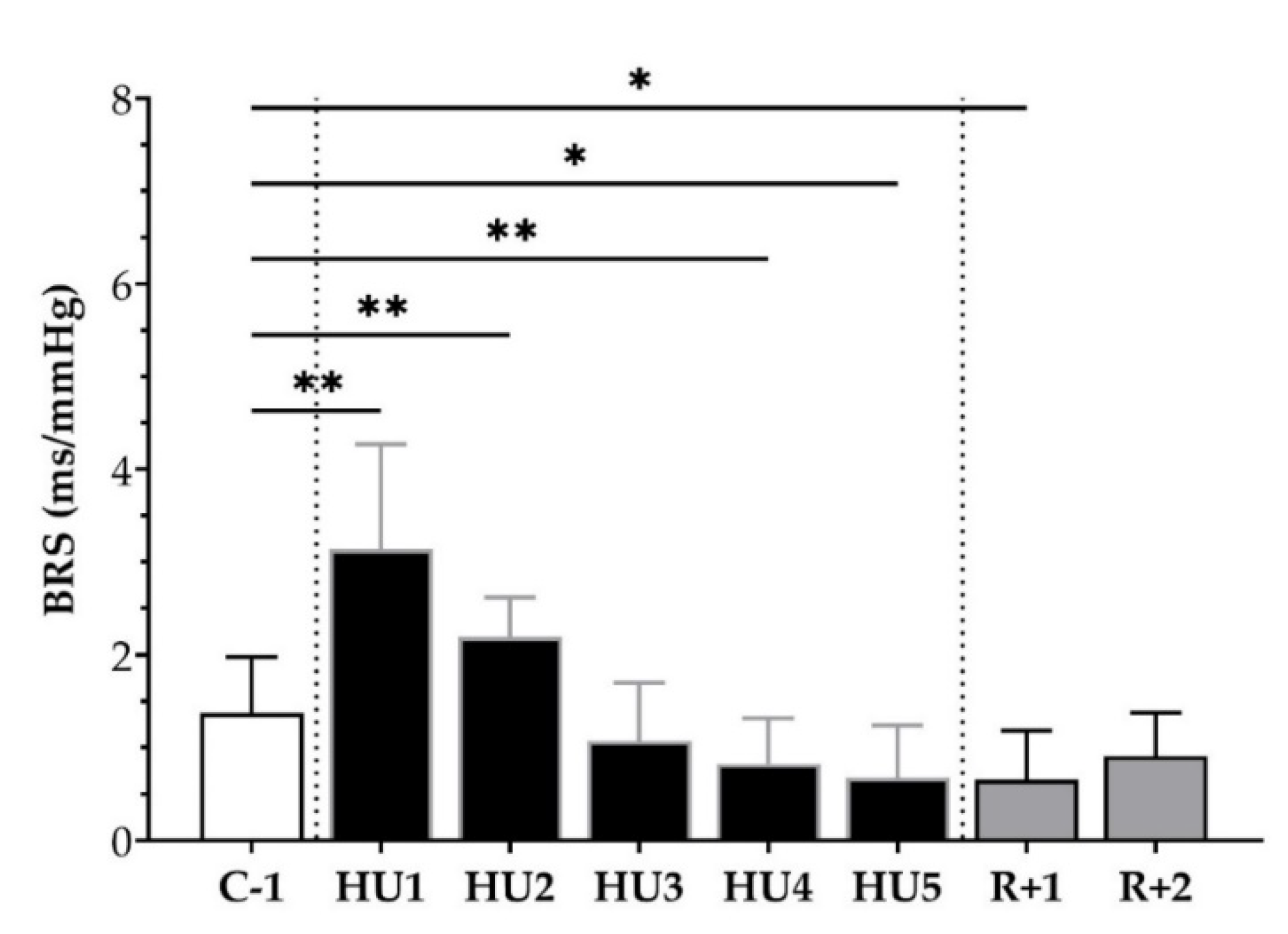
3.2. ANS Activity Analysis
3.3. Phenotyping Circadian Rhythms
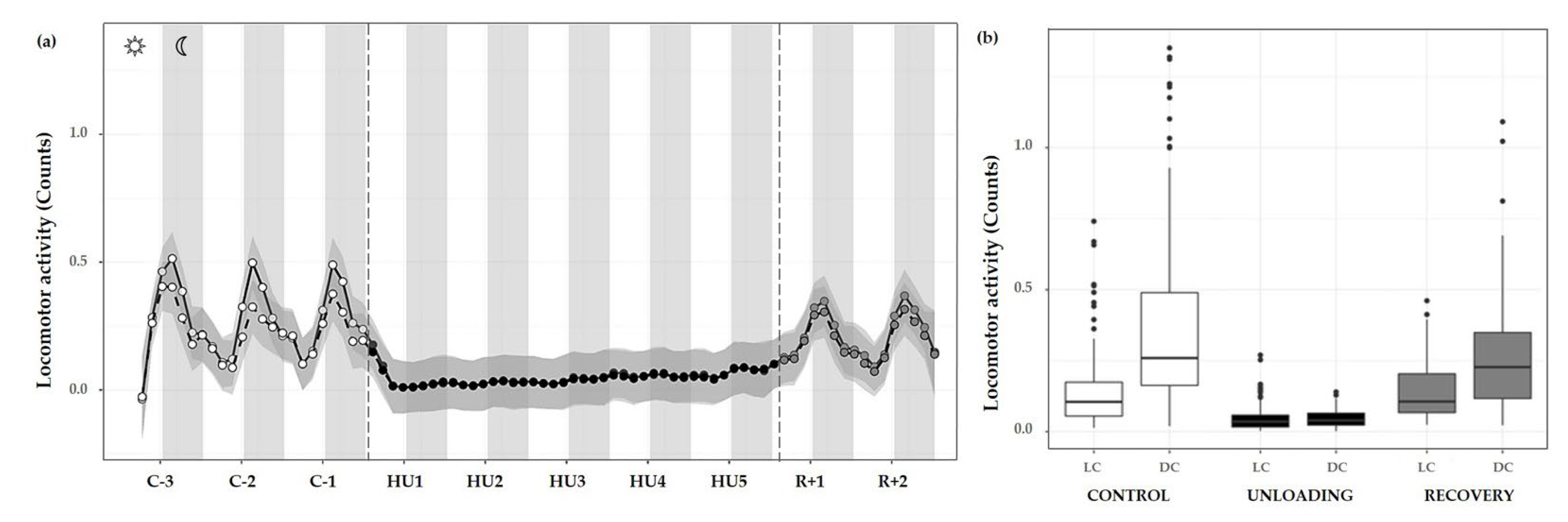
3.3.1. Locomotor Activity
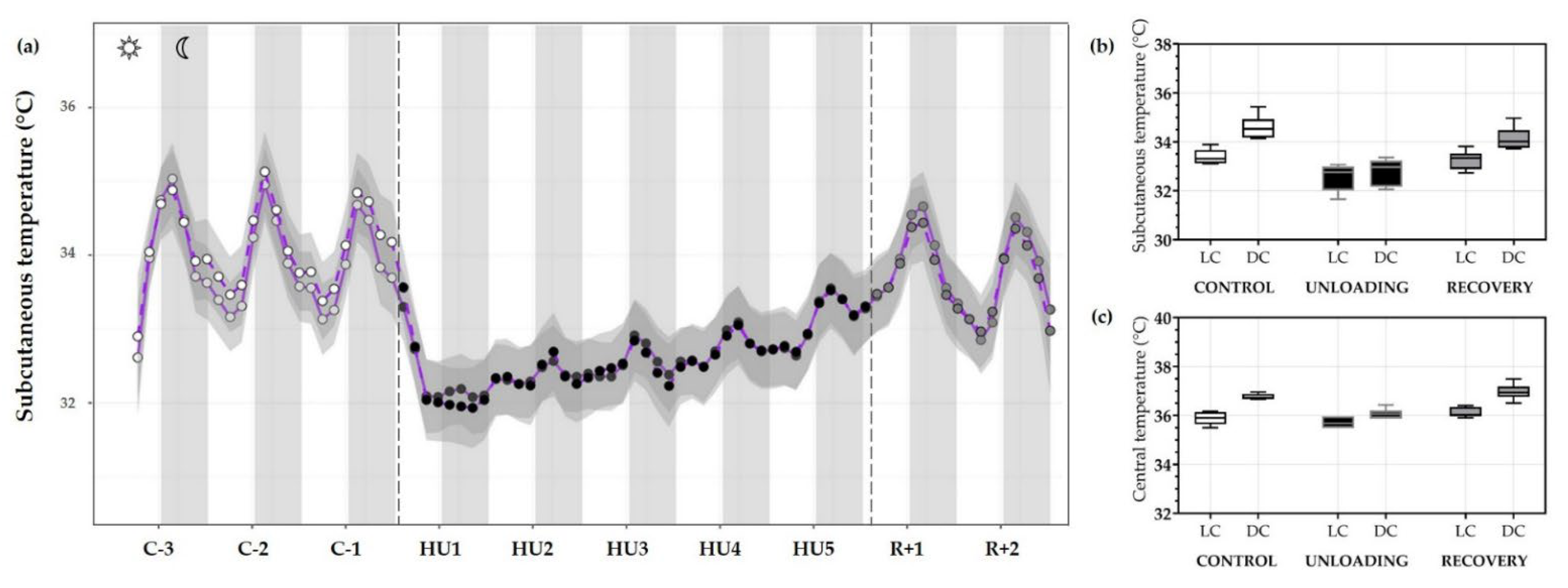
3.3.2. Temperature
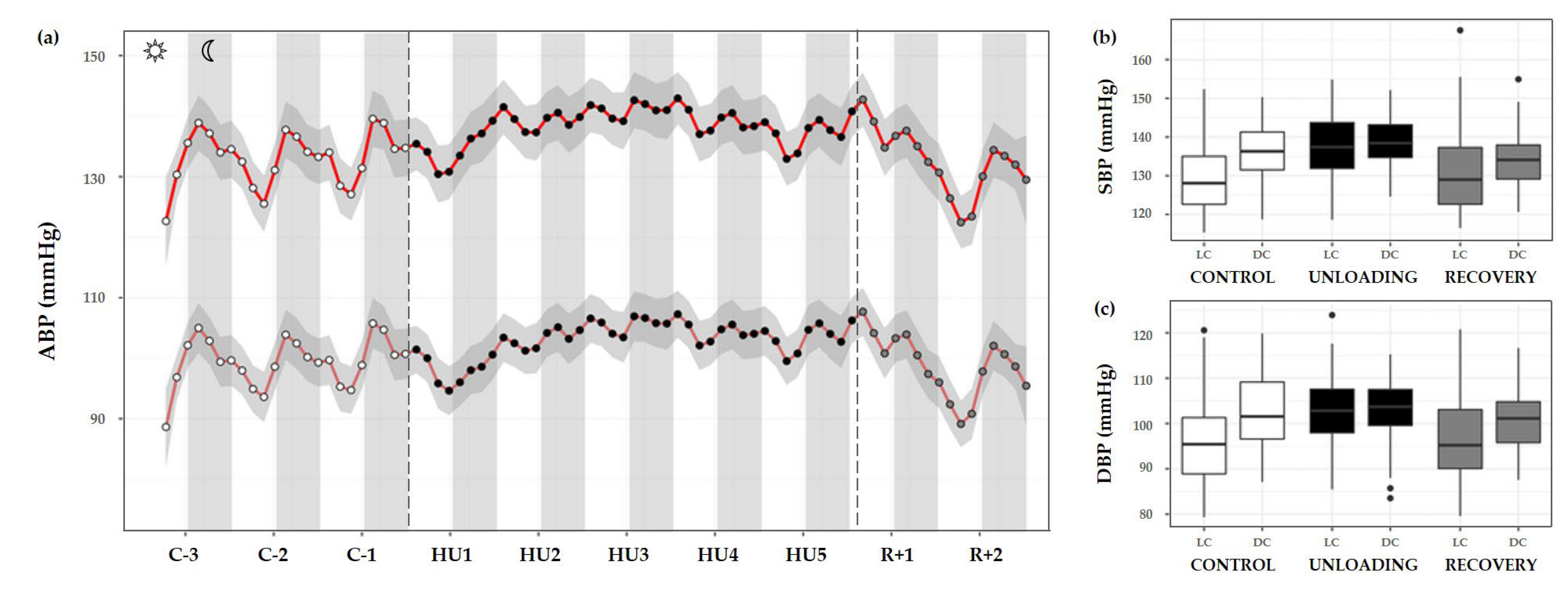
3.3.3. Arterial Blood Pressure
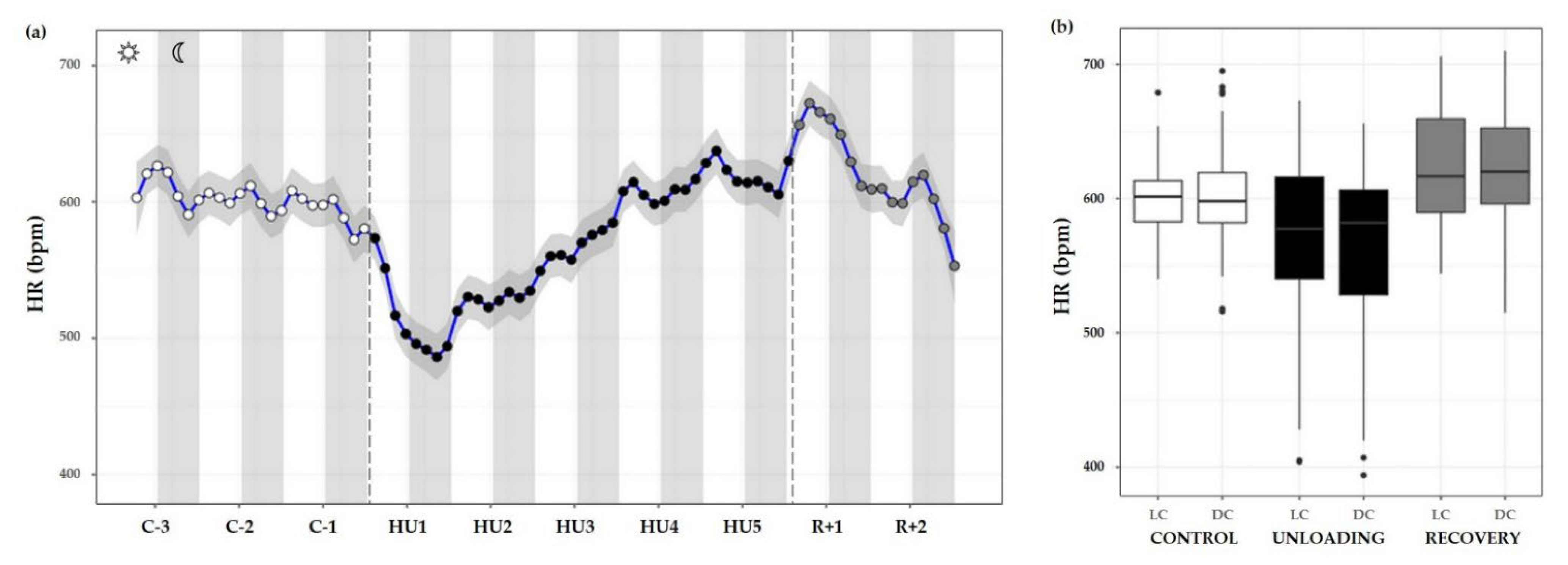
3.3.4. Heart Rate
3.3.5. Cosinor Analyses
4. Discussion
4.1. Dynamic Responses of the ANS to HU and Recovery
4.2. Light/Dark Cycles and Circadian Rhythms during HU and Recovery
4.3. Limitations of the Model and Analyses
4.4. Conclusions and Potential New Insights
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patel, Z.S.; Brunstetter, T.J.; Tarver, W.J.; Whitmire, A.M.; Zwart, S.R.; Smith, S.M.; Huff, J.L. Red Risks for a Journey to the Red Planet: The Highest Priority Human Health Risks for a Mission to Mars. NPJ Microgravity 2020, 6, 33. [Google Scholar] [CrossRef]
- Afshinnekoo, E.; Scott, R.T.; MacKay, M.J.; Pariset, E.; Cekanaviciute, E.; Barker, R.; Gilroy, S.; Hassane, D.; Smith, S.M.; Zwart, S.R.; et al. Fundamental Biological Features of Spaceflight: Advancing the Field to Enable Deep-Space Exploration. Cell 2020, 183, 1162–1184. [Google Scholar] [CrossRef]
- Ly, V.; Velichala, S.R.; Hargens, A.R. Cardiovascular, Lymphatic, and Ocular Health in Space. Life 2022, 12, 268. [Google Scholar] [CrossRef] [PubMed]
- Gallo, C.; Ridolfi, L.; Scarsoglio, S. Cardiovascular Deconditioning during Long-Term Spaceflight through Multiscale Modeling. NPJ Microgravity 2020, 6, 27. [Google Scholar] [CrossRef]
- Man, J.; Graham, T.; Squires-Donelly, G.; Laslett, A.L. The Effects of Microgravity on Bone Structure and Function. NPJ Microgravity 2022, 8, 9. [Google Scholar] [CrossRef]
- Coulombe, J.C.; Senwar, B.; Ferguson, V.L. Spaceflight-Induced Bone Tissue Changes That Affect Bone Quality and Increase Fracture Risk. Curr. Osteoporos. Rep. 2020, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Comfort, P.; McMahon, J.J.; Jones, P.A.; Cuthbert, M.; Kendall, K.; Lake, J.P.; Haff, G.G. Effects of Spaceflight on Musculoskeletal Health: A Systematic Review and Meta-Analysis, Considerations for Interplanetary Travel. Sports Med. 2021, 51, 2097–2114. [Google Scholar] [CrossRef] [PubMed]
- ElGindi, M.; Sapudom, J.; Ibrahim, I.H.; Al-Sayegh, M.; Chen, W.; Garcia-Sabaté, A.; Teo, J.C.M. May the Force Be with You (Or Not): The Immune System under Microgravity. Cells 2021, 10, 1941. [Google Scholar] [CrossRef] [PubMed]
- Brainard, G.C.; Barger, L.K.; Soler, R.R.; Hanifin, J.P. The Development of Lighting Countermeasures for Sleep Disruption and Circadian Misalignment during Spaceflight. Curr. Opin. Pulm. Med. 2016, 22, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Baran, R.; Marchal, S.; Garcia Campos, S.; Rehnberg, E.; Tabury, K.; Baselet, B.; Wehland, M.; Grimm, D.; Baatout, S. The Cardiovascular System in Space: Focus on In Vivo and In Vitro Studies. Biomedicines 2021, 10, 59. [Google Scholar] [CrossRef]
- McCrory, C.; Berkman, L.F.; Nolan, H.; O’Leary, N.; Foley, M.; Kenny, R.A. Speed of Heart Rate Recovery in Response to Orthostatic Challenge. Circ. Res. 2016, 119, 666–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandsager, K.T.; Robertson, D.; Diedrich, A. The Function of the Autonomic Nervous System during Spaceflight. Clin. Auton. Res. 2015, 25, 141–151. [Google Scholar] [CrossRef]
- Powers, J.; Bernstein, D. The Mouse as a Model of Cardiovascular Adaptations to Microgravity. J. Appl. Physiol. (1985) 2004, 97, 1686–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hargens, A.R.; Richardson, S. Cardiovascular Adaptations, Fluid Shifts, and Countermeasures Related to Space Flight. Respir. Physiol. Neurobiol. 2009, 169 (Suppl. 1), S30–S33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Hamamura, K.; Yokota, H. A Brief Review of Bone Adaptation to Unloading. Genom. Proteom. Bioinform. 2008, 6, 4–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, N.; Otsuka, K.; Kubo, Y.; Hayashi, M.; Mizuno, K.; Ohshima, H.; Mukai, C. Effects of Long-Term Microgravity Exposure in Space on Circadian Rhythms of Heart Rate Variability. Chronobiol. Int. 2015, 32, 327–340. [Google Scholar] [CrossRef]
- Chouchou, F.; Pichot, V.; Costes, F.; Guillot, M.; Barthélémy, J.-C.; Bertoletti, L.; Roche, F. Autonomic Cardiovascular Adaptations to Acute Head-out Water Immersion, Head-down Tilt and Supine Position. Eur. J. Appl. Physiol. 2020, 120, 337–347. [Google Scholar] [CrossRef]
- Pavy-Le Traon, A.; Heer, M.; Narici, M.V.; Rittweger, J.; Vernikos, J. From Space to Earth: Advances in Human Physiology from 20 Years of Bed Rest Studies (1986–2006). Eur. J. Appl. Physiol. 2007, 101, 143–194. [Google Scholar] [CrossRef]
- Pandiarajan, M.; Hargens, A.R. Ground-Based Analogs for Human Spaceflight. Front. Physiol. 2020, 11, 716. [Google Scholar] [CrossRef] [PubMed]
- Globus, R.K.; Morey-Holton, E. Hindlimb Unloading: Rodent Analog for Microgravity. J. Appl. Physiol. (1985) 2016, 120, 1196–1206. [Google Scholar] [CrossRef] [PubMed]
- Hawliczek, A.; Brix, B.; Al Mutawa, S.; Alsuwaidi, H.; Du Plessis, S.; Gao, Y.; Qaisar, R.; Siddiqui, R.; Elmoselhi, A.B.; Goswami, N. Hind-Limb Unloading in Rodents: Current Evidence and Perspectives. Acta Astronaut. 2022, 195, 574–582. [Google Scholar] [CrossRef]
- Tsvirkun, D.; Bourreau, J.; Mieuset, A.; Garo, F.; Vinogradova, O.; Larina, I.; Navasiolava, N.; Gauquelin-Koch, G.; Gharib, C.; Custaud, M.-A. Contribution of Social Isolation, Restraint, and Hindlimb Unloading to Changes in Hemodynamic Parameters and Motion Activity in Rats. PLoS ONE 2012, 7, e39923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasser, E.M.; Moffitt, J.A. Regulation of Sympathetic Nervous System Function after Cardiovascular Deconditioning. Ann. N. Y. Acad. Sci. 2001, 940, 454–468. [Google Scholar] [CrossRef]
- Mueller, P.J.; Cunningham, J.T.; Patel, K.P.; Hasser, E.M. Proposed Role of the Paraventricular Nucleus in Cardiovascular Deconditioning. Acta Physiol. Scand. 2003, 177, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb Unloading Rodent Model: Technical Aspects. J. Appl. Physiol. (1985) 2002, 92, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Garg, P.; Strigini, M.; Peurière, L.; Vico, L.; Iandolo, D. The Skeletal Cellular and Molecular Underpinning of the Murine Hindlimb Unloading Model. Front. Physiol. 2021, 12, 749464. [Google Scholar] [CrossRef]
- van den Buuse, M.; Malpas, S.C. 24-Hour Recordings of Blood Pressure, Heart Rate and Behavioural Activity in Rabbits by Radio-Telemetry: Effects of Feeding and Hypertension. Physiol. Behav. 1997, 62, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Thireau, J.; Zhang, B.L.; Poisson, D.; Babuty, D. Heart Rate Variability in Mice: A Theoretical and Practical Guide: Heart Rate Variability in Mice. Exp. Physiol. 2008, 93, 83–94. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2; Springer: New York, NY, USA, 2009; ISBN 978-0-387-98140-6. [Google Scholar]
- Cleveland, W.S.; Devlin, S.J. Locally Weighted Regression: An Approach to Regression Analysis by Local Fitting. J. Am. Stat. Assoc. 1988, 83, 596–610. [Google Scholar] [CrossRef]
- Cornelissen, G. Cosinor-Based Rhythmometry. Theor. Biol. Med. Model. 2014, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- Mutak, A. Cosinor2: Extended Tools for Cosinor Analysis of Rhythms 2018. R Package Version 0.2.1. Available online: https://CRAN.R-project.org/package=cosinor2 (accessed on 16 March 2023).
- Sachs, M. Cosinor: Tools for Estimating and Predicting the Cosinor Model 2022. R Package Version 1.2.2. Available online: https://cran.r-project.org/web/packages/cosinor/index.html (accessed on 16 March 2023).
- Shaffer, F.; Ginsberg, J.P. An Overview of Heart Rate Variability Metrics and Norms. Front. Public Health 2017, 5, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauquelin, G.; Kazek, C.; Allevard, A.-M.; Garcin, R.; Bonnod, J.; Gutkowska, J.; Cantin, M.; Gharib, C. Early (1 to 24h) Plasma Atrial Natriuretic Factor Changes in the Rat during Antiorthostatic Hypokinetic Syspension. Biochem. Biophys. Res. Commun. 1987, 148, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Antunes-Rodrigues, J.; Machado, B.H.; Andrade, H.A.; Mauad, H.; Ramalho, M.J.; Reis, L.C.; Silva-Netto, C.R.; Favaretto, A.L.; Gutkowska, J.; McCann, S.M. Carotid-Aortic and Renal Baroreceptors Mediate the Atrial Natriuretic Peptide Release Induced by Blood Volume Expansion. Proc. Natl. Acad. Sci. USA 1992, 89, 6828–6831. [Google Scholar] [CrossRef] [Green Version]
- Shellock, F.G.; Swan, H.J.; Rubin, S.A. Early Central Venous Pressure Changes in the Rat during Two Different Levels of Head-down Suspension. Aviat. Space Environ. Med. 1985, 56, 791–795. [Google Scholar] [PubMed]
- Moffitt, J.A.; Grippo, A.J.; Beltz, T.G.; Johnson, A.K. Hindlimb Unloading Elicits Anhedonia and Sympathovagal Imbalance. J. Appl. Physiol. (1985) 2008, 105, 1049–1059. [Google Scholar] [CrossRef] [Green Version]
- Moffitt, J.A.; Henry, M.K.; Welliver, K.C.; Jepson, A.J.; Garnett, E.R. Hindlimb Unloading Results in Increased Predisposition to Cardiac Arrhythmias and Alters Left Ventricular Connexin 43 Expression. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R362–R373. [Google Scholar] [CrossRef]
- Greening, G.J.; Miller, K.P.; Spainhour, C.R.; Cato, M.D.; Muldoon, T.J. Effects of Isoflurane Anesthesia on Physiological Parameters in Murine Subcutaneous Tumor Allografts Measured via Diffuse Reflectance Spectroscopy. Biomed. Opt. Express BOE 2018, 9, 2871–2886. [Google Scholar] [CrossRef] [PubMed]
- Rose, A.; Steffen, J.M.; Musacchia, X.J.; Mandel, A.D.; Sonnenfeld, G. Effect of Antiorthostatic Suspension on Interferon-Alpha/Beta Production by the Mouse. Proc. Soc. Exp. Biol. Med. 1984, 177, 253–256. [Google Scholar] [CrossRef]
- Musacchia, X.J.; Fagette, S. Weightlessness Simulations for Cardiovascular and Muscle Systems: Validity of Rat Models. J. Gravit. Physiol. 1997, 4, 49–59. [Google Scholar] [PubMed]
- Foley, C.M.; Mueller, P.J.; Hasser, E.M.; Heesch, C.M. Hindlimb Unloading and Female Gender Attenuate Baroreflex-Mediated Sympathoexcitation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R1440–R1447. [Google Scholar] [CrossRef] [Green Version]
- Mueller, P.J.; Sullivan, M.J.; Grindstaff, R.R.; Cunningham, J.T.; Hasser, E.M. Regulation of Plasma Vasopressin and Renin Activity in Conscious Hindlimb-Unloaded Rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R46–R52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagette, S.; Lo, M.; Gharib, C.; Gauquelin, G. Cardiovascular Variability and Baroreceptor Reflex Sensitivity over a 14-Day Tail Suspension in Rats. J. Appl. Physiol. (1985) 1995, 78, 717–724. [Google Scholar] [CrossRef]
- Gordon, C.J. The Mouse Thermoregulatory System: Its Impact on Translating Biomedical Data to Humans. Physiol. Behav. 2017, 179, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Ripperger, J.A.; Jud, C.; Albrecht, U. The Daily Rhythm of Mice. FEBS Lett. 2011, 585, 1384–1392. [Google Scholar] [CrossRef] [Green Version]
- Shimatani, H.; Inoue, Y.; Maekawa, Y.; Miyake, T.; Yamaguchi, Y.; Doi, M. Thermographic Imaging of Mouse across Circadian Time Reveals Body Surface Temperature Elevation Associated with Non-Locomotor Body Movements. PLoS ONE 2021, 16, e0252447. [Google Scholar] [CrossRef] [PubMed]
- Gaskill, B.N.; Gordon, C.J.; Pajor, E.A.; Lucas, J.R.; Davis, J.K.; Garner, J.P. Impact of Nesting Material on Mouse Body Temperature and Physiology. Physiol. Behav. 2013, 110–111, 87–95. [Google Scholar] [CrossRef]
- Stahn, A.C.; Werner, A.; Opatz, O.; Maggioni, M.A.; Steinach, M.; von Ahlefeld, V.W.; Moore, A.; Crucian, B.E.; Smith, S.M.; Zwart, S.R.; et al. Increased Core Body Temperature in Astronauts during Long-Duration Space Missions. Sci. Rep. 2017, 7, 16180. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, R. Regulation of Circadian Blood Pressure—From Mice to Astronauts. Curr. Opin. Nephrol. Hypertens. 2010, 19, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Makino, M.; Hayashi, H.; Takezawa, H.; Hirai, M.; Saito, H.; Ebihara, S. Circadian Rhythms of Cardiovascular Functions Are Modulated by the Baroreflex and the Autonomic Nervous System in the Rat. Circulation 1997, 96, 1667–1674. [Google Scholar] [CrossRef]
- Tahimic, C.G.T.; Paul, A.M.; Schreurs, A.-S.; Torres, S.M.; Rubinstein, L.; Steczina, S.; Lowe, M.; Bhattacharya, S.; Alwood, J.S.; Ronca, A.E.; et al. Influence of Social Isolation During Prolonged Simulated Weightlessness by Hindlimb Unloading. Front. Physiol. 2019, 10, 1147. [Google Scholar] [CrossRef] [Green Version]
- Zhai, B.; Fu, J.; Xiang, S.; Shang, Y.; Yan, Y.; Yin, T.; Zhang, T. Repetitive Transcranial Magnetic Stimulation Ameliorates Recognition Memory Impairment Induced by Hindlimb Unloading in Mice Associated with BDNF/TrkB Signaling. Neurosci. Res. 2020, 153, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Dikareva, A.; Bacon, S.L.; Daskalopoulou, S.S. The Impact of Physical Activity on Mortality in Patients with High Blood Pressure: A Systematic Review. J. Hypertens. 2012, 30, 1277–1288. [Google Scholar] [CrossRef] [Green Version]
- Reimers, A.K.; Knapp, G.; Reimers, C.-D. Effects of Exercise on the Resting Heart Rate: A Systematic Review and Meta-Analysis of Interventional Studies. J. Clin. Med. 2018, 7, 503. [Google Scholar] [CrossRef] [Green Version]
- Qaisar, R.; Karim, A.; Elmoselhi, A.B. Muscle unoading: A comparison between spaceflight and ground-based models. Acta Physiol. 2020, 228, e13431. [Google Scholar] [CrossRef] [PubMed]
- Tabibzadeh, S. CircadiOmic Medicine and Aging. Ageing Res. Rev. 2021, 71, 101424. [Google Scholar] [CrossRef]
- Rutovskaya, M.V.; Kosyreva, A.M.; Diatroptov, M.E. Ultradian and Infradian Rhythms in the Dynamic of Testosterone Concentration in the Serum of the White-Breasted Hedgehog Erinaceus Roumanicus. Sci. Rep. 2020, 10, 6334. [Google Scholar] [CrossRef] [Green Version]
- Coyne, M.D.; Kesick, C.M.; Doherty, T.J.; Kolka, M.A.; Stephenson, L.A. Circadian Rhythm Changes in Core Temperature over the Menstrual Cycle: Method for Noninvasive Monitoring. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R1316–R1320. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, A.S.; Hoffmann, H.M. Editorial: Hormone Release Patterns in Mammals. Mol. Cell Endocrinol. 2020, 507, 110781. [Google Scholar] [CrossRef]
- Blum, I.D.; Zhu, L.; Moquin, L.; Kokoeva, M.V.; Gratton, A.; Giros, B.; Storch, K.-F. A Highly Tunable Dopaminergic Oscillator Generates Ultradian Rhythms of Behavioral Arousal. eLife 2014, 3, e05105. [Google Scholar] [CrossRef]
- Baik, J.-H. Stress and the Dopaminergic Reward System. Exp. Mol. Med. 2020, 52, 1879–1890. [Google Scholar] [CrossRef] [PubMed]
- Popova, N.K.; Kulikov, A.V.; Kondaurova, E.M.; Tsybko, A.S.; Kulikova, E.A.; Krasnov, I.B.; Shenkman, B.S.; Bazhenova, E.Y.; Sinyakova, N.A.; Naumenko, V.S. Risk Neurogenes for Long-Term Spaceflight: Dopamine and Serotonin Brain System. Mol. Neurobiol. 2015, 51, 1443–1451. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, K.; Izumi, R.; Ishioka, N.; Ohshima, H.; Mukai, C. Chronomics of Heart Rate Variability on Earth and in Space. Respir. Physiol. Neurobiol. 2009, 169 (Suppl. 1), S69–S72. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | |||
|---|---|---|---|
| Control | Unloading | Recovery | |
| Locomotor activity (Counts) | |||
| MESOR | 0.26 ± 0.11 | 0.04 ± 0.01 *** | 0.21 ± 0.05 ### |
| Acrophase | −1.10 ± 0.82 | −0.64 ± 0.68 | 0.95 ± 1.00 *** ## |
| Amplitude | 0.18 ± 0.11 | 0.01 ± 0.01 *** | 0.13 ± 0.08 ## |
| Rhythm detection test p-value | 0.005 * | 0.420 | 0.070 |
| Subcutaneous temperature (°C) | |||
| MESOR | 33.81 ± 0.72 | 32.69 ± 0.51 *** | 33.73 ± 0.40 ## |
| Acrophase | −0.69 ± 0.25 | −0.72 ± 1.01 | 0.18 ± 1.36 |
| Amplitude | 0.78 ± 0.19 | 0.23 ± 0.07 *** | 0.71 ± 0.17 ### |
| Rhythm detection test p-value | 0.004 * | 0.129 | 0.026 * |
| Systolic blood pressure (mmHg) | |||
| MESOR | 128.3 ± 14.4 | 134.4 ± 14.9 * | 129.0 ± 12.8 |
| Acrophase | −0.76 ± 0.96 | −0.41 ± 0.39 | −0.26 ± 1.06 |
| Amplitude | 6.32 ± 2.74 | 2.66 ± 1.28 *** | 3.39 ± 1.89 ** |
| Rhythm detection test p-value | 0.118 | 0.192 | 0.486 |
| Diastolic blood pressure (mmHg) | |||
| MESOR | 102.3 ± 2.8 | 106.4 ± 7.1 | 101.8 ± 8.6 # |
| Acrophase | −1.18 ± 0.14 | −0.40 ± 0.64 | −0.05 ± 1.25 * |
| Amplitude | 5.37 ± 2.53 | 2.09 ± 1.10 ** | 2.28 ± 4.95 |
| Rhythm detection test p-value | 0.104 | 0.316 | 0.372 |
| Heart rate (bpm) | |||
| MESOR | 590.6 ± 14.5 | 569.9 ± 22.9 * | 616.6 ± 22.2 * ### |
| Acrophase | 0.30 ± 0.87 | 0.17 ± 0.97 | −0.09 ± 0.66 |
| Amplitude | 18.23 ± 8.29 | 8.57 ± 6.01 | 16.47 ± 10.46 |
| Rhythm detection test p-value | 0.170 | 0.562 | 0.486 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hélissen, O.; Kermorgant, M.; Déjean, S.; Mercadie, A.; Le Gonidec, S.; Zahreddine, R.; Calise, D.; Nasr, N.; Galès, C.; Arvanitis, D.N.; et al. Autonomic Nervous System Adaptation and Circadian Rhythm Disturbances of the Cardiovascular System in a Ground-Based Murine Model of Spaceflight. Life 2023, 13, 844. https://doi.org/10.3390/life13030844
Hélissen O, Kermorgant M, Déjean S, Mercadie A, Le Gonidec S, Zahreddine R, Calise D, Nasr N, Galès C, Arvanitis DN, et al. Autonomic Nervous System Adaptation and Circadian Rhythm Disturbances of the Cardiovascular System in a Ground-Based Murine Model of Spaceflight. Life. 2023; 13(3):844. https://doi.org/10.3390/life13030844
Chicago/Turabian StyleHélissen, Ophélie, Marc Kermorgant, Sébastien Déjean, Aurélie Mercadie, Sophie Le Gonidec, Rana Zahreddine, Denis Calise, Nathalie Nasr, Céline Galès, Dina N. Arvanitis, and et al. 2023. "Autonomic Nervous System Adaptation and Circadian Rhythm Disturbances of the Cardiovascular System in a Ground-Based Murine Model of Spaceflight" Life 13, no. 3: 844. https://doi.org/10.3390/life13030844