

The Abundant Phytocannabinoids in Rheumatoid Arthritis: Therapeutic Targets and Molecular Processes Identified Using Integrated Bioinformatics and Network Pharmacology



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
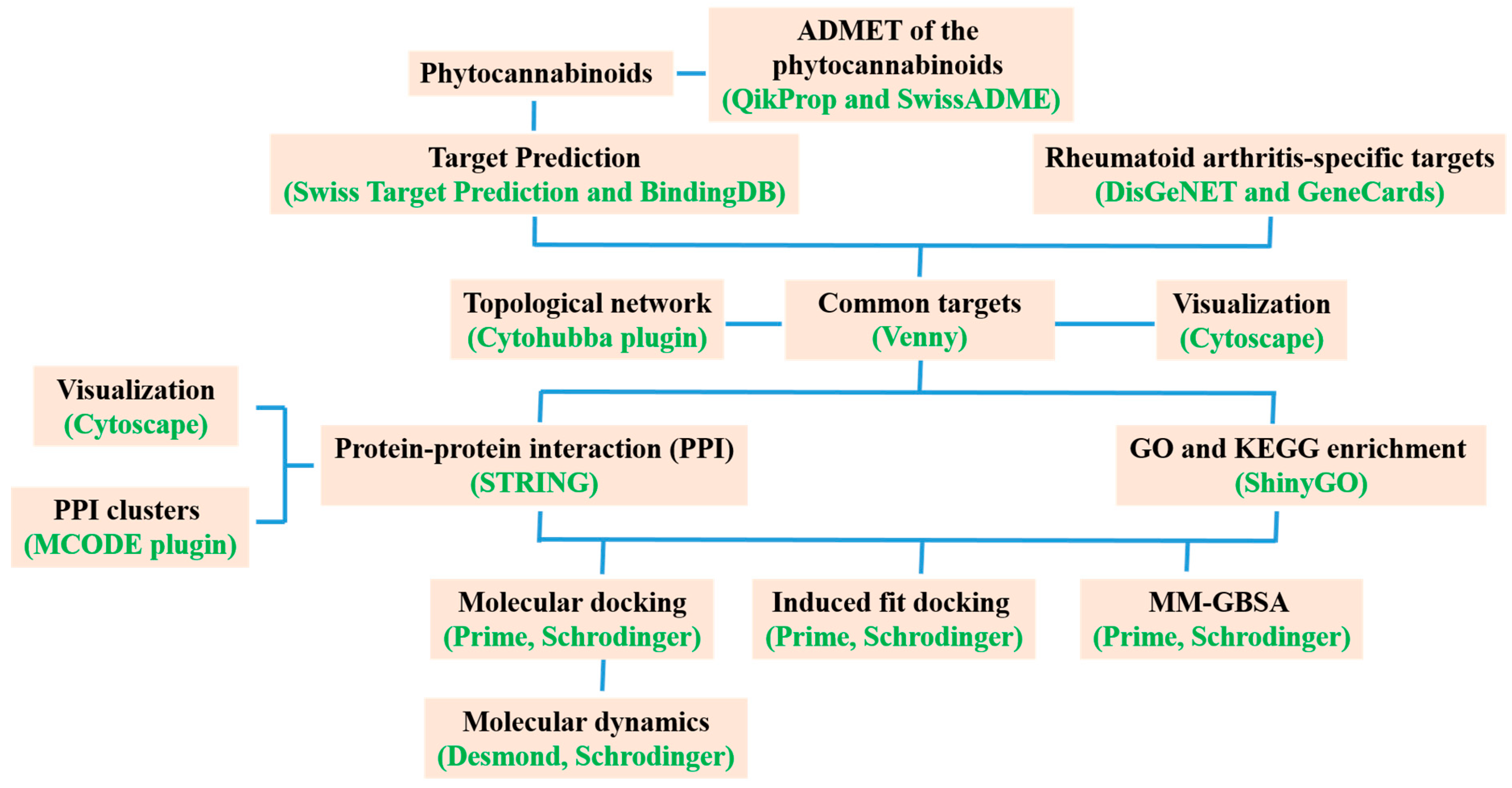
2. Materials and Methods
2.1. Identification of Bioactive Compounds and Targets
2.2. Collection of Protein–Protein Interaction (PPI) Data
2.3. GO and KEGG Enrichment Analyses
2.4. Molecular Docking Validation
2.5. Molecular Dynamics (MD) Simulation
2.6. Induced-Fit Docking (IFD)
2.7. Molecular Mechanics-Generalized Born Surface Area (MM-GBSA) Calculation
2.8. Physicochemical and ADME/T Studies
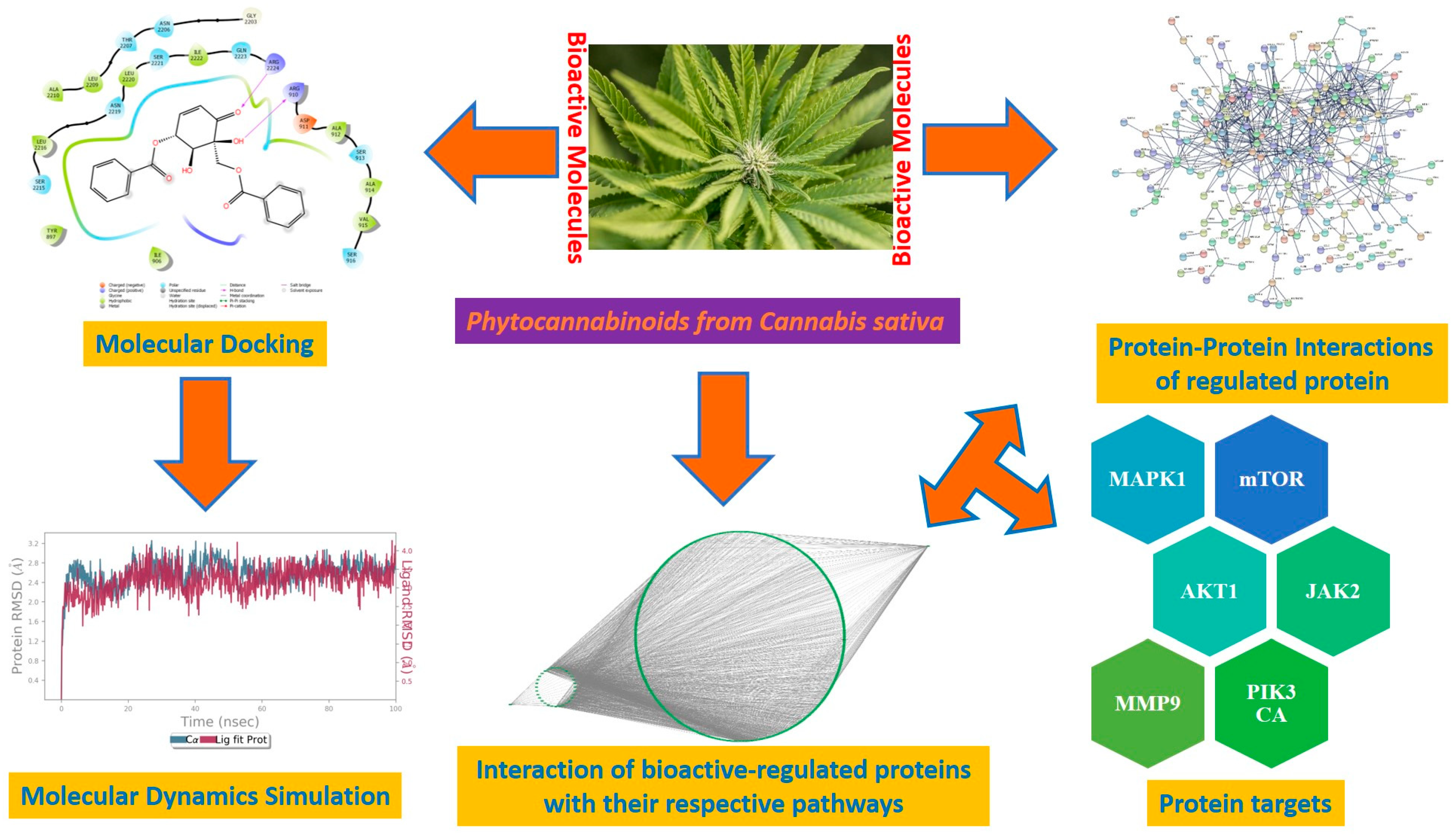
3. Results
3.1. Identification of Bioactive Compounds and Targets
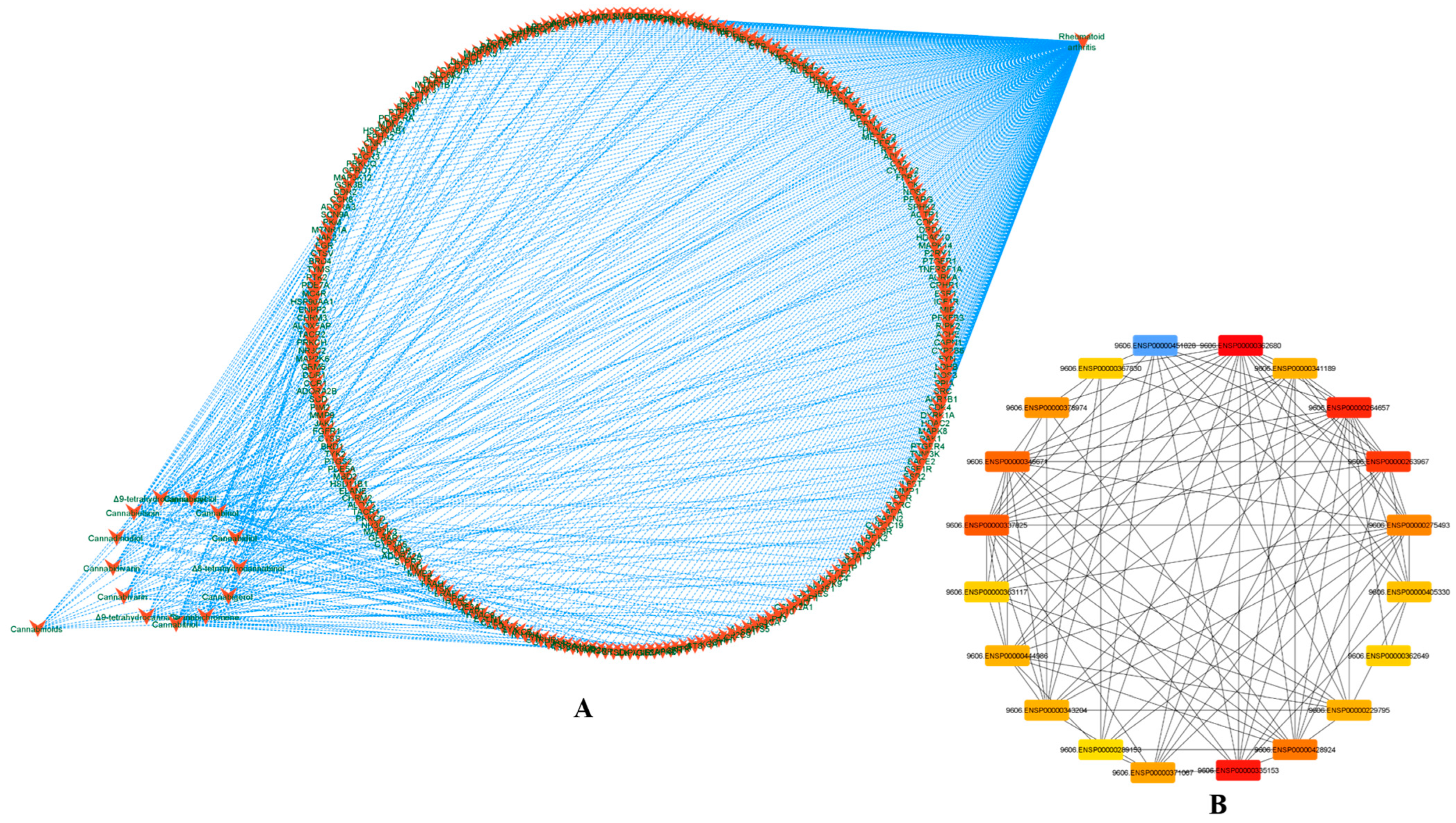
3.2. Phytocannabinoids–Rheumatoid Arthritis Target and PPI Network Analysis
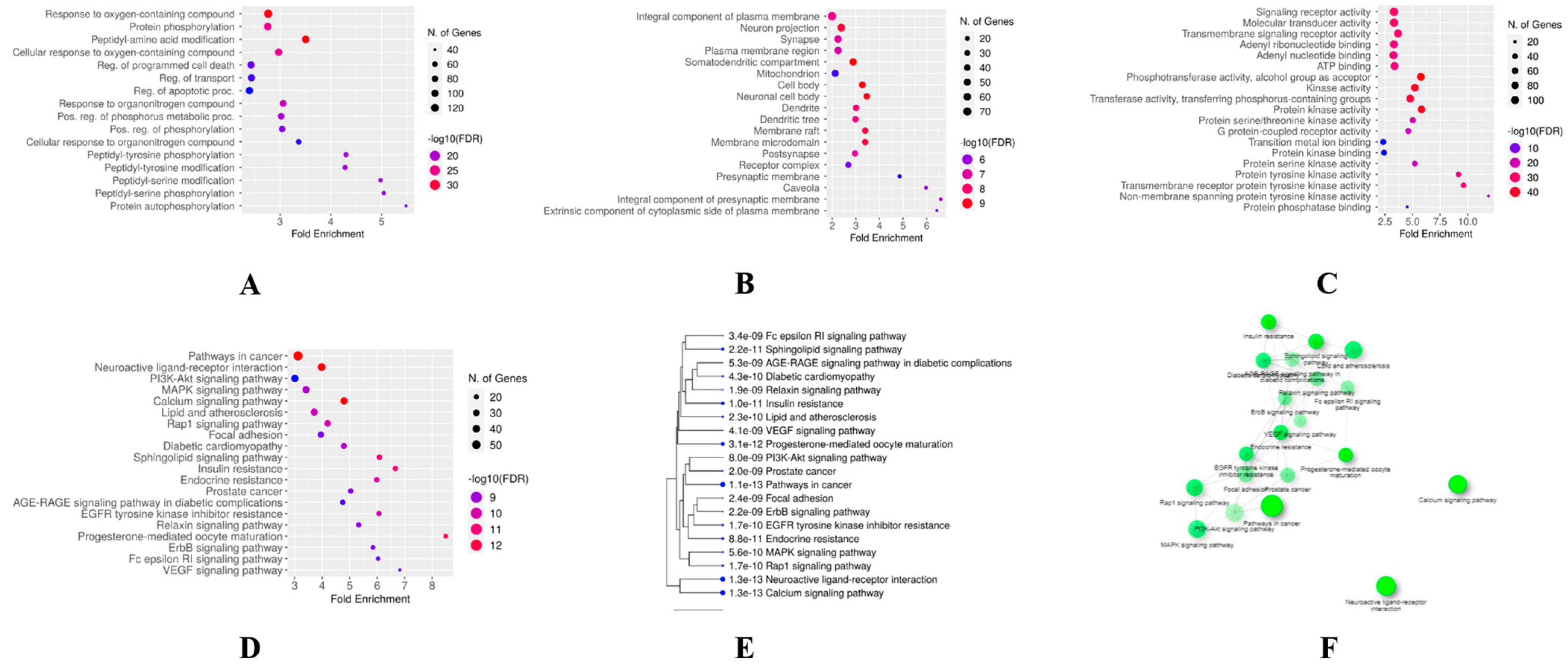
3.3. GO and KEGG Enrichment Analyses
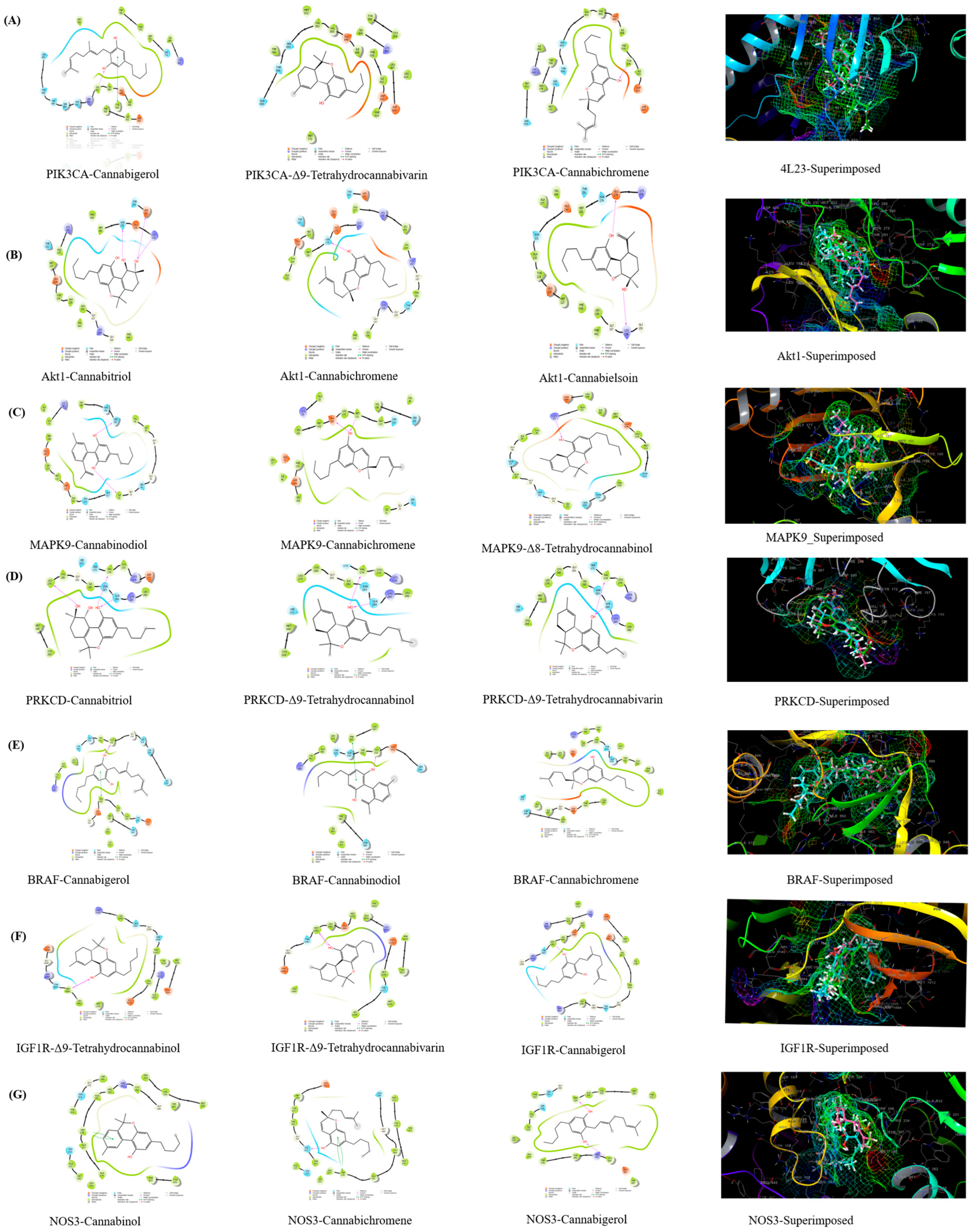
3.4. Molecular Docking Validation
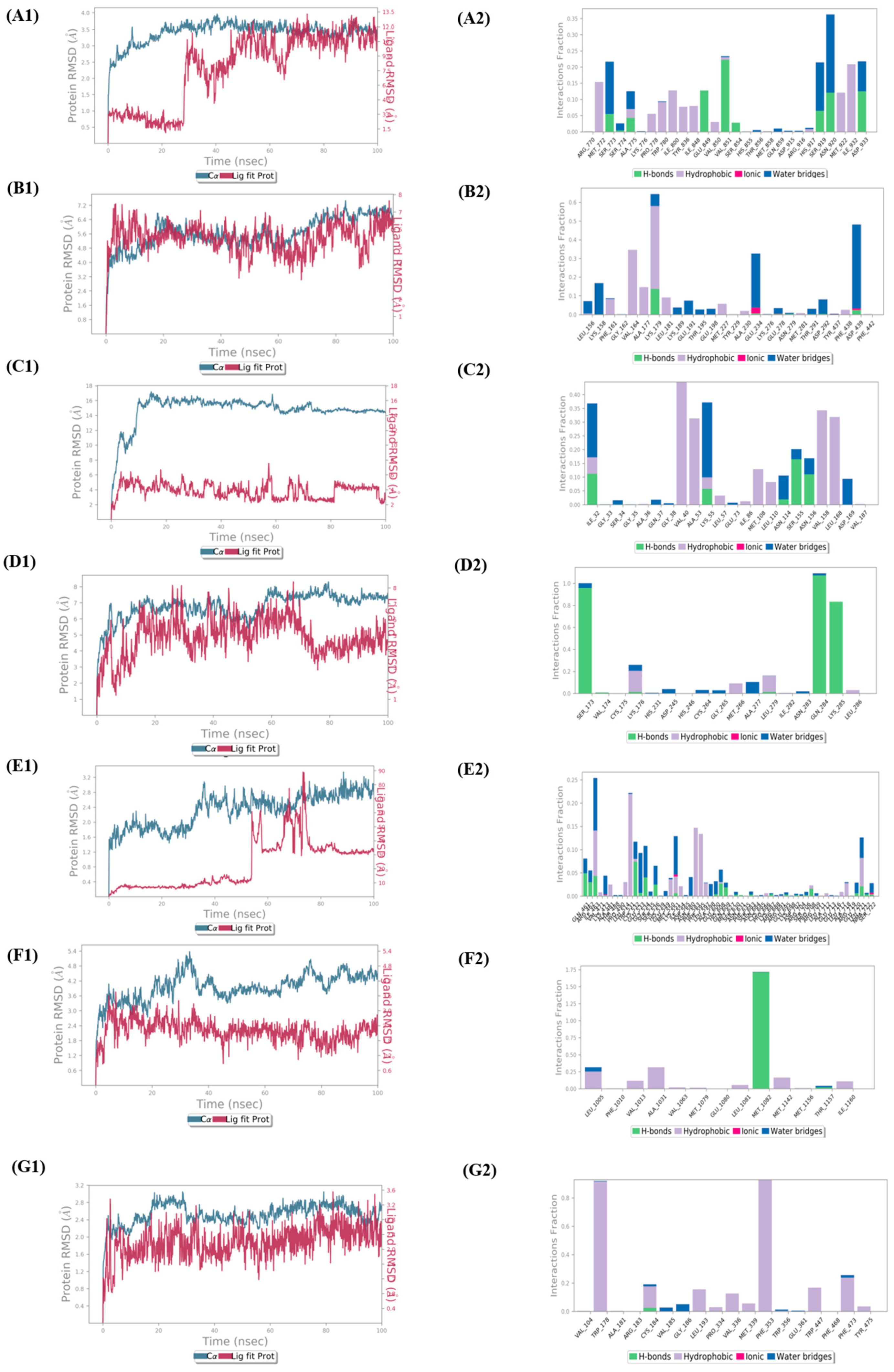
3.5. Molecular Dynamics
3.6. Induced-Fit Docking and MM-GBSA Calculation
3.7. Physicochemical and ADME/T Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADMET | Absorption, distribution, metabolism, excretion, and toxicity |
| BBB | Blood-brain barrier |
| CB | Cannabinoid receptor |
| CVD | Cardiovascular disease |
| GO | Gene Ontology |
| IFD | Induced Fit docking |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| MCODE | Molecular Complex Detection |
| MM-GBSA | Molecular mechanics-generalized Born surface area |
| NOS | Nitric oxide synthetase |
| NSAIDs | Non-steroidal anti-inflammatory drugs |
| PPI | Protein-protein interaction |
| PDB | Protein data bank |
| PGP | P-glycoprotein |
| RA | Rheumatoid arthritis |
| RMSD | Root mean square deviation |
| RMSF | Root mean square fluctuation |
| SPC | Simple point charge |
| THC | Tetrahydrocannabinoids |
| vdW | van der Waals |
References
- Yeşilada, E.; Üstün, O.; Sezik, E.; Takaishi, Y.; Ono, Y.; Honda, G. Inhibitory Effects of Turkish Folk Remedies on Inflammatory Cytokines: Interleukin-1α, Interleukin-1β and Tumor Necrosis Factor α. J. Ethnopharmacol. 1997, 58, 59–73. [Google Scholar] [CrossRef]
- Butler, S.H.; Godefroy, F.; Besson, J.M.; Weil-Fugazza, J. A Limited Arthritic Model for Chronic Pain Studies in the Rat. Pain 1992, 48, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Corley, D.A.; Kerlikowse, K.; Verma, R.; Buffler, P. Protective Association of Aspirin/NSAIDs and Esophageal Cancer: A Systematic Review and Meta-Analysis. Gastroenterology 2003, 124, 47–56. [Google Scholar] [CrossRef]
- Scott, C.; Neira Agonh, D.; Lehmann, C. Antibacterial Effects of Phytocannabinoids. Life 2022, 12, 1394. [Google Scholar] [CrossRef]
- Haddad, F.; Dokmak, G.; Karaman, R. The Efficacy of Cannabis on Multiple Sclerosis-Related Symptoms. Life 2022, 12, 682. [Google Scholar] [CrossRef] [PubMed]
- Calapai, F.; Cardia, L.; Calapai, G.; Di Mauro, D.; Trimarchi, F.; Ammendolia, I.; Mannucci, C. Effects of Cannabidiol on Locomotor Activity. Life 2022, 12, 652. [Google Scholar] [CrossRef]
- Mechoulam, R. The Pharmacohistory of Cannabis sativa. In Cannabinoids as Therapeutic Agents; Chapman and Hall/CRC: Boca Raton, FL, USA, 2019; pp. 1–20. ISBN 9780429260667. [Google Scholar]
- Gaoni, Y.; Mechoulam, R. Isolation, Structure, and Partial Synthesis of an Active Constituent of Hashish. J. Am. Chem. Soc. 1964, 86, 1646–1647. [Google Scholar] [CrossRef]
- Hussain, T.; Jeena, G.; Pitakbut, T.; Vasilev, N.; Kayser, O. Cannabis Sativa Research Trends, Challenges, and New-Age Perspectives. iScience 2021, 24, 103391. [Google Scholar] [CrossRef]
- Hawley, P.; Gobbo, M.; Afghari, N. The Impact of Legalization of Access to Recreational Cannabis on Canadian Medical Users with Cancer. BMC Health Serv. Res. 2020, 20, 977. [Google Scholar] [CrossRef]
- Citation, S. Marijuana and Medicine; National Academy Press: Washington, DC, USA, 1999; ISBN 9780309071550. [Google Scholar]
- Straus, S.E. Immunoactive Cannabinoids: Therapeutic Prospects for Marijuana Constituents. Proc. Natl. Acad. Sci. USA 2000, 97, 9363–9364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ElSohly, M.A.; Gul, W. Constituents of Cannabis sativa. In Handbook of Cannabis; Oxford University Press: Oxford, UK, 2014; pp. 3–22. [Google Scholar]
- Morales, P.; Hurst, D.P.; Reggio, P.H. Molecular Targets of the Phytocannabinoids: A Complex Picture. In Progress in the Chemistry of Organic Natural Products; Springer: Cham, Switzerland, 2017; Volume 103, pp. 103–131. [Google Scholar]
- Scott, D.L.; Wolfe, F.; Huizinga, T.W.J. Rheumatoid Arthritis. Lancet 2010, 376, 1094–1108. [Google Scholar] [CrossRef]
- Gui, H.; Tong, Q.; Qu, W.; Mao, C.M.; Dai, S.M. The Endocannabinoid System and Its Therapeutic Implications in Rheumatoid Arthritis. Int. Immunopharmacol. 2015, 26, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef]
- Khanal, P.; Duyu, T.; Patil, B.M.; Dey, Y.N.; Pasha, I.; Wanjari, M.; Gurav, S.S.; Maity, A. Network pharmacology of AYUSH recommended immune-boosting medicinal plants against COVID-19. J. Ayurveda Integr. Med. 2022, 13, 100374. [Google Scholar] [CrossRef] [PubMed]
- Khanal, P.; Zargari, F.; Far, B.F.; Kumar, D.; Morgana, R.; Mahdi, Y.K.; Jubair, N.K.; Saraf, S.K.; Bansal, P.; Singh, R.; et al. Integration of System Biology Tools to Investigate Huperzine A as an anti-alzheimer agent. Front. Pharmacol. 2021, 12, 785964. [Google Scholar] [CrossRef] [PubMed]
- Duyu, T.; Khanal, P.; Dey, Y.N.; Jha, S. Network pharmacology of Withania somnifera against stress associated neurodegenerative diseases. Adv. Tradit. Med. 2021, 21, 565–578. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, J.; Zhao, X.; Zhao, Y.; Zhu, S. Network Pharmacology and Molecular Docking Study on the Active Ingredients of Qidengmingmu Capsule for the Treatment of Diabetic Retinopathy. Sci. Rep. 2021, 11, 7382. [Google Scholar] [CrossRef]
- Song, S.; Zhou, J.; Li, Y.; Liu, J.; Li, J.; Shu, P. Network Pharmacology and Experimental Verification Based Research into the Effect and Mechanism of Aucklandiae Radix–Amomi Fructus against Gastric Cancer. Sci. Rep. 2022, 12, 9401. [Google Scholar] [CrossRef]
- Hong, Z.; Zhang, T.; Zhang, Y.; Xie, Z.; Lu, Y.; Yao, Y.; Yang, Y.; Wu, H.; Liu, B. Reveals of Candidate Active Ingredients in Justicia and Its Anti-Thrombotic Action of Mechanism Based on Network Pharmacology Approach and Experimental Validation. Sci. Rep. 2021, 11, 17187. [Google Scholar] [CrossRef]
- Abdizadeh, R.; Heidarian, E.; Hadizadeh, F.; Abdizadeh, T. Investigation of Pyrimidine Analogues as Xanthine Oxidase Inhibitors to Treat of Hyperuricemia and Gout through Combined QSAR Techniques, Molecular Docking and Molecular Dynamics Simulations. J. Taiwan Inst. Chem. Eng. 2020, 113, 72–100. [Google Scholar] [CrossRef]
- Wang, J.; Li, B.; Zhang, X.; Hu, Q.; Yu, W.; Wang, H.; Duan, D.; Li, J.; Zhao, B. Docking and Molecular Dynamics Studies on the Mechanism of Phospholipase D-Mediated Transphosphatidylation to Construct the Reaction Kinetic Model: Application in Phosphatidylserine Production. J. Taiwan Inst. Chem. Eng. 2019, 96, 82–92. [Google Scholar] [CrossRef]
- Bowers, K.J.; Chow, D.E.; Xu, H.; Dror, R.O.; Eastwood, M.P.; Gregersen, B.A.; Klepeis, J.L.; Kolossvary, I.; Moraes, M.A.; Sacerdoti, F.D.; et al. Scalable Algorithms for Molecular Dynamics Simulations on Commodity Clusters. In Proceedings of the SC ’06: Proceedings of the 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 28 February 2007; Institute of Electrical and Electronics Engineers (IEEE): New York, NY, USA; p. 43. [Google Scholar]
- Auti, P.S.; Nandi, A.; Kumari, V.; Paul, A.T. Design, Synthesis, Biological Evaluation and Molecular Modelling Studies of Oxoacetamide Warhead Containing Indole-Quinazolinone Based Novel Hybrid Analogues as Potential Pancreatic Lipase Inhibitors. New J. Chem. 2022, 46, 11648–11661. [Google Scholar] [CrossRef]
- Chattaraj, B.; Nandi, A.; Das, A.; Sharma, A.; Dey, Y.N.; Kumar, D.; Mogana, R. Inhibitory activity of Enhydra fluctuans lour. on calcium oxalate crystallisation through in silico and in vitro studies. Front. Pharmacol. 2023, 13, 5495. [Google Scholar] [CrossRef] [PubMed]
- Tutone, M.; Pibiri, I.; Lentini, L.; Pace, A.; Almerico, A.M. Deciphering the Nonsense Readthrough Mechanism of Action of Ataluren: An in Silico Compared Study. ACS Med. Chem. Lett. 2019, 10, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA Methods to Estimate Ligand-Binding Affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef]
- QikProp|Schrödinger. Available online: https://www.schrodinger.com/products/qikprop (accessed on 19 August 2022).
- Jacobson, M.P.; Pincus, D.L.; Rapp, C.S.; Day, T.J.F.; Honig, B.; Shaw, D.E.; Friesner, R.A. A Hierarchical Approach to All-Atom Protein Loop Prediction. Proteins Struct. Funct. Genet. 2004, 55, 351–367. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malemud, C.J. The PI3K/Akt/PTEN/MTOR Pathway: A Fruitful Target for Inducing Cell Death in Rheumatoid Arthritis? Future Med. Chem. 2015, 7, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Ozaita, A.; Puighermanal, E.; Maldonado, R. Regulation of PI3K/Akt/GSK-3 Pathway by Cannabinoids in the Brain. J. Neurochem. 2007, 102, 1105–1114. [Google Scholar] [CrossRef]
- Luo, Z.; Chen, S.; Chen, X. CircMAPK9 Promotes the Progression of Fibroblast-like Synoviocytes in Rheumatoid Arthritis via the MiR-140-3p/PPM1A Axis. J. Orthop. Surg. Res. 2021, 16, 395. [Google Scholar] [CrossRef]
- Altman, A.; Kong, K.F. Protein Kinase C Inhibitors for Immune Disorders. Drug Discov. Today 2014, 19, 1217–1221. [Google Scholar] [CrossRef] [Green Version]
- Powles, T.; Te Poele, R.; Shamash, J.; Chaplin, T.; Propper, D.; Joel, S.; Oliver, T.; Wai, M.L. Cannabis-Induced Cytotoxicity in Leukemic Cell Lines: The Role of the Cannabinoid Receptors and the MAPK Pathway. Blood 2005, 105, 1214–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, H.; Liu, X.; Wang, Z.W.; He, D.Y.; Su, D.F.; Dai, S.M. Expression of Cannabinoid Receptor 2 and Its Inhibitory Effects on Synovial Fibroblasts in Rheumatoid Arthritis. Rheumatology 2014, 53, 802–809. [Google Scholar] [CrossRef] [Green Version]
- Weisbart, R.H.; Chan, G.; Heinze, E.; Mory, R.; Nishimura, R.N.; Colburn, K. BRAF Drives Synovial Fibroblast Transformation in Rheumatoid Arthritis. J. Biol. Chem. 2010, 285, 34299–34303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erlandsson, M.C.; Töyrä Silfverswärd, S.; Nadali, M.; Turkkila, M.; Svensson, M.N.D.; Jonsson, I.M.; Andersson, K.M.E.; Bokarewa, M.I. IGF-1R Signalling Contributes to IL-6 Production and T Cell Dependent Inflammation in Rheumatoid Arthritis. Biochim. Biophys. Acta—Mol. Basis Dis. 2017, 1863, 2158–2170. [Google Scholar] [CrossRef] [PubMed]
- Kerola, A.M.; Rollefstad, S.; Semb, A.G. Atherosclerotic Cardiovascular Disease in Rheumatoid Arthritis: Impact of Inflammation and Antirheumatic Treatment. Eur. Cardiol. Rev. 2021, 16, e18. [Google Scholar] [CrossRef]
- Cavagna, L.; Boffini, N.; Cagnotto, G.; Inverardi, F.; Grosso, V.; Caporali, R. Atherosclerosis and Rheumatoid Arthritis: More than a Simple Association. Mediat. Inflamm. 2012, 2012, 147354. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Zeng, Z.; Lang, Y.; Xiang, X.; Qi, G.; Lu, G.; Yang, X. Cannabis Seed Oil Alleviates Experimental Atherosclerosis by Ameliorating Vascular Inflammation in Apolipoprotein-E-Deficient Mice. J. Agric. Food Chem. 2021, 69, 9102–9110. [Google Scholar] [CrossRef]
- Carruthers, D.M.; Naylor, W.G.; Allen, M.E.; Kitas, G.D.; Bacon, P.A.; Young, S.P. Characterization of Altered Calcium Signalling in T Lymphocytes from Patients with Rheumatoid Arthritis (RA). Clin. Exp. Immunol. 1996, 105, 291–296. [Google Scholar] [CrossRef]
- Ye, Z.; Shen, Y.; Jin, K.; Qiu, J.; Hu, B.; Jadhav, R.R.; Sheth, K.; Weyand, C.M.; Goronzy, J.J. Arachidonic Acid-Regulated Calcium Signaling in T Cells from Patients with Rheumatoid Arthritis Promotes Synovial Inflammation. Nat. Commun. 2021, 12, 907. [Google Scholar] [CrossRef]
- Remans, P.; Reedquist, K.; Bos, J.; Verweij, C.; Breedveid, F.; van Laar, J.; Gringhuis, S. Deregulated Ras and Rap1 Signaling in rheumatoid Arthritis T Cells Leads to Persistent Production of Free Radicals. Arthritis Res. Ther. 2002, 4, 18. [Google Scholar]
- Jordan, J.D.; He, J.C.; Eungdamrong, N.J.; Gomes, I.; Ali, W.; Nguyen, T.; Bivona, T.G.; Philips, M.R.; Devi, L.A.; Iyengar, R. Cannabinoid Receptor-Induced Neurite Outgrowth Is Mediated by Rap1 Activation through Gαo/i-Triggered Proteasomal Degradation of Rap1GAPII. J. Biol. Chem. 2005, 280, 11413–11421. [Google Scholar] [CrossRef] [Green Version]
- Miyasaka, N. Nitric Oxide Production in Rheumatoid Arthritis. Jpn. J. Rheumatol. 1997, 7, 165–172. [Google Scholar] [CrossRef]
- Mbvundula, E.C.; Bunning, R.A.D.; Rainsford, K.D. Effects of Cannabinoids on Nitric Oxide Production by Chondrocytes and Proteoglycan Degradation in Cartilage. Biochem. Pharmacol. 2005, 69, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.-K.; Adnan, M.; Cho, D.-H. Drug Investigation to Dampen the Comorbidity of Rheumatoid Arthritis and Osteoporosis via Molecular Docking Test. Curr. Issues Mol. Biol. 2022, 44, 1046–1061. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.; Adnan, M.; Cho, D. Uncovering Mechanisms of Zanthoxylum piperitum Fruits for the Alleviation of Rheumatoid Arthritis Based on Network Pharmacology. Biology 2021, 10, 703. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nandi, A.; Das, A.; Dey, Y.N.; Roy, K.K. The Abundant Phytocannabinoids in Rheumatoid Arthritis: Therapeutic Targets and Molecular Processes Identified Using Integrated Bioinformatics and Network Pharmacology. Life 2023, 13, 700. https://doi.org/10.3390/life13030700
Nandi A, Das A, Dey YN, Roy KK. The Abundant Phytocannabinoids in Rheumatoid Arthritis: Therapeutic Targets and Molecular Processes Identified Using Integrated Bioinformatics and Network Pharmacology. Life. 2023; 13(3):700. https://doi.org/10.3390/life13030700
Chicago/Turabian StyleNandi, Arijit, Anwesha Das, Yadu Nandan Dey, and Kuldeep K. Roy. 2023. "The Abundant Phytocannabinoids in Rheumatoid Arthritis: Therapeutic Targets and Molecular Processes Identified Using Integrated Bioinformatics and Network Pharmacology" Life 13, no. 3: 700. https://doi.org/10.3390/life13030700