
Synthetic Analogues of Gibbilimbol B Induce Bioenergetic Damage and Calcium Imbalance in Trypanosoma cruzi
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and General Procedures
2.2. Hemolytic Activity
2.3. Parasites and Mammalian Cells
2.4. Mechanism of Action (MoA) Studies
2.5. Plasma Membrane Electric Potential (Δψp)
2.6. Mitochondrial Membrane Electric Potential (ΔΨm)
2.7. Adenosine Triphosphate (ATP)
2.8. Reactive Oxygen Species (ROS)
2.9. Intracellular Calcium (Ca2+)
2.10. Acidocalcisomes
2.11. Immunomodulatory Response
2.12. Protein Modelling, Binding Site Identification, Molecular Docking Studies, and Analyses of Crucial Interactions
2.13. Statistical Analysis
3. Results
3.1. Hemolytic Activity
3.2. Mechanism of Action
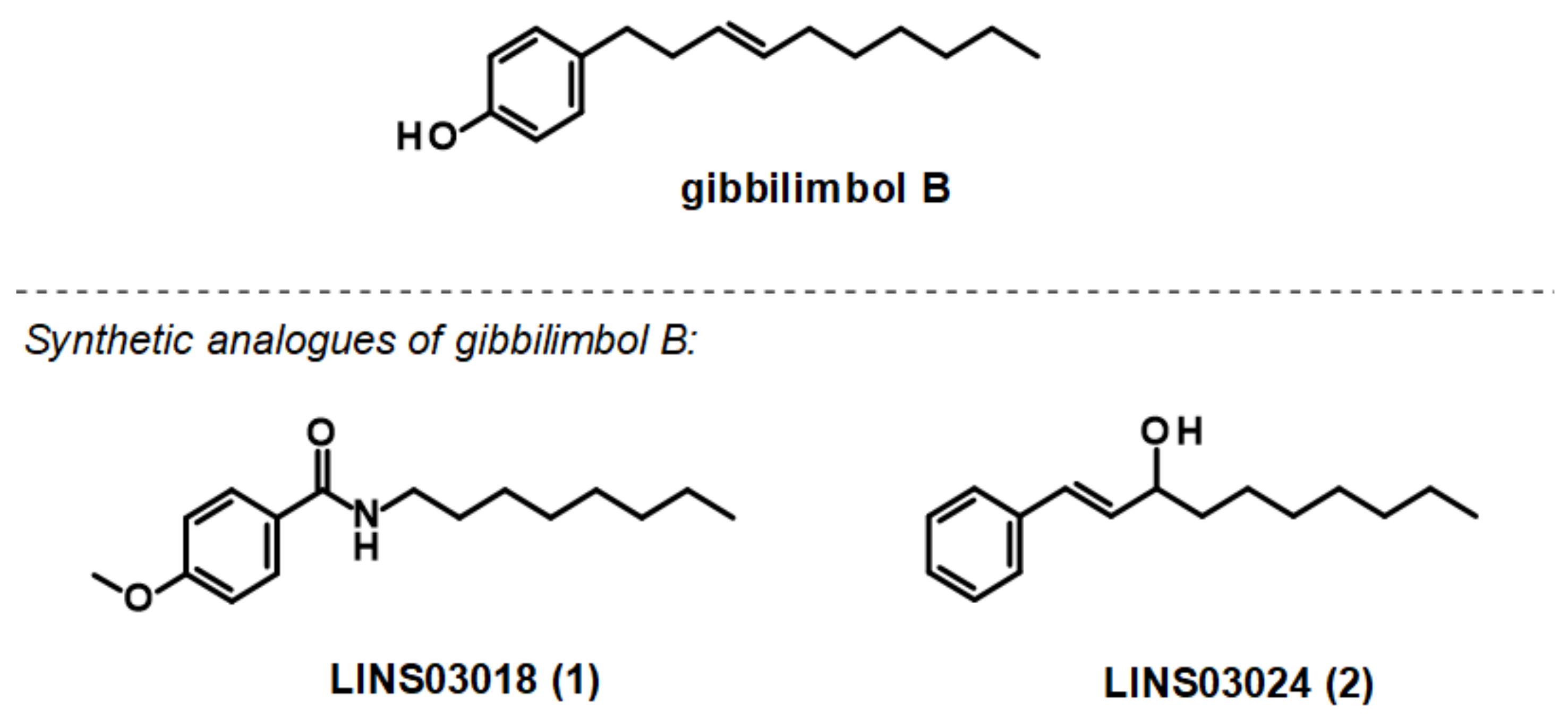
3.3. Plasma Membrane Electric Potential (ΔΨp)
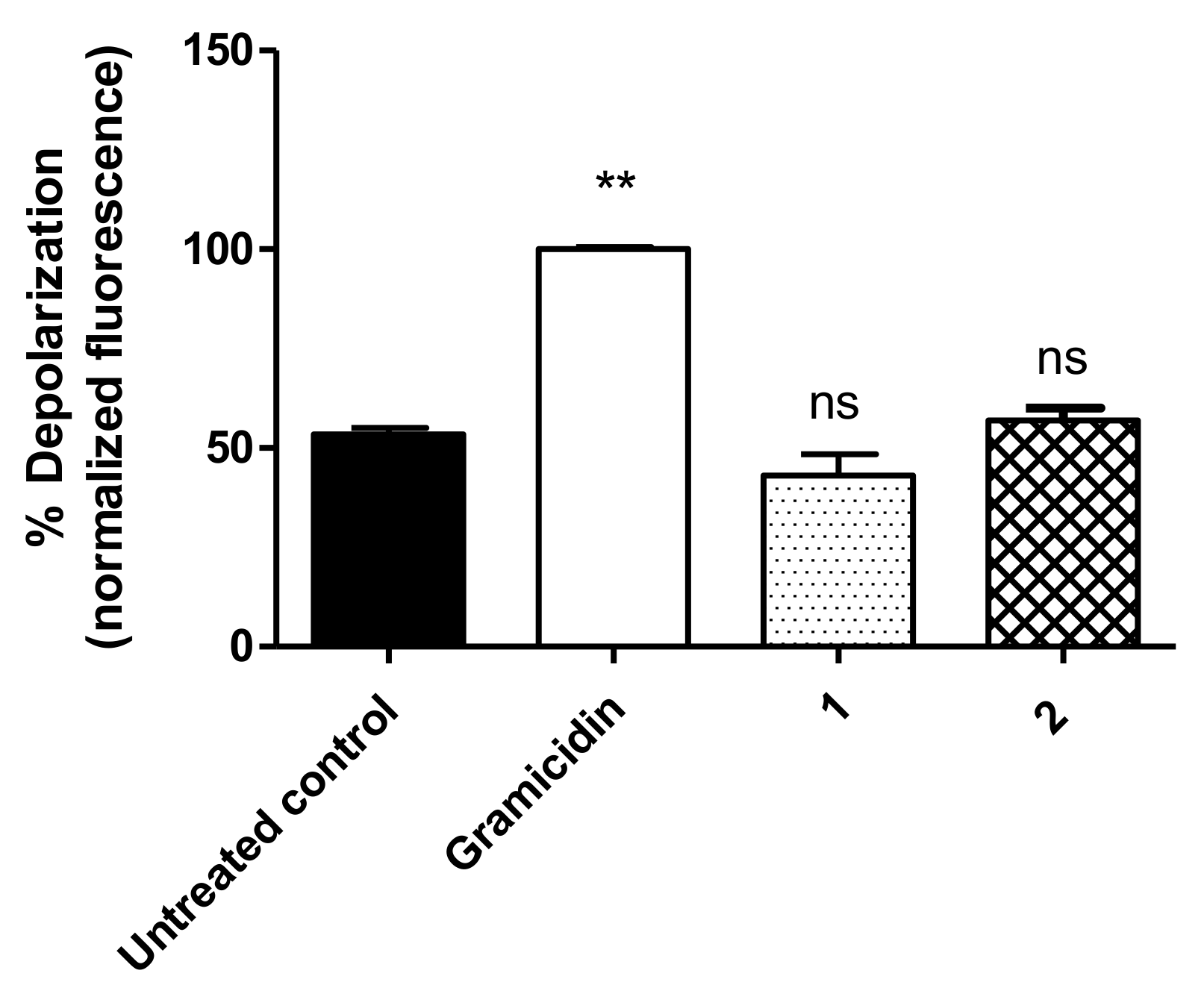
3.4. Mitochondrial Membrane Electric Potential (ΔΨm)
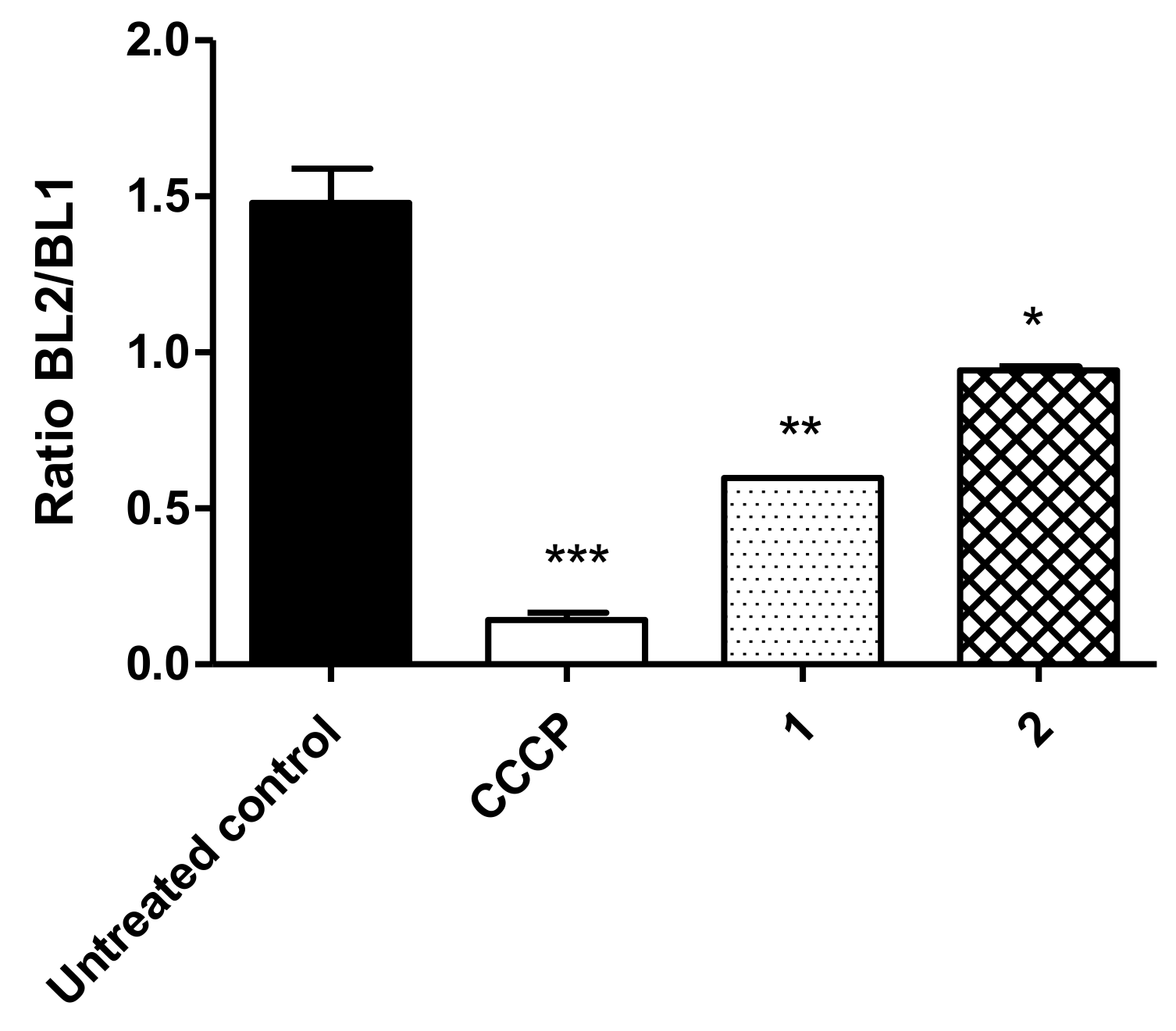
3.5. ATP Levels
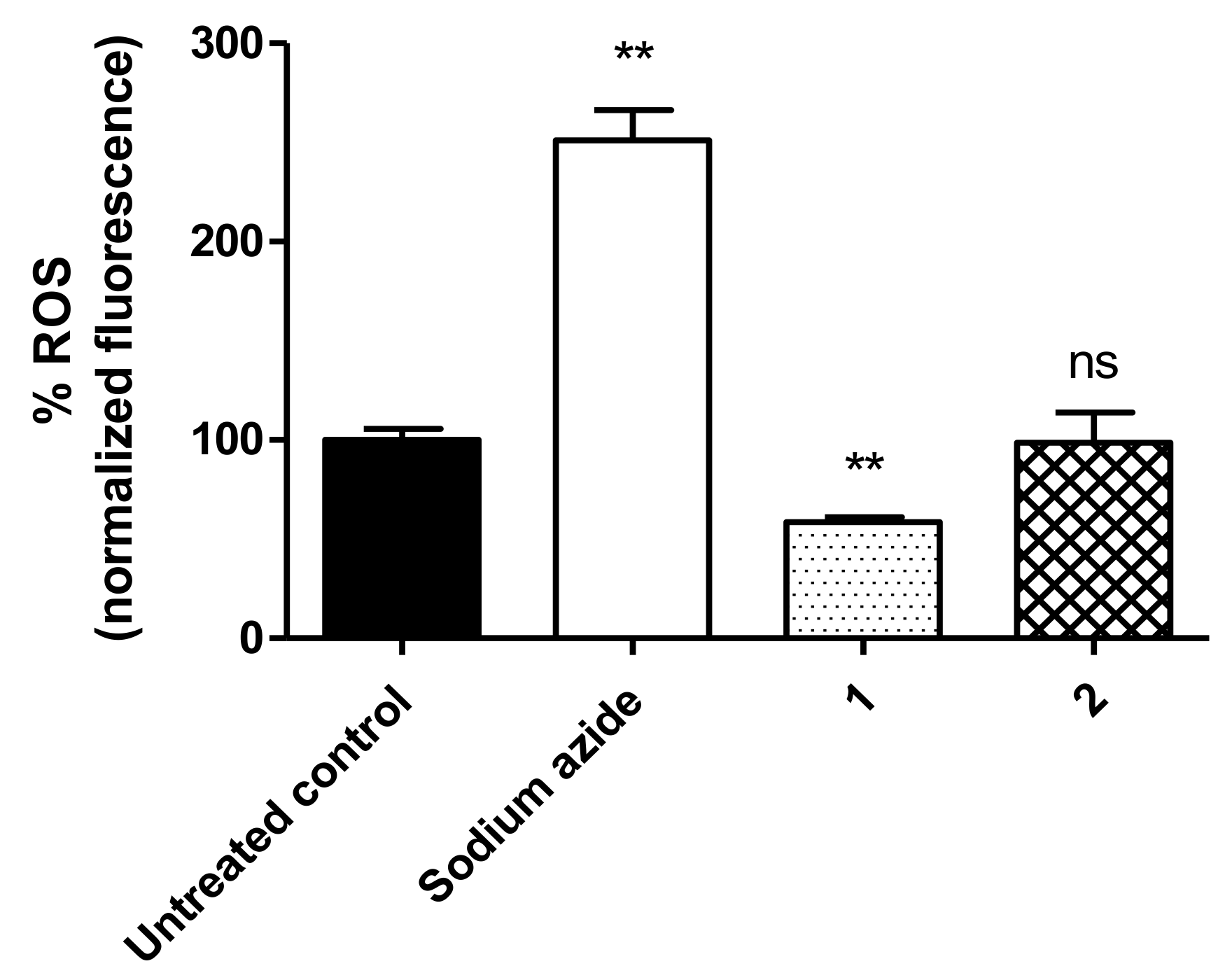
3.6. ROS Levels
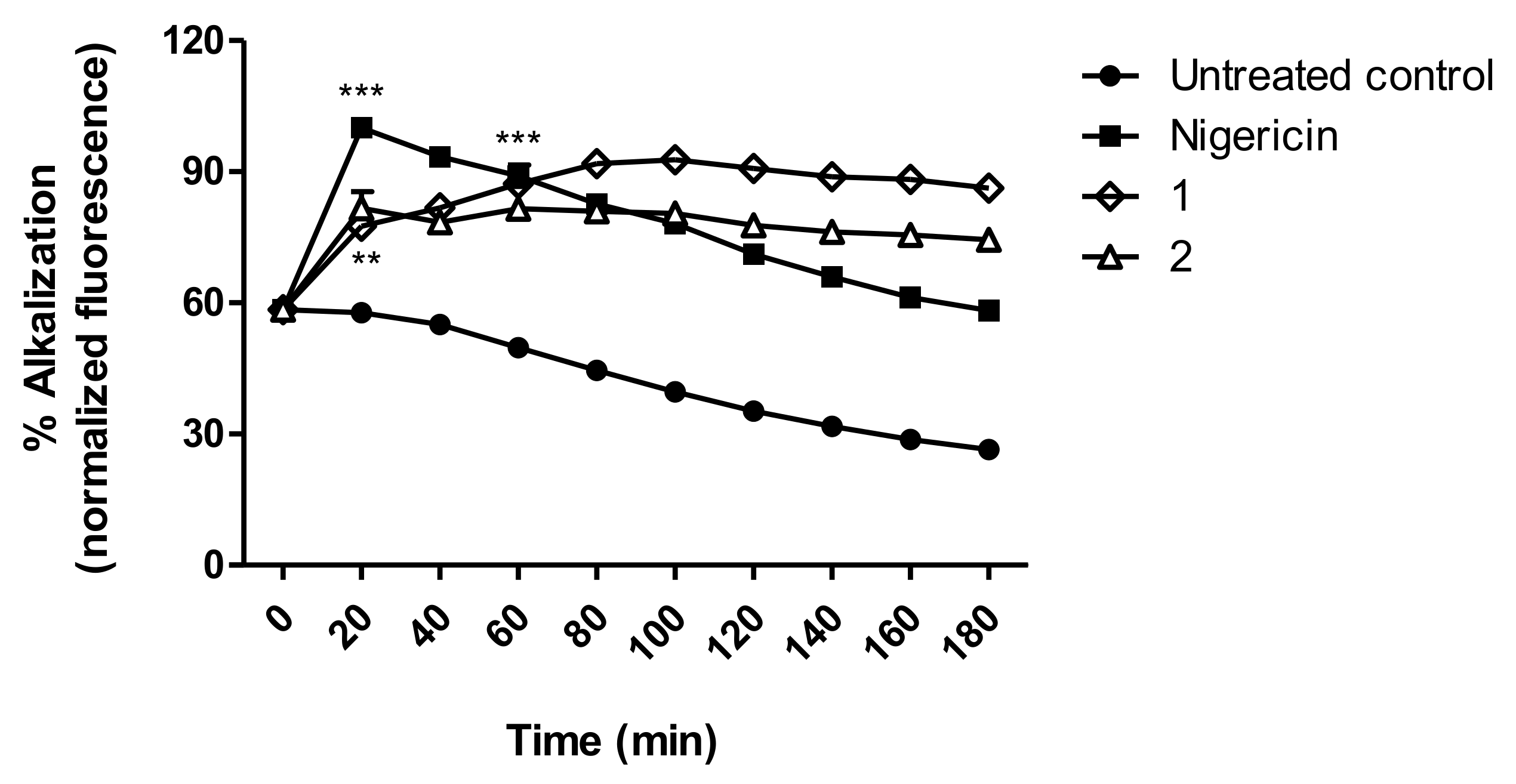
3.7. Acidocalcisomes
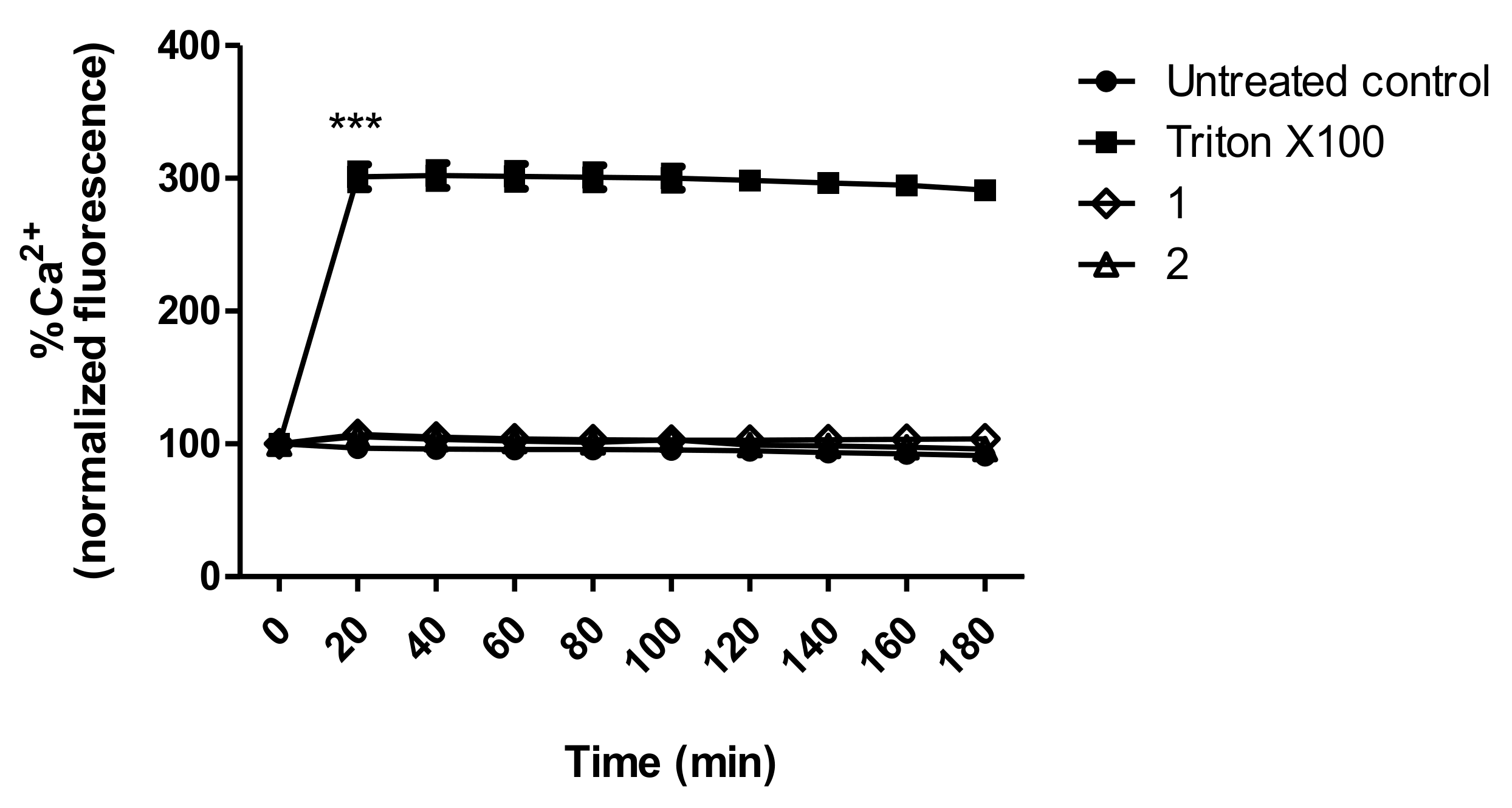
3.8. Intracellular Calcium Levels
3.9. Nitric Oxide Response of Macrophages Infected with T. cruzi
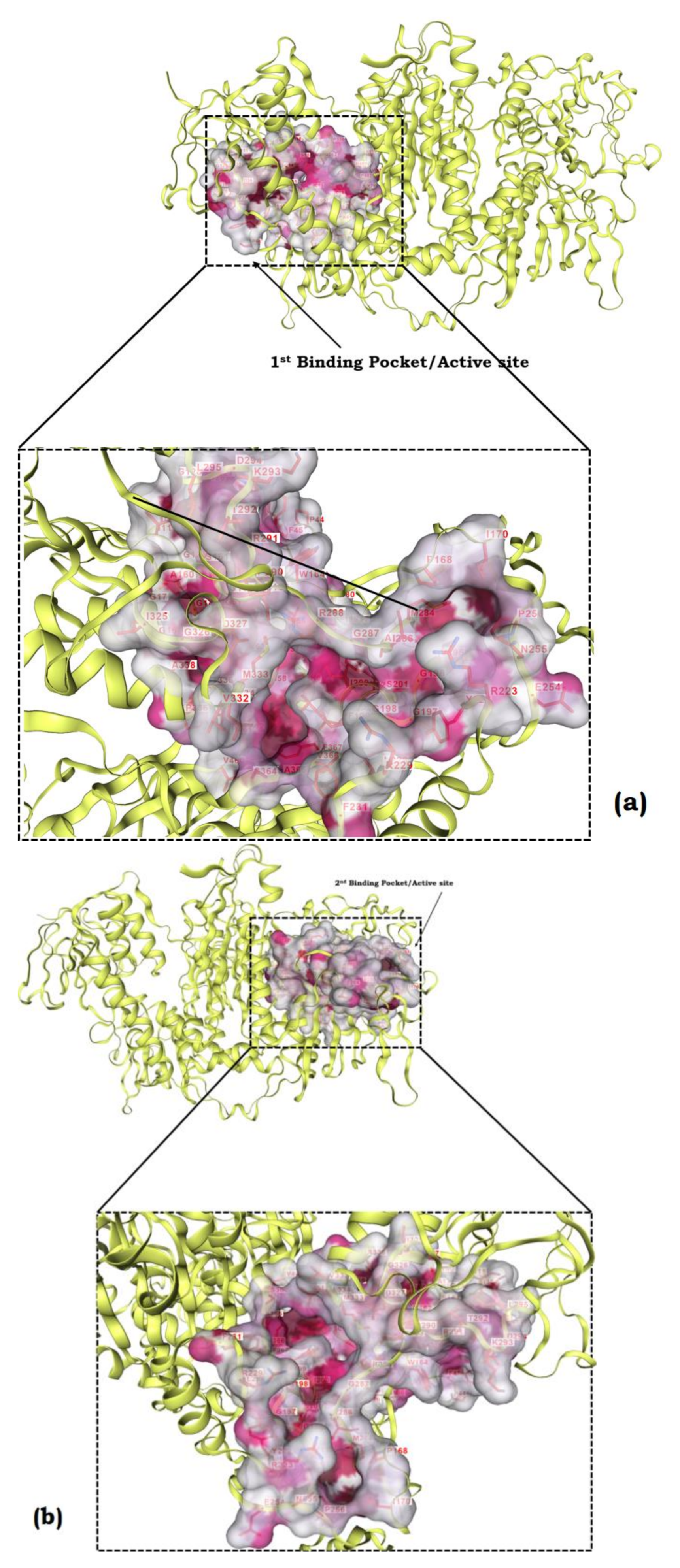
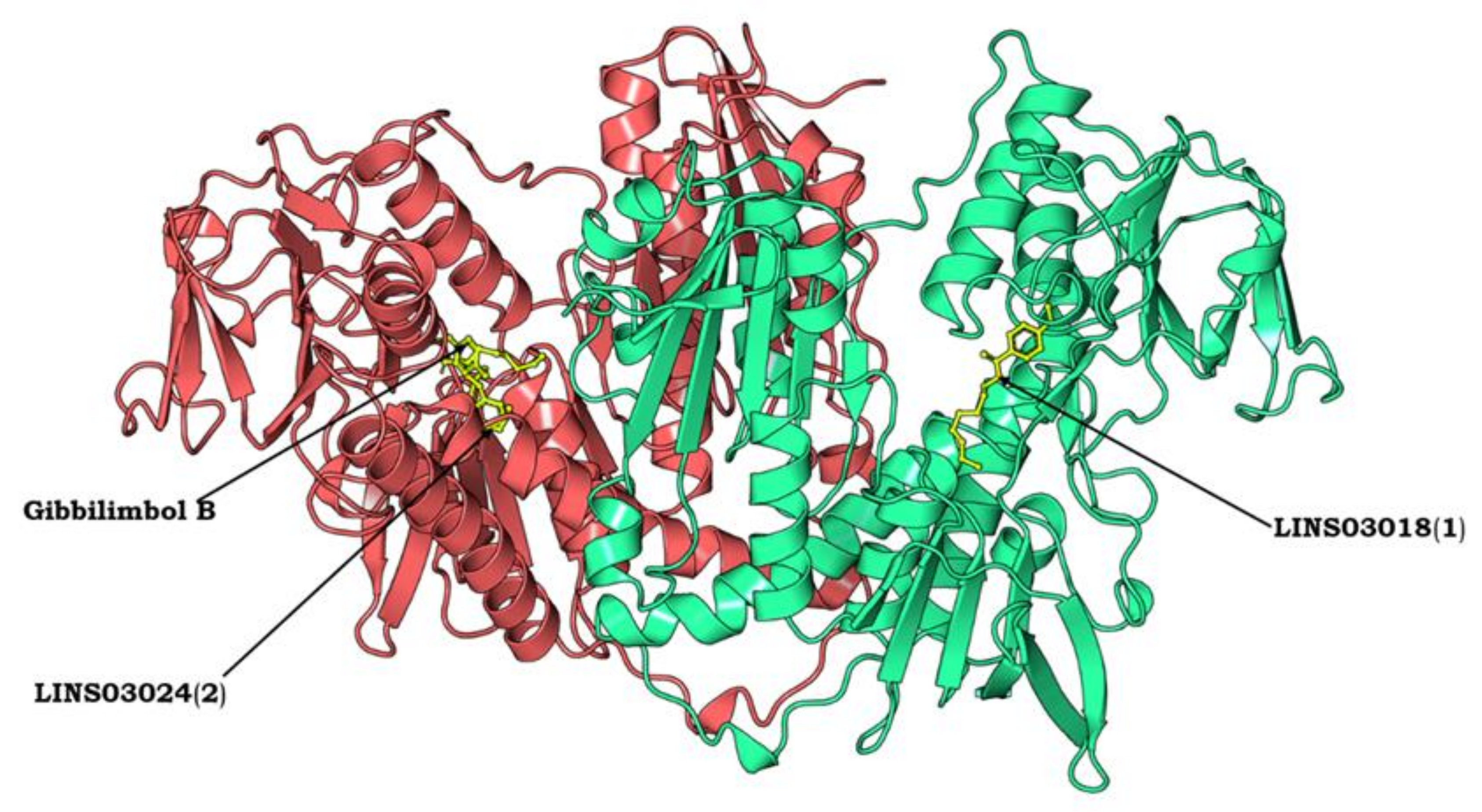
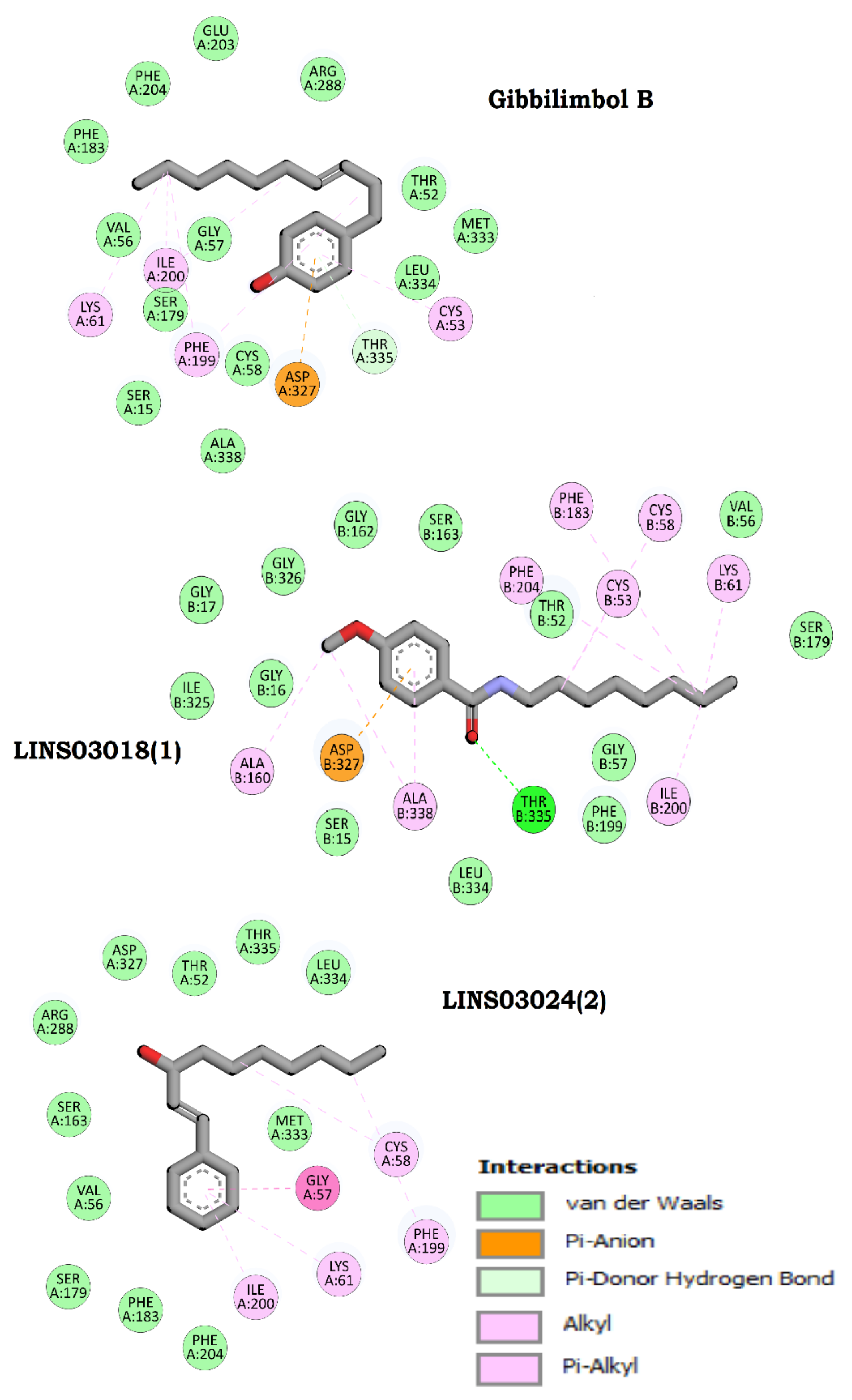
3.10. Protein Modelling, Binding Site Identification, Molecular Docking Studies, and Analyses of Crucial Interactions
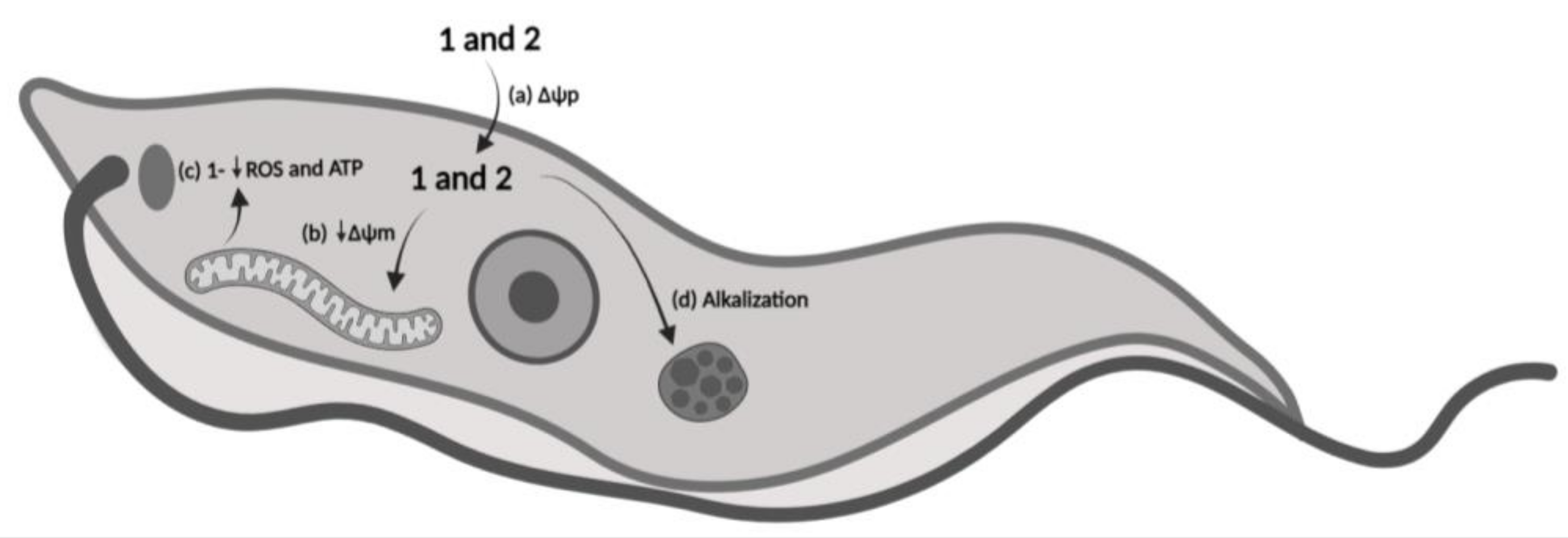
3.11. Mechanism of Action Proposal
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miranda-Schaeubinger, M.; Chakravarti, I.; Lidani, K.C.F.; Omidian, Z.; Gilman, R.H. Systematic Review of the Epidemiology of Chagas Disease in the Americas: A Call for Standardized Reporting of Chagas Disease Prevalence. Curr. Trop. Med. Rep. 2019, 6, 23–34. [Google Scholar] [CrossRef]
- Rodríguez-Bejarano, O.H.; Avendaño, C.; Patarroyo, M.A. Mechanisms Associated with Trypanosoma cruzi Host Target Cell Adhesion, Recognition and Internalization. Life 2021, 11, 534. [Google Scholar] [CrossRef] [PubMed]
- Kratz, J.M. Drug discovery for chagas disease: A viewpoint. Acta Trop. 2019, 198, 105107. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, V.; Dias, N.; Paiva, T.; Hagström-Bex, L.; Nitz, N.; Pratesi, R.; Hecht, M. Current trends in the pharmacological management of Chagas disease. IJP Drugs Drug Resist. 2020, 12, 7–17. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [Green Version]
- Chopra, B.; Dhingra, A.K. Natural products: A lead for drug discovery and development. Phytother. Res. 2021, 35, 4660–4702. [Google Scholar] [CrossRef]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The Traditional Medicine and Modern Medicine from Natural Products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.; Mesquita, J.T.; Tempone, A.G.; Lago, J.H.G.; Guimarães, E.F.; Kato, M.J. Leishmanicidal activity of an alkenylphenol from Piper malacophyllum is related to plasma membrane disruption. Exp. Parasitol. 2012, 132, 383–387. [Google Scholar] [CrossRef] [Green Version]
- Varela, M.T.; Romaneli, M.M.; Lima, M.L.; Borborema, S.E.T.; Tempone, A.G.; Fernandes, J.P.S. Antiparasitic activity of new gibbilimbol analogues and SAR analysis through efficiency and statistical methods. Eur. J. Pharm. Sci. 2018, 122, 31–41. [Google Scholar] [CrossRef]
- Varela, M.T.; Costa-Silva, T.A.; Lago, J.H.G.; Tempone, A.G.; Fernandes, J.P.S. Evaluation of the antitrypanosoma activity and SAR study of novel LINS03 derivatives. Bioorg. Chem. 2019, 89, 102996. [Google Scholar] [CrossRef]
- Varela, M.T.; Dias, R.Z.; Martins, L.F.; Ferreira, D.D.; Tempone, A.G.; Ueno, A.K.; Lago, J.G.H.; Fernandes, J.P.S. Gibbilimbol analogues as antiparasitic agents–synthesis and biological activity against Trypanosoma cruzi and Leishmania (L.) infantum. Bioorg. Med. Chem. Lett. 2016, 26, 1180–1183. [Google Scholar] [CrossRef] [PubMed]
- Rea, A.; Tempone, A.G.; Pinto, E.G.; Mesquita, J.T.; Rodrigues, E.; Silva, L.G.; Sartorelli, P.; Lago, J.H. Soulamarin isolated from Calophyllum brasiliense (Clusiaceae) induces plasma membrane permeabilization of Trypanosoma cruzi and mytochondrial dysfunction. PLoS Negl. Trop. Dis. 2013, 7, e2556. [Google Scholar] [CrossRef] [Green Version]
- Weischenfeldt, J.; Porse, B. Bone marrow-derived macrophages (BMM): Isolation and applications. CSH Protoc. 2008, pdb prot5080. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, H.; Silva, R.L.C.G.; Costa-Silva, T.A.; Tempone, A.G.; Antar, G.M.; Lago, J.H.G.; Caseli, L. Interaction of dicentrinone, an antitrypanosomal aporphine alkaloid isolated from Ocotea puberula (Lauraceae), in cell membrane models at the air-water interface. Bioorg. Chem. 2020, 101, 103978. [Google Scholar] [CrossRef]
- Luque-Ortega, J.R.; Rivas, L. Characterization of the leishmanicidal activity of antimicrobial peptides. Methods Mol. Biol. 2010, 618, 393–420. [Google Scholar] [PubMed]
- Morais, T.R.; Costa-Silva, T.A.; Ferreira, D.D.; Novais, B.J.; Torrecilhas, A.C.T.; Tempone, A.G.; Lago, J.H.G. Antitrypanosomal activity and effect in plasma membrane permeability of (−)-bornyl p-coumarate isolated from Piper cernuum (Piperaceae). Bioorg. Chem. 2019, 89, 103001. [Google Scholar] [CrossRef] [PubMed]
- Umehara, E.; Costa Silva, T.A.; Mendes, V.M.; Guadagnin, R.C.; Sartorelli, P.; Tempone, A.G.; Lago, J.H.G. Differential lethal action of C17:2 and C17:0 anacardic acid derivatives in Trypanosoma cruzi—A mechanistic study. Bioorg. Chem. 2020, 102, 104068. [Google Scholar] [CrossRef] [PubMed]
- Dolai, S.; Yadav, R.K.; Pal, S.; Adak, S. Overexpression of mitochondrial Leishmania major ascorbate peroxidase enhances tolerance to oxidative stress-induced programmed cell death and protein damage. Eukaryot Cell 2009, 8, 1721–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Londero, V.S.; Costa-Silva, T.A.; Tempone, A.G.; Namiyama, G.M.; Thevenard, F.; Antar, G.M.; Baitello, J.B.; Lago, J.H.G. Anti-Trypanosoma cruzi activity of costic acid isolated from Nectandra barbellata (Lauraceae) is associated with alterations in plasma membrane electric and mitochondrial membrane potentials. Bioorg. Chem. 2020, 95, 103510. [Google Scholar] [CrossRef]
- Corral, M.J.; Benito-Peña, E.; Jiménez-Antón, M.D.; Cuevas, L.; Moreno-Bondi, M.C.; Alunda, J.M. Allicin Induces Calcium and Mitochondrial Dysregulation Causing Necrotic Death in Leishmania. PLoS Negl. Trop. Dis. 2016, 10, e0004525. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Martín, X.; Garcia-Marchán, Y.; Fernandez, A.; Rodriguez, N.; Rojas, H.; Visbal, G.; Benaim, G. Amiodarone Destabilizes Intracellular Ca2+ Homeostasis and Biosynthesis of Sterols in Leishmania mexicana. Antimicrob. Agents Chemother. 2009, 53, 1403–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, S.J.; Meltzer, M.S.; Hibbs, J.B., Jr.; Nacy, C.A. Activated macrophages destroy intracellular Leishmania major amastigotes by an L-arginine-dependent killing mechanism. J. Immunol. 1990, 144, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.S.; Choi, E.Y.; Jin, J.Y.; Park, H.R.; Choi, J.I.; Kim, S.J. Kaempferol inhibits P. intermedia lipopolysaccharide-induced production of nitric oxide through translational regulation in murine macrophages: Critical role of heme oxygenase-1-mediated ROS reduction. J. Periodontol. 2013, 84, 545–555. [Google Scholar] [CrossRef]
- Kant, R.; Jha, P.; Saluja, D.; Chopra, M. Identification of novel inhibitors of Neisseria gonorrhoeae MurI using homology modeling, structure-based pharmacophore, molecular docking, and molecular dynamics simulation-based approach. J. Biomol. Struct. Dyn. 2022, 15, 1–14. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Žídek, A.; Potapenko, A.; Bridgland, A. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 30, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Castro Oliveira, L.G.; Brito, L.M.; Moraes Alves, M.M.; Amorim, L.V.; Sobrinho-Júnior, E.P.; Carvalho, C.E.; Franca Rodrigues, K.A.; Arcanjo, D.D.; Graças Lopes Citó, A.M.; Amorim Carvalho, F.A. In Vitro effects of the neolignan 2,3-Dihydrobenzofuran against Leishmania amazonensis. Basic Clin. Pharmacol. Toxicol. 2017, 120, 52–58. [Google Scholar] [CrossRef]
- Katsuno, K.; Burrows, J.N.; Duncan, K.; Hooft van Huijsduijnen, R.; Kaneko, T.; Kita, K.; Mowbray, C.E.; Schmatz, D.; Warner, P.; Slingsby, B.T. Hit and lead criteria in drug discovery for infectious diseases of the developing world. Nat. Rev. Drug Discov. 2015, 14, 751–758. [Google Scholar] [CrossRef]
- Tiwari, N.; Gedda, M.R.; Tiwari, V.K.; Singh, S.P.; Singh, R.K. Limitations of current therapeutic options, possible drug targets and scope of natural products in control of leishmaniasis. Mini Rev. Med. Chem. 2018, 18, 26–41. [Google Scholar] [CrossRef]
- Meade, J.C. P-type transport ATPases in Leishmania and Trypanosoma. Parasite 2019, 26, 69. [Google Scholar] [CrossRef] [Green Version]
- Menna-Barreto, R.F.S.; Castro, S.L. The double-edged sword in pathogenic trypanosomatids: The pivotal role of mitochondria in oxidative stress and bioenergetics. BioMed. Res. Int. 2014, 2014, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brindha, J.; Balamurali, M.M.; Chanda, K. An Overview on the Therapeutics of Neglected Infectious Diseases-Leishmaniasis and Chagas Diseases. Front. Chem. 2021, 9, 622286. [Google Scholar]
- Rajendran, M.; Dane, E.; Conley, J.; Tantama, M. Imaging adenosine triphosphate (ATP). Biol. Bull. 2016, 231, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Machado-Silva, A.; Cerqueira, P.G.; Grazielle-Silva, V.; Gadelha, F.R.; Peloso, E.F.; Teixeira, S.M.R.; Machado, C.R. How Trypanosoma cruzi deals with oxidative stress: Antioxidant defence and DNA repair pathways. Mutat. Res. Rev. 2016, 767, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, E.; Rojas, D.A.; Morales, S.; Miralles, V.; Solari, A. Dual and opposite roles of reactive oxygen species (ROS) in chagas disease: Beneficial on the pathogen and harmful on the host. Oxid. Med. Cell Longev. 2020, 2020, 8867701. [Google Scholar] [CrossRef]
- Ramakrishnan, S.; Docampo, R. Membrane Proteins in Trypanosomatids Involved in Ca2+ Homeostasis and Signaling. Genes 2018, 9, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benaim, G.; Paniz-Mondolfi, A.E.; Sordillo, E.M.; Martinez-Sotillo, N. Disruption of intracellular calcium homeostasis as a therapeutic target against Trypanosoma cruzi. Front. Cell Infect. Microbiol. 2020, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Scarpelli, P.H.; Pecenin, M.F.; Garcia, C.R.S. Intracellular Ca2+ signaling in protozoan parasites: An overview with a focus on mitochondria. Int. J. Mol. Sci. 2021, 22, 469. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, M.S.; Reis-Cunha, J.L.; Bartholomeu, D.C. Evasion of the Immune Response by Trypanosoma cruzi during Acute Infection. Front. Immunol. 2016, 6, 659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piacenza, L.; Trujillo, M.; Radi, R. Reactive species and pathogen antioxidant networks during phagocytosis. J. Exp. Med. 2019, 216, 501–516. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, K.; Paulino, M.; Salas, C.O.; Zarate-Ramos, J.J.; Vera, B.; Rivera, G. Trypanothione Reductase: A Target for the Development of Anti- Trypanosoma cruzi Drugs. Mini Rev. Med. Chem. 2017, 17, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.T.; Lima, M.L.; Galuppo, M.K.; Tempone, A.G.; Oliveira, A.; Lago, J.H.G.; Fernandes, J.P.S. New alkenyl derivative from and analogues: Antiparasitic activity against Trypanosoma cruzi and Leishmania (L.) infantum. Chem. Biol. Drug Des. 2017, 90, 1007–1011. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 Trypomastigote (µM) ± SD | IC50 Amastigote (µM) ± SD | CC50 NCTC Cells (µM) ± SD | SI |
|---|---|---|---|---|
| gibbilimbol B 1 | 75.3 ± 9.2 | >100 | >200 | ND |
| LINS03018 (1) 2 | 11.4 ± 5.8 | 5.2 ± 0.6 | 91.5 ± 34.8 | 17.6 |
| LINS03024 (2) 3 | 9.8 ± 1.5 | 5.8 ± 0.1 | >200 | >34.5 |
| Compound | Structure | Docking Score (kcal/mol) | Estimated ΔG (kcal/mol) | Binding Affinity (kcal/mol) |
|---|---|---|---|---|
| Gibbilimbol B |  | −4290.01 | −7.79 | 5.5 |
| LINS03018 (1) |  | −4288.93 | −7.90 | 6.0 |
| LINS03024 (2) |  | −4287.92 | −7.93 | 5.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amaral, M.; Varela, M.T.; Kant, R.; Christodoulides, M.; Fernandes, J.P.S.; Tempone, A.G. Synthetic Analogues of Gibbilimbol B Induce Bioenergetic Damage and Calcium Imbalance in Trypanosoma cruzi. Life 2023, 13, 663. https://doi.org/10.3390/life13030663
Amaral M, Varela MT, Kant R, Christodoulides M, Fernandes JPS, Tempone AG. Synthetic Analogues of Gibbilimbol B Induce Bioenergetic Damage and Calcium Imbalance in Trypanosoma cruzi. Life. 2023; 13(3):663. https://doi.org/10.3390/life13030663
Chicago/Turabian StyleAmaral, Maiara, Marina T. Varela, Ravi Kant, Myron Christodoulides, João Paulo S. Fernandes, and Andre G. Tempone. 2023. "Synthetic Analogues of Gibbilimbol B Induce Bioenergetic Damage and Calcium Imbalance in Trypanosoma cruzi" Life 13, no. 3: 663. https://doi.org/10.3390/life13030663