

Santin (5,7-Dihydroxy-3,6,4′-Trimetoxy-Flavone) Enhances TRAIL-Mediated Apoptosis in Colon Cancer Cells

Department of Microbiology and Immunology, Faculty of Medical Sciences in Zabrze, Medical University of Silesia, 40-055 Katowice, Poland
*
Author to whom correspondence should be addressed.
Life 2023, 13(2), 592; https://doi.org/10.3390/life13020592
Submission received: 7 November 2022
/
Revised: 5 January 2023
/
Accepted: 13 January 2023
/
Published: 20 February 2023
(This article belongs to the Collection The Role of Bioactive Natural Compounds in the Treatment of Diseases)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:TRAIL (Tumor necrosis factor–Related Apoptosis-Inducing Ligand) has the ability to selectively kill cancer cells without being toxic to normal cells. This endogenous ligand plays an important role in surveillance and anti-tumor immunity. However, numerous tumor cells are resistant to TRAIL-induced apoptosis. In this study, the apoptotic effect of santin in combination with TRAIL on colon cancer cells was examined. Flow cytometry was used to detect the apoptosis and expression of death receptors (TRAIL-R1/DR4 and TRAIL-R2/DR5). Mitochondrial membrane potential (ΔΨm) was evaluated by DePsipher staining with the use of fluorescence microscopy. We have shown for the first time that flavonoid santin synergizes with TRAIL to induce apoptosis in colon cancer cells. Santin induced TRAIL-mediated apoptosis through increased expression of death receptors TRAIL-R1 and TRAIL-R2 and augmented disruption of the mitochondrial membrane in SW480 and SW620 cancer cells. The obtained data may indicate the potential role of santin in colon cancer chemoprevention through the enhancement of TRAIL-mediated apoptosis.
1. Introduction
Flavonoids belong to polyphenols synthesized in plants as bioactive secondary metabolites that affect their flavor, color, and pharmacological activities. Moreover, flavonoids play a significant role in the physiology of plants. By imparting color to flowers, they help attract insects. In addition, flavonoids protect plant cells from the harmful effects of ultraviolet radiation as well as perform function of plant hormones and growth regulators [1]. In humans, these compounds are associated with many health benefits which result from their broad bioactive properties, such as anti-inflammatory, anticancer, antioxidant, cytoprotective, immunomodulatory, and antimicrobial properties [2,3,4].
The structure of flavonoids is composed of two benzene rings (A and B) linked by a heterocyclic pyran ring (C-ring). According to their molecular structure, they are subdivided into different groups: anthocyanidins, flavanols, flavanones, flavonols, flavones, and isoflavones.
Flavonoids play a major role in the human diet. The main flavonoid sources are fruits, vegetables, grains, herbs, and beverages (e.g., wine, tea, and beer) [5,6,7,8]. There is still a growing interest in the field of the research on flavonoid compounds. It was shown repeatedly, that high intake of flavonoids may provide protection against oxidation, inflammation, and consequently against cancer and chronic diseases. These compounds are able to change the activity of cell enzymes and affect the cellular systems, which can be very beneficial to the organism [2,3].
The main biological activity of these substances, which has been studied for years, is their antioxidant activity. These properties of flavonoids are structure dependent, due to the position of the B-ring on the pyran C-ring but mostly to the presence and location of hydroxyl groups in their molecule. The functional hydroxyl groups demonstrate their antioxidative properties by scavenging free radicals, chelating metal ions, but also by activation of antioxidant enzymes and inhibition of oxidases [9].
The anti-inflammatory properties have an important impact on the development of cancer. Several mechanisms have been proposed to explain anticancer properties of flavonoids. These compounds can affect the initiation and promotion stages of cancer development by, for example, induction of apoptosis, inhibiting of cell cycle, anti-proliferative effect and growth inhibition but also by affecting angiogenesis [5,10,11]. Moreover, it was demonstrated that elevated levels of ROS can be oncogenic, acting destructive on nucleic acids, proteins and lipids. Consequently it can promote genetic instability and lead to cancerogenesis. Therefore, flavonoids have been shown to inhibit tumor cell generation and proliferation by inhibition of ROS formation and reducing oxidative stress [11].
Apoptosis is a form of programmed cell death and plays a crucial role in cancer prevention. During apoptosis significant changes occur, such as nuclear chromatin condensation, DNA fragmentation and apoptotic body formation. Chemoprevention plays a significant role in inhibiting the process of carcinogenesis. Many in vitro studies have shown that flavonoids exhibit anti-cancer effects through the induction of apoptosis. This effect results from impact of flavonoids on pro- and antiapoptotic proteins, as well as on proteins that regulate cell cycle pathways. Flavonoids inhibit cell proliferation of tumor cells in the G1/S and G2/M phases of the cell cycle [12]. Either natural or synthetic flavonoids can induce apoptosis in many cancer cells. Apoptosis of cancer cells under the influence of flavones depends on the type of cells and the concentration of flavonoids used. At low concentrations, leukemias are more susceptible than solid tumors such as skin, prostate or lung cancer [13].
Compared to other flavonoids, flavones have a double bond between the second and third carbon in the flavonoid backbone. They have no substituents at the third carbon and are also oxidized at the fourth carbon [14]. Flavones are usually found in plants as O-and C-glycosides. It is estimated that an adult human takes in about 1 g of flavonoid compounds per day, mainly in the form of glycosides. In the gastrointestinal tract the deglycosylation process occurs and aglycones are absorbed in the small intestine via β-glucosidase present in the intestinal epithelium. The dietary flavonoids appear in the plasma not only as aglycones but also as sulfate or glucuronidates. Also, a significant portion of flavonoids consumed undergoes conversion by enzymes of the digestive tract microflora to simple phenols [15].
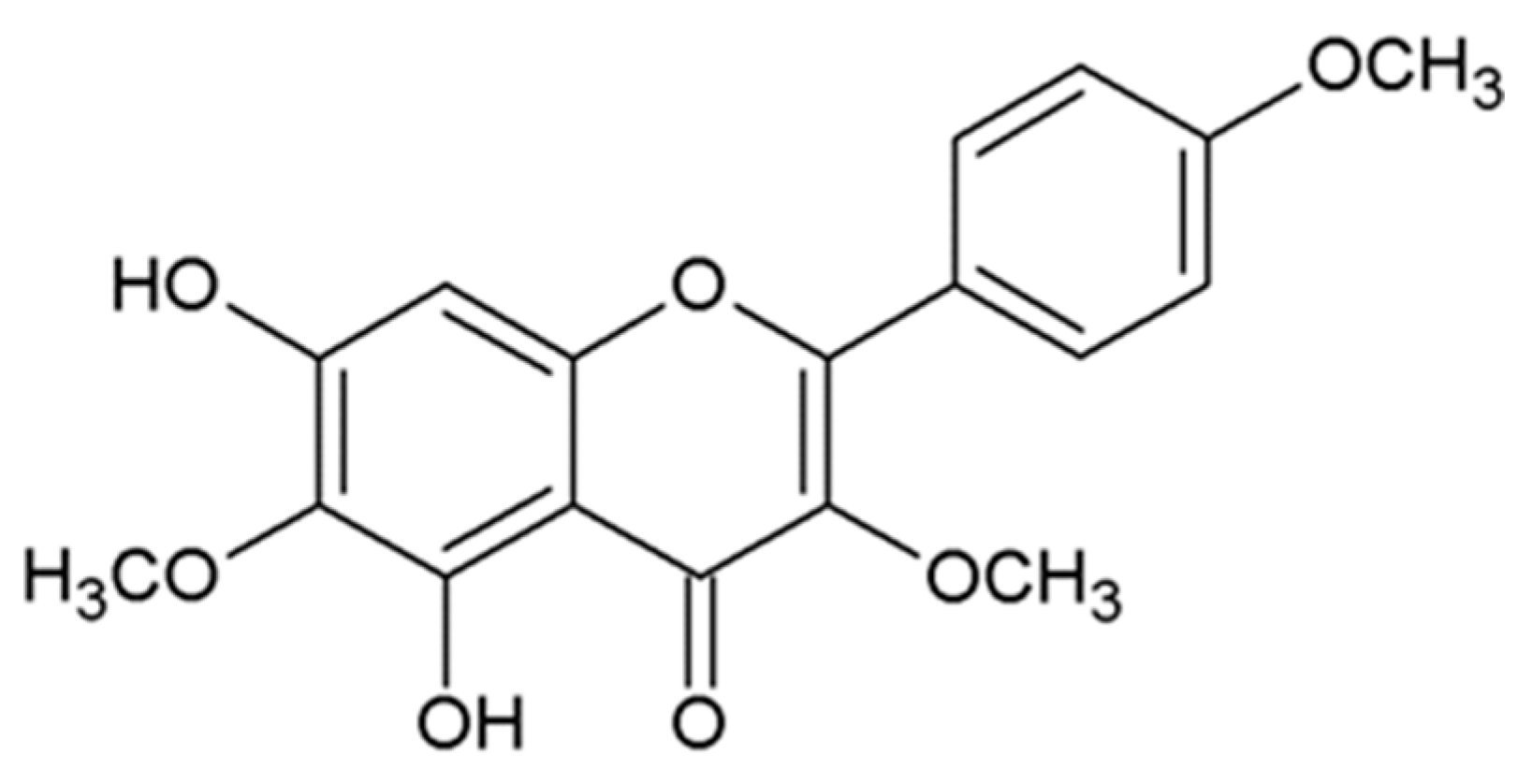
Santin (5,7-dihydroxy-3,6,4’-trimetoxy-flavone) is a relatively unknown compound that belongs to the flavones, which possess antimicrobial, antioxidant, anti-inflammatory, and anticancer properties. Numerous epidemiological and experimental studies provide evidence for the potential beneficial effects of flavones in neoplastic diseases. Santin is isolated from aerial parts of Tanacetum parthenium [16], Tanacetum microphyllum [17], the leaves of Dodonaea angustifolia [18], the buds or leaves of Betula pubescens and Betula pendula [19], and all plant parts of Achillea cappadocica, Achillea setacea, Achillea santolinoides, and Achillea arabica [20]. The compound structure is presented in Figure 1. In the available medical literature databases, little is known about the biological activity of santin.
TRAIL/Apo2L (tumor necrosis factor-related apoptosis-inducing ligand) is a cytokine that belongs to the tumor necrosis factor (TNF) superfamily. It was discovered by two independent research groups in the middle of the 1990s [21,22]. TRAIL exists as a transmembrane protein on the surface of the immune system cells: T cells, natural killer (NK) cells, natural killer T cells (NKT cells), macrophages, and dendritic cells. It can also be cleaved into a soluble form (sTRAIL) [23]. TRAIL has the ability to induce apoptosis in different types of cancer cells with no toxicity to normal cells. The pre-clinical and clinical trials show that the recombinant human TRAIL (rhTRAIL) is a promising anticancer therapeutic agent. It is well tolerated, but the therapeutic effects were often insufficient in patients with cancer because some cancer cells are resistant to apoptosis mediated by TRAIL.
Five TRAIL receptors have been identified so far. Cytoplasmic death domains (DDs) in TRAIL-R1/DR4 and TRAIL-R2/DR5, called death receptors, activate an extrinsic pathway of apoptosis in a caspase-dependent manner in cancer cells. The remaining three receptors (TRAIL-R3/DcR1, TRAIL-R4/DcR2, osteoprotegerin/OPG) are called ‘decoy receptors’ [24,25]. TRAIL induces apoptosis through binding to its death receptors TRAIL-R1 and/or TRAIL-R2. It leads to the recruitment of caspase-8 and FAS-associated protein with death domain (FADD) to the formation of functional death-inducing signaling complex (DISC). Thereafter, caspase 8 is cleaved, and it causes cleave caspase 3 and BH3-interacting domain death agonist (BID). The BID in a truncated form (tBID) translocates to the mitochondria and binds to proapoptotic proteins Bax (Bcl-2-associated X protein) and Bak (Bcl-2 homologous antagonist killer) to undergo oligomerization. It results in the formation of pores in the mitochondrial membrane, mitochondrial outer membrane permeabilization, and release of cytochrome c and Smac/DIABLO (second mitochondria-derived activator of caspases/direct IAP binding protein with low pI) into the cytosol. The apoptotic protease-activating factor 1 (APAF-1), cytochrome c, and caspase 9, along with ATP, form apoptosome, which is necessary to activate caspase 9. The caspase-9 in its active form can activate effector caspases: -3, -6, and -7 and then induce apoptosis. In type, I cancer cells, the ’extrinsic pathway,’ in which death receptors and the activation of caspase-8 are involved, is sufficient to activate caspase-3 and lead to apoptosis. In other cells named type II, the induced apoptosis requires amplification of the signal via the ‘intrinsic pathway’ involvement of the mitochondria [26]. Several mechanisms of TRAIL resistance in tumor cells have been described [27]. The cause is a decreased expression of death receptors and pro-apoptotic proteins and also an increased expression of decoy receptors and anti-apoptotic proteins in a cancer cell which leads to the insensitivity of the cytotoxic and apoptotic effects of TRAIL.
Inducing apoptosis is one of the main strategies of cancer chemoprevention. We and others have shown that natural and/or synthetic flavonoids can sensitize tumor cells to TRAIL-mediated apoptosis [28,29,30,31].
In this study, the apoptotic and cytotoxic effects of rhTRAIL combined with santin on SW480 and SW620 colon cancer cells were examined. We investigated for the first time the mechanisms by which santin enhances TRAIL/Apo2L-mediated apoptosis in cancer cells.
2. Materials and Methods
2.1. Reagents
Santin (5,7-dihydroxy-3,6,4′-trimetoxy-flavone) was obtained from Alexis Biochemicals (San Diego, CA, USA). It was dissolved in 100 mM of DMSO to obtain a final concentration equal to 0.1% (v/v) in the culture media. The tested santin’s purity was ≥98% (HPLC). Soluble recombinant human TRAIL (rhsTRAIL) was bought from PeproTech Inc. (Rocky Hill, NJ, USA).
2.2. Cell Culture
The experiments were conducted on SW480 and SW620 human colon cancer cells purchased from ATCC (American Type Culture Collection, Manassas, VA, USA). These colon cancer cell lines come from the same patient. The SW480 cell line was obtained from a Dukes’ type B primary adenocarcinoma of the colon (grown through the muscular layer of the large intestine but non-metastatic). The SW620 (Dukes’ type C) cell line was isolated a year later than the line SW480 and was derived from a lymph node after cancer recurred with widespread metastasis. The cells were grown in monolayer cultures in Leibovitz’s L-15 medium supplemented with 10% heat-inactivated fetal bovine serum (FBS), 100 μg/mL streptomycin, and 100 U/mL penicillin at 37 °C in a humidified atmosphere of 100% air. The reagents for the cell culture were obtained from ATCC or PAA Laboratories (Pasching, Austria). The passages were conducted twice a week. The adhered SW480 and SW620 cells were treated with a 0.25% trypsin solution. Suspensions of 0.5 × 106 cells in 1 ml of medium were used for experiments [32].
2.3. Apoptosis Determination by Flow Cytometry with Annexin V-Fitc Staining
Apoptosis was detected by flow cytometry with the FITC Annexin V Apoptosis Detection Kit (Becton Dickinson Biosciences, San Jose, CA, USA). The SW480 and SW620 cells (0.5 × 106/mL) were adhered in 24-well plates for 48 h before the experiment and then exposed to TRAIL (25–100 ng/mL) and/or santin (25 μM–100 μM) for 48 h. After this incubation, the cells treated with a 0.25% trypsin solution were washed two times with PBS (phosphate-buffered saline solution) and resuspended in 1× Binding Buffer (100 µL). Afterward, 290 μL of the cell suspension was subject to incubation with (5 μL of propidium iodide and 5 μL of annexin V-FITC for 10 min in the dark at room temperature. The population of annexin V-positive cells was analyzed by flow cytometry (LSR II, Becton Dickinson Biosciences, San Jose, CA, USA) within 1 h [33].
2.4. Analysis of Death Receptor Expression on the Cancer Cell Surface by Flow Cytometry
The expression of TRAIL-R1 and TRAIL-R2 receptors on the cell surface was determined by flow cytometry (LSR II, Becton Dickinson Biosciences, San Jose, CA, USA). SW480 and SW620 cancer cells (0.5 × 106/mL) were adhered in 24-well plates for 48 h and stimulated with santin (50–100 μM). Accutase Solution (PAN-Biotech GmbH, Aidenbach, Germany) was used to harvest the cells, and then the cells were washed twice in PBS and resuspended in PBS containing 0.5% BSA (bovine serum albumin) [18].
2.5. Mitochondrial Membrane Potential
The mitochondrial membrane potentials were measured by The DePsipher Kit (R and D Systems, Minneapolis, MN) in fluorescence microscopy. SW480 and SW620 cancer cells (5 × 105/mL) were seeded in a plate of 24-well for 24 h prior to the experiments. TRAIL (100 ng/mL) with or without santin (50–100 μM) was added to the cancer cells. After 48 h, the cells were washed with PBS and trypsinized to separate them from the cell culture. The cancer cells were incubated with DePsipher solution (5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl-benzimidazolyl carbocyanine iodide) at a concentration of 5 μg/mL for 30 min at 37 °C in the dark. After that, the cells were washed with a reaction buffer containing a stabilizer. Then cell suspension was placed on a glass slide and covered with a glass cover slip. A fluorescence inverted microscope IX51 (Olympus, Tokyo, Japan) with filter sets for FITC and TRITC was used to observe stained cells. DePsipher visualizes the potential-dependent accumulation in mitochondria, indicated by a fluorescence emission shift from red (590 nm) to green (530 nm). The cells were counted, and the number of cells with disruption of the ΔΨm was expressed as a percentage of the total cells.
2.6. Statistical Analysis
The data are shown as the mean ± standard deviation of three experiments performed independently in duplicate or quadruplicate (n = 2 or n = 4). The results were subject to analysis by ANOVA or Student’s t-test. The value of p < 0.05 was regarded as significant. StatSoftStatistic version 12 and Microsoft Excel 2010 were used to perform statistical analysis.
3. Results
We investigated the apoptotic effects of TRAIL at the concentration of 25–100 ng/mL in combination with santin at the concentration of 25–100 μM on SW480 and SW620 cancer cells. Santin significantly increased TRAIL-induced apoptosis to 42.68% ± 0.74%–73.78% ± 0.62% in SW480 cancer cells and to 39.90% ± 0.70%–93.67% ± 0.62% in SW620 cancer cells in comparison to TRAIL alone (Figure 2). Santin cooperates with TRAIL to induce apoptosis in cancer cells.
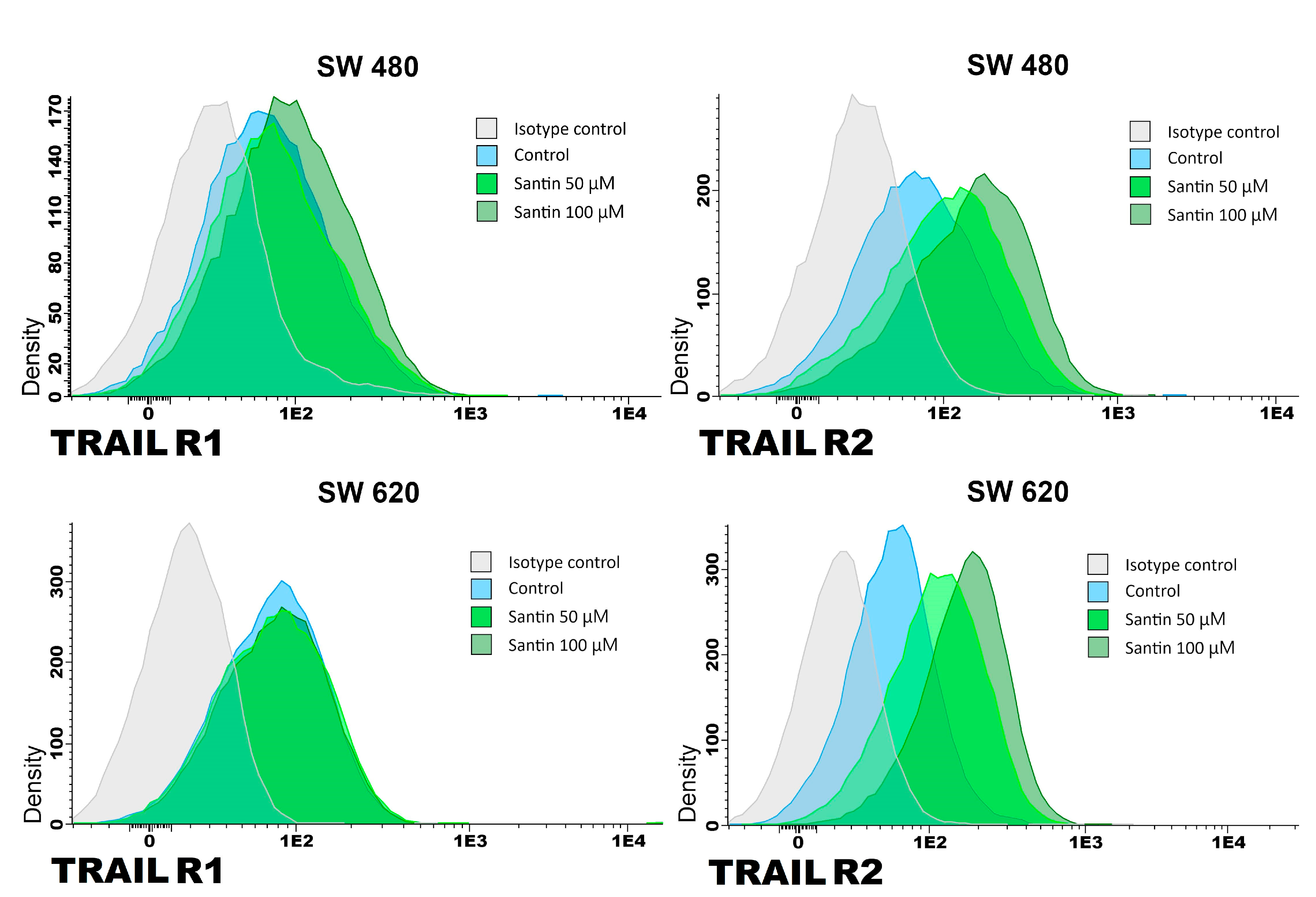
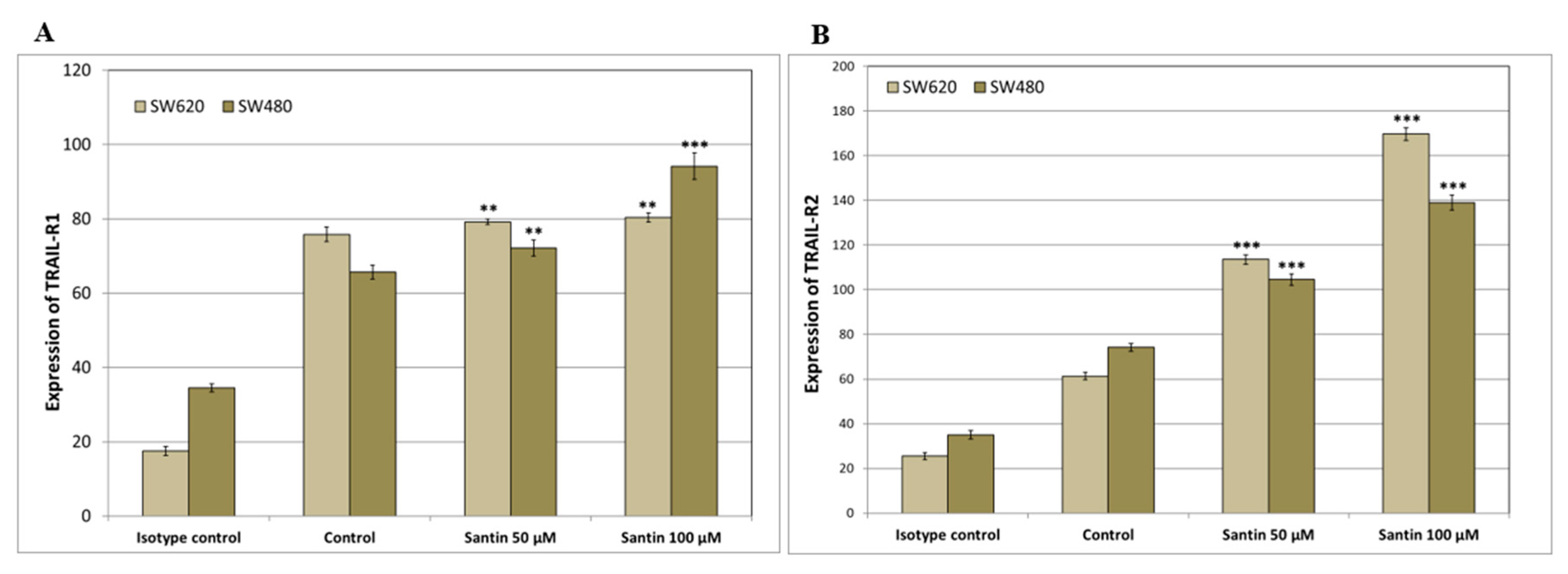
We analyzed the expression of TRAIL-R1 and TRAIL-R2 proteins in SW480 and SW620 cancer cells after 24-hour treatment with santin at the concentration of 50–100 μM by flow cytometry (Figure 3 and Figure 4).
Santin significantly increased TRAIL-R1 and TRAIL-R2 protein levels on the surface of both examined cell lines in all tested concentrations.
In order to check whether the induction of apoptosis by the combination of TRAIL and santin was mediated through death receptor TRAIL-R1 and/or TRAIL-R2, we used the TRAIL-R1/Fc and TRAIL-R2/Fc chimera proteins, which have a dominant negative function against TRAIL-R1/TRAIL-R2. The proteins blocked apoptosis caused by the co-treatment of santin with TRAIL. The obtained results suggest that santin exerted the apoptotic effect of TRAIL through the intrinsic pathway.
Changes in the permeability of the mitochondrial membrane and the loss of the mitochondrial membrane potential (ΔΨm) led to apoptosis. The intrinsic mitochondrial pathway plays a crucial role in amplifying TRAIL-induced apoptosis, and the collapse of (ΔΨm) is considered to be a hallmark of this pathway. We determined whether santin sensitizes mitochondrial dysfunction induced by TRAIL. Incubation of SW480 and SW620 cancer cells with 100 ng/ml TRAIL or 50–100 μM santin alone caused a little effect on ΔΨm (12.67% ± 1.50%, 11.89% ± 1.54% and 14.22% ± 1.56% respectively for SW480 cells and 13.11% ± 1.90%, 12.22% ± 1.39% and 14.00% ± 1.66% respectively for SW620 cells). Either TRAIL or santin treatment had little effect on ΔΨm, but when the cancer cells were subjected to the combined treatment, TRAIL with santin ΔΨm was significantly increased. The incubation of colon cancer cells with TRAIL in combination with santin resulted in a synergistic enhancement of ΔΨm loss in a large percentage of cancer cells to 55.67% ± 2.96% for SW480 cells and to 62.00% ± 4.80% for SW620 cells (Figure 5).
The results show that santin, in combination with TRAIL, is involved in the mitochondrial (intrinsic) pathway in cancer cells.
4. Discussion
Numerous epidemiological and experimental studies confirm the possible beneficial effects of flavonoids on human health [34]. In vitro and in vivo tests have shown that flavonoids could exert immunomodulatory and strong anticancer activities [35,36,37,38]. There are some clinical trials based on flavonoid administration in neoplastic diseases. For example, apigenin is in a phase II clinical trial in colorectal cancer treatment, and genistein is in phase I/II clinical trial in the colon and rectal cancer treatment [39]. Cancer is a heterogeneous disease that is characterized by uncontrolled growth and spread of abnormal cells which invade and metastasize to other parts of the body [40]. Flavonoids possess a wide variety of anticancer effects: they control ROS-scavenging enzyme activities, partake in arresting the cell cycle, suppress the proliferation or invasiveness of cancer cells, and induce apoptosis [41,42,43,44,45,46]. Cancer cells are resistant to apoptosis, an ordered and orchestrated death of cells that is caused by a number of signal transduction pathways as well as pro-apoptotic proteins (Bcl-2 family proteins and caspases) [35]. Apoptosis is initiated by two major pathways, i.e., extrinsic that is associated with the superfamily of tumor necrosis factor (TNF) with main signaling protein death receptors TRAIL-R1/DR4 and TRAIL-R2/DR5 and caspase 8, and intrinsic pathway, involving the mitochondria, in which Bcl-2 family proteins activate caspases 9, 3 and 7 [47,48].
Chemoprevention using naturally occurring flavonoids, due to its incidence, prevalence, and disease-related morbidity and mortality, is an attractive option in colon cancer [36,49]. Epidemiological studies have demonstrated that flavonoid intake was inversely associated with colon cancer risk [50,51]. It has already been demonstrated that the chemopreventive activity of flavonoids is exerted through the modulation of apoptotic signaling pathways [52].
Endogenous TRAIL expressed on monocytes, dendritic cells, macrophages, natural killer cells, and activated T cells is an important component of the immune defense [53]. Dysregulation of apoptotic pathways plays a significant role in the initiation and development of colon cancer [54]. Numerous in vitro tests suggest that dietary flavonoids, due to their chemopreventive advantages, are associated with the enhancement of TRAIL-induced death in cancer cells [28,29,55,56,57,58,59]. Despite the fact that the majority of colon cancer cell lines are resistant to TRAIL-mediated apoptosis, TRAIL, in combination with flavonoids, leads to the synergistic induction of cell death [60,61] Further studies on the intracellular mechanism of TRAIL-mediated apoptosis may help overcome TRAIL resistance and develop approaches to colon cancer prevention based on flavonoids.
Many in vitro studies have shown that natural and synthetic flavonoids enhance apoptosis in cancer cells. Wang et al. indicated that baicalein markedly induced apoptosis in HCT-116 colorectal cancer cells. Caspase activation are critical point in programmed cell death. By exploring the apoptotic mechanism of baicalein, the activation of caspase 3 and 9 were assayed [62]. Ha et al. found the augmentation of apoptosis in HCT-15 and HT-29 colon cancer cells through increased activation of caspase 3 and 9 and alteration of mitochondrial membrane potential and dysfunction [63]. An essential step in tumorigenesis is tumor migration and invasion. Xu M. demonstrated that apigenin at 20 and 40 µM significantly inhibited migration and invasion in SW480 colon cancer cells [64]. The anti-tumor growth and anti-metastasis effects of apigenin were obtained by Chunhua Li in three colorectal adenocarcinoma cell lines: SW480, DLD-1, and LS174T [65]. The protective effects of dietary apigenin enrichment in a chronic colitis model induced by dextran sulfate sodium in mice has been shown by Márquez-Flores Y. [66]. The studies of Xiao-Yu Ai confirm that apigenin effectively inhibits inflammatory bowel disease and colitis-associated cancer through decreased levels of the inflammatory cytokines TNF-α, IL-1β, IL-6, MCP-1, and CSF-1 and of COX-2 in tumors [67]. Moreover, the tested flavonoids inhibit the STAT3/NF-κB pathway in HCT-116 colon carcinoma cells. Another flavonoid showing chemopreventive activity against colon cancer cells is quercetin. In vitro studies have shown that quercetin reduced cell viability in a dose-dependent manner in SW480 colon cancer cells [68]. Quercetin downregulated markedly Cyclin D1 and the survivin gene in a dose-dependent manner at both the transcription and protein expression levels [68]. Dysregulation of the Wnt/β-catenin pathway plays an essential role in the early stages of colorectal carcinogenesis. Park C. demonstrated that quercetin inhibited the transcriptional activity of β-catenin/Tcf in SW480 colon cancer cells [69]. In light of available literature, little is known about the anti-cancer activity of santin. Szoka et al. have shown that santin significantly reduced the proliferation, viability, and clonogenicity of gastric cancer cells line AGS, liver cancer cells line HepG2, and colon cancer cells line DLD-1 [19]. Santin activation caspases: -3, -7, -8, -9 in cancer cells lead to apoptosis. The irreversible process of cell death determines the permeability of the membrane mitochondrial and caspase activation. Our results also have shown that the co-treatment with santin plus TRAIL causes increase in mitochondrial membrane potential in SW480 and SW620 colon cancer cells.
The TRAIL’s receptors DR4 and DR5 have varying functions in different cancer cell types. The induction of apoptosis into colon cancer cells occurs with DR5 rather than DR4. Overexpression of DRs in cancer cells is one of the reasons for cell resistance to TRAIL. Taniguchi H. et al. have shown that baicalein up-regulates DR5 expression in SW480 colon cancer cells [70]. They used a small interfering RNA (siRNA) in suppression of this up-regulation, and it efficiently reduced the apoptosis induced by TRAIL and baicalein. They suggested that the sensitization was mediated through DR5 induction. Yoshida T. et al. showed that kaempferol markedly up-regulated DR4 and DR5 receptors in SW480 colon cancer cells [60]. They confirmed, as the previous researchers, that DR5 but not DR4 siRNA efficiently blocked induced apoptosis combined with kaempferol and TRAIL, and they indicated that DR5 up-regulation by kaempferol helps to enhance TRAIL action. In turn, a flavonolignan silibinin isolated from the Silybum marianum significantly increased DR4 and DR5 expressions in SW480 and SW620 cells [71]. Horinaka et al. demonstrated sensitization of colon cancer DLD-1 cells does TRAIL-mediated apoptosis by apigenin through the increased expression of DR5. A co-treatment with apigenin and TRAIL-induced the activation of caspases and Bid. Additionally, DR5/Fc chimera protein efficiently blocked the activations of Bid and caspases promoted by co-treatment with TRAIL and apigenin [72]. Ding et al. described the overcome of TRAIL-resistance colon cancer HT-29 cells by wogonin, apigenin, and chrysin by up-regulation of DR5 receptor expression [73]. Wu B. et al. have shown that luteolin alone and in combination with TRAIL significantly increased mRNA expression of death receptor 5 in non-small cell lung cancer (NSCLC) cells [74]. Apigenin also upregulated the levels of DR4 and DR5 in NSCLC cells, thus sensitizing NSCLC cells to TRAIL-induced apoptosis [75]. Min K. demonstrated that fisetin-induced DR5 expression at the transcriptional level in human renal carcinoma cells [76]. Our results have shown that santin increases the expression of death receptors DR4 and DR5 as a potential mechanism by which this flavonoid augments TRAIL-mediated apoptosis in SW480 and SW620 colon cancer cells.
5. Conclusions
Santin enhances TRAIL-mediated apoptosis mainly by upregulating the expression of death receptors TRAIL-R1 and TRAIL-R2 in colon cancer cells. Sensitization of cancer cells to TRAIL-induced apoptosis by santin can be a potential mechanism of their anti-tumor and chemopreventive activity and affect immune surveillance.
Author Contributions
Conceptualization, E.S., M.K.; data curation, E.S.; formal analysis, M.K., D.J., G.P., E.S.; funding acquisition, E.S, M.K.; investigation, M.K., D.J., G.P., E.S.; methodology, M.K., D.J., E.S.; resources, M.K., D.J., E.S.; software, M.K., D.J.; supervision, E.S.; validation, M.K., D.J., E.S.; visualization, M.K., D.J., E.S.; writing—original draft, M.K., E.S.; writing—review and editing, M.K., E.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a research grant PCN-1-200/K/2/I from the Medical University of Silesia in Katowice (Poland).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The manuscript includes all the data generated or analyzed during this study. Please contact the corresponding author to access the data presented in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as Important Molecules of Plant Interactions with the Environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jucá, M.M.; Cysne Filho, F.M.S.; de Almeida, J.C.; da S. Mesquita, D.; de M. Barriga, J.R.; Dias, K.C.F.; Barbosa, T.M.; Vasconcelos, L.C.; Leal, L.K.A.M.; Ribeiro, J.E.; et al. Flavonoids: Biological Activities and Therapeutic Potential. Nat. Prod. Res. 2018, 34, 692–705. [Google Scholar] [CrossRef] [PubMed]
- Peluso, I.; Miglio, C.; Morabito, G.; Ioannone, F.; Serafini, M. Flavonoids and Immune Function in Human: A Systematic Review. Crit. Rev. Food Sci. Nutr. 2014, 55, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Slika, H.; Mansour, H.; Wehbe, N.; Nasser, S.A.; Iratni, R.; Nasrallah, G.; Shaito, A.; Ghaddar, T.; Kobeissy, F.; Eid, A.H. Therapeutic Potential of Flavonoids in Cancer: ROS-Mediated Mechanisms. Biomed. Pharmacother. 2022, 146, 112442. [Google Scholar] [CrossRef]
- Fernandes, I.; Pérez-Gregorio, R.; Soares, S.; Mateus, N.; De Freitas, V.; Santos-Buelga, C.; Feliciano, A.S. Wine Flavonoids in Health and Disease Prevention. Molecules 2017, 22, 292. [Google Scholar] [CrossRef]
- Ohishi, T.; Fukutomi, R.; Shoji, Y.; Goto, S.; Isemura, M. The Beneficial Effects of Principal Polyphenols from Green Tea, Coffee, Wine, and Curry on Obesity. Molecules 2021, 26, 453. [Google Scholar] [CrossRef]
- Tronina, T.; Popłó, J.; Bartmá, A. Flavonoids as Phytoestrogenic Components of Hops and Beer. Molecules 2020, 25, 4201. [Google Scholar] [CrossRef]
- Toh, J.Y.; Tan, V.M.H.; Lim, P.C.Y.; Lim, S.T.; Chong, M.F.F. Flavonoids from Fruit and Vegetables: A Focus on Cardiovascular Risk Factors. Curr. Atheroscler. Rep. 2013, 15, 368. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant Flavonoids: Classification, Distribution, Biosynthesis, and Antioxidant Activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.C.G.A.; Silva, A.M.S.; Giovinazzo, G.; Gerardi, C.; Mosca, L. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Moloney, J.N.; Cotter, T.G. ROS Signalling in the Biology of Cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef]
- Jeong, S.H.; Kim, H.H.; Ha, S.E.; Park, M.Y.; Bhosale, P.B.; Abusaliya, A.; Park, K.I.; Heo, J.D.; Jeong, H.; Kim, H.H.; et al. Citation: Flavones: Six Selected Flavones and Their Related Signaling Pathways That Induce Apoptosis in Cancer. Int. J. Mol. Sci. 2022, 23, 965. [Google Scholar] [CrossRef]
- Jiang, N.; Doseff, A.I.; Grotewold, E. Plants Flavones: From Biosynthesis to Health Benefits. Plants 2016, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food Sources, Bioavailability, Metabolism, and Bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Rechner, A.R.; Smith, M.A.; Kuhnle, G.; Gibson, G.R.; Debnam, E.S.; Srai, S.K.S.; Moore, K.P.; Rice-Evans, C.A. Colonic Metabolism of Dietary Polyphenols: Influence of Structure on Microbial Fermentation Products. Free Radic. Biol. Med. 2004, 36, 212–225. [Google Scholar] [CrossRef]
- Long, C.; Sauleau, P.; David, B.; Lavaud, C.; Cassabois, V.V.; Frédé, F.; Ausseil, F.; Massiot, G. Bioactive Flavonoids of Tanacetum Parthenium Revisited. Phytochemistry 2003, 64, 567–569. [Google Scholar] [CrossRef]
- Martinez, J.; Silvá, A.M.; Abad, M.J.; Bermejo, P.; Villar, A.; Söllhuber, M. Isolation of Two Flavonoids from Tanacetum Microphyllum as PMA-Induced Ear Edema Inhibitors. J. Nat. Prod. 1997, 60, 142–144. [Google Scholar] [CrossRef]
- Melaku, Y.; Worku, T.; Tadesse, Y.; Mekonnen, Y.; Schmidt, J.; Arnold, N.; Dagne, E. Antiplasmodial Compounds from Leaves of Dodonaea Angustifolia. Curr. Bioact. Compd. 2017, 13, 268–273. [Google Scholar] [CrossRef] [Green Version]
- Szoka, L.; Nazaruk, J.; Stocki, M.; Isidorov, V. Santin and Cirsimaritin from Betula Pubescens and Betula Pendula Buds Induce Apoptosis in Human Digestive System Cancer Cells. J. Cell Mol. Med. 2021, 25, 11085–11096. [Google Scholar] [CrossRef]
- Smeriglio, A.; Raudone, L.; Radušiene, J.; Seyis, F.; Yayla, F.; Marksa, M.; Ivanauskas, L.; Cırak, C. Distribution of Phenolic Compounds and Antioxidant Activity in Plant Parts and Populations of Seven Underutilized Wild Achillea Species. Plants 2022, 11, 447. [Google Scholar] [CrossRef]
- Pitti, R.M.; Marsters, S.A.; Ruppert, S.; Donahue, C.J.; Moore, A.; Ashkenazi, A. Induction of Apoptosis by Apo-2 Ligand, a New Member of the Tumor Necrosis Factor Cytokine Family. J. Biol. Chem. 1996, 271, 12687–12690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiley, S.R.; Schooley, K.; Smolak, P.J.; Din, W.S.; Huang, C.P.; Nicholl, J.K.; Sutherland, G.R.; Smith, T.D.; Rauch, C.; Smith, C.A.; et al. Identification and Characterization of a New Member of the TNF Family That Induces Apoptosis. Immunity 1995, 3, 673–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falschlehner, C.; Schaefer, U.; Walczak, H. Following TRAIL’s Path in the Immune System. Immunology 2009, 127, 145. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A. Targeting Death and Decoy Receptors of the Tumour-Necrosis Factor Superfamily. Nat. Rev. Cancer 2002, 2, 420–430. [Google Scholar] [CrossRef]
- Emery, J.G.; McDonnell, P.; Burke, M.B.; Deen, K.C.; Lyn, S.; Silverman, C.; Dul, E.; Appelbaum, E.R.; Eichman, C.; DiPrinzio, R.; et al. Osteoprotegerin Is a Receptor for the Cytotoxic Ligand TRAIL. J. Biol. Chem. 1998, 273, 14363–14367. [Google Scholar] [CrossRef] [Green Version]
- Gonzalvez, F.; Ashkenazi, A. New Insights into Apoptosis Signaling by Apo2L/TRAIL. Oncogene 2010, 29, 4752–4765. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Lin, J.; Xu, R. The Molecular Mechanisms of TRAIL Resistance in Cancer Cells: Help in Designing New Drugs. Curr. Pharm. Des. 2014, 20, 6714–6722. [Google Scholar] [CrossRef]
- Kłósek, M.; Mertas, A.; Król, W.; Jaworska, D.; Szymszal, J.; Szliszka, E. Molecular Sciences Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand-Induced Apoptosis in Prostate Cancer Cells after Treatment with Xanthohumol-A Natural Compound Present in Humulus lupulus L. Int. J. Mol. Sci. 2016, 17, 837. [Google Scholar] [CrossRef] [Green Version]
- Szliszka, E.; Jaworska, D.; Kłósek, M.; Czuba, Z.P.; Król, W. Targeting Death Receptor TRAIL-R2 by Chalcones for TRAIL-Induced Apoptosis in Cancer Cells. Int. J. Mol. Sci. 2012, 13, 15343–15359. [Google Scholar] [CrossRef] [Green Version]
- Bronikowska, J.; Szliszka, E.; Kostrzewa-Susłow, E.; Jaworska, D.; Czuba, Z.P.; Bednarski, P.; Król, W. Novel Structurally Related Flavones Augment Cell Death Induced by RhsTRAIL. Int. J. Mol. Sci. 2017, 18, 1211. [Google Scholar] [CrossRef] [Green Version]
- Bronikowska, J.; Szliszka, E.; Czuba, Z.P.; Zwolinski, D.; Szmydki, D.; Krol, W. The Combination of TRAIL and Isoflavones Enhances Apoptosis in Cancer Cells. Molecules 2010, 15, 2000–2015. [Google Scholar] [CrossRef] [Green Version]
- Bronikowska, J.; Kłósek, M.; Janeczko, T.; Kostrzewa-Susłow, E.; Czuba, Z.P. The Modulating Effect of Methoxy-Derivatives of 2′-Hydroxychalcones on the Release of IL-8, MIF, VCAM-1 and ICAM-1 by Colon Cancer Cells. Biomed. Pharmacother. 2022, 145, 112428. [Google Scholar] [CrossRef]
- Jaworska, D.; Szliszka, E. Targeting Apoptotic Activity Against Prostate Cancer Stem Cells. Int. J. Mol. Sci. 2017, 18, 1648. [Google Scholar] [CrossRef] [Green Version]
- Aoi, W.; Iwasa, M.; Marunaka, Y. Metabolic Functions of Flavonoids: From Human Epidemiology to Molecular Mechanism. Neuropeptides 2021, 88, 102163. [Google Scholar] [CrossRef]
- Abotaleb, M.; Mathews Samuel, S.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Review Flavonoids in Cancer and Apoptosis. Cancers 2019, 11, 28. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-García, C.; Sánchez-Quesada, C.; Gaforio, J.J. Dietary Flavonoids as Cancer Chemopreventive Agents: An Updated Review of Human Studies. Antioxidants 2019, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Chirumbolo, S.; Bjørklund, G.; Lysiuk, R.; Vella, A.; Lenchyk, L.; Upyr, T. Molecular Sciences Targeting Cancer with Phytochemicals via Their Fine Tuning of the Cell Survival Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 3568. [Google Scholar] [CrossRef] [Green Version]
- Raffa, D.; Maggio, B.; Raimondi, M.V.; Plescia, F.; Daidone, G. Recent Discoveries of Anticancer Flavonoids. Eur. J. Med. Chem. 2017, 142, 213–228. [Google Scholar] [CrossRef]
- Ponte, L.G.S.; Pavan, I.C.B.; Mancini, M.C.S.; Da Silva, L.G.S.; Morelli, A.P.; Severino, M.B.; Bezerra, R.M.N.; Simabuco, F.M. The Hallmarks of Flavonoids in Cancer. Molecules 2021, 26, 2029. [Google Scholar] [CrossRef]
- Neagu, M.; Constantin, C.; Popescu, I.D.; Zipeto, D.; Tzanakakis, G.; Nikitovic, D.; Fenga, C.; Stratakis, C.A.; Spandidos, D.A.; Tsatsakis, A.M. Inflammation and Metabolism in Cancer Cell—Mitochondria Key Player. Front. Oncol. 2019, 9, 348. [Google Scholar] [CrossRef] [Green Version]
- Sandoval-Acuña, C.; Ferreira, J.; Speisky, H. Polyphenols and Mitochondria: An Update on Their Increasingly Emerging ROS-Scavenging Independent Actions. Arch. Biochem. Biophys. 2014, 559, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Tavsan, Z.; Kayali, H.A. Flavonoids Showed Anticancer Effects on the Ovarian Cancer Cells: Involvement of Reactive Oxygen Species, Apoptosis, Cell Cycle and Invasion. Biomed. Pharmacother. 2019, 116, 109004. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lim, W.; Bazer, F.W.; Song, G. Naringenin Suppresses Growth of Human Placental Choriocarcinoma via Reactive Oxygen Species-Mediated P38 and JNK MAPK Pathways. Phytomedicine 2018, 50, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Palit, S.; Kar, S.; Sharma, G.; Das, P.K. Hesperetin Induces Apoptosis in Breast Carcinoma by Triggering Accumulation of ROS and Activation of ASK1/JNK Pathway. J. Cell Physiol. 2015, 230, 1729–1739. [Google Scholar] [CrossRef]
- Jin, S.; Zhang, Q.Y.; Kang, X.M.; Wang, J.X.; Zhao, W.H. Daidzein Induces MCF-7 Breast Cancer Cell Apoptosis via the Mitochondrial Pathway. Ann. Oncol. 2010, 21, 263–268. [Google Scholar] [CrossRef]
- SALMANI, J.M.M.; ZHANG, X.P.; JACOB, J.A.; CHEN, B.A. Apigenin’s Anticancer Properties and Molecular Mechanisms of Action: Recent Advances and Future Prospectives. Chin. J. Nat. Med. 2017, 15, 321–329. [Google Scholar] [CrossRef]
- Srivastava, R.K. TRAIL/Apo-2L: Mechanisms and Clinical Applications in Cancer. Neoplasia 2001, 3, 535–546. [Google Scholar] [CrossRef] [Green Version]
- McArthur, K.; Kile, B.T. Apoptotic Caspases: Multiple or Mistaken Identities? Trends Cell Biol. 2018, 28, 475–493. [Google Scholar] [CrossRef]
- Mohan Shankar, G.; Swetha, M.; Keerthana, C.K.; Rayginia, T.P.; Anto, R.J. Cancer Chemoprevention: A Strategic Approach Using Phytochemicals. Front. Pharmacol. 2022, 12, 809308. [Google Scholar] [CrossRef]
- Chang, H.; Lei, L.; Zhou, Y.; Ye, F.; Zhao, G. Dietary Flavonoids and the Risk of Colorectal Cancer: An Updated Meta-Analysis of Epidemiological Studies. Nutrients 2018, 10, 950. [Google Scholar] [CrossRef] [Green Version]
- Woo, H.D.; Kim, J. Dietary Flavonoid Intake and Risk of Stomach and Colorectal Cancer. World J. Gastroenterol. 2013, 19, 1011–1019. [Google Scholar] [CrossRef]
- Mansuri, M.L.; Parihar, P.; Solanki, I.; Parihar, M.S. Flavonoids in Modulation of Cell Survival Signalling Pathways. Genes Nutr. 2014, 9, 400. [Google Scholar] [CrossRef]
- Beyer, K.; Baukloh, A.K.; Stoyanova, A.; Kamphues, C.; Sattler, A.; Kotsch, K. Interactions of Tumor Necrosis Factor–Related Apoptosis-Inducing Ligand (TRAIL) with the Immune System: Implications for Inflammation and Cancer. Cancers 2019, 11, 1161. [Google Scholar] [CrossRef] [Green Version]
- Malki, A.; Elruz, R.A.; Gupta, I.; Allouch, A.; Vranic, S.; Al Moustafa, A.E. Molecular Mechanisms of Colon Cancer Progression and Metastasis: Recent Insights and Advancements. Int. J. Mol. Sci. 2021, 22, 130. [Google Scholar] [CrossRef]
- Jung, Y.H.; Heo, J.; Lee, Y.J.; Kwon, T.K.; Kim, Y.H. Quercetin Enhances TRAIL-Induced Apoptosis in Prostate Cancer Cells via Increased Protein Stability of Death Receptor 5. Life Sci. 2010, 86, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Nazim, U.M.D.; Park, S.Y. Luteolin Sensitizes Human Liver Cancer Cells to TRAIL-induced Apoptosis via Autophagy and JNK-mediated Death Receptor 5 Upregulation. Int. J. Oncol. 2019, 54, 665–672. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Tian, B.; Wang, Y.; Ding, H. Kaempferol Sensitizes Human Ovarian Cancer Cells-OVCAR-3 and SKOV-3 to Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL)-Induced Apoptosis via JNK/ERK-CHOP Pathway and Up-Regulation of Death Receptors 4 and 5. Med. Sci. Monit. 2017, 23, 5096. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.-Y.; Park, C.; Hwang, H.J.; Kim, G.-Y.; Tae Choi, B.; Kim, W.-J.; Choi, Y.H. Naringenin Up-Regulates the Expression of Death Receptor 5 and Enhances TRAIL-Induced Apoptosis in Human Lung Cancer A549 Cells. Mol. Nutr. 2011, 55, 300–309. [Google Scholar] [CrossRef]
- Kłósek, M.; Kuropatnicki, A.K.; Szliszka, E.; Korzonek-Szlacheta, I.; Król, W. Chalcones Target the Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL) Signaling Pathway for Cancer Chemoprevention; Elsevier: Amsterdam, The Netherlands, 2017; ISBN 9780128092996. [Google Scholar] [CrossRef]
- Yoshida, T.; Konishi, M.; Horinaka, M.; Yasuda, T.; Goda, A.E.; Taniguchi, H.; Yano, K.; Wakada, M.; Sakai, T. Kaempferol Sensitizes Colon Cancer Cells to TRAIL-Induced Apoptosis. Biochem. Biophys. Res. Commun. 2008, 375, 129–133. [Google Scholar] [CrossRef]
- Tang, S.Y.; Zhong, M.Z.; Yuan, G.J.; Hou, S.P.; Yin, L.L.; Jiang, H.; Yu, Z.Y. Casticin, a Flavonoid, Potentiates TRAIL-Induced Apoptosis through Modulation of Anti-Apoptotic Proteins and Death Receptor 5 in Colon Cancer Cells. Oncol. Rep. 2013, 29, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Z.; Zhang, C.F.; Chen, L.; Anderson, S.; Lu, F.; Yuan, C.S. Colon Cancer Chemopreventive Effects of Baicalein, an Active Enteric Microbiome Metabolite from Baicalin. Int. J. Oncol. 2015, 47, 1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, T.K.; Kim, M.E.; Yoon, J.H.; Bae, S.J.; Yeom, J.; Lee, J.S. Galangin Induces Human Colon Cancer Cell Death via the Mitochondrial Dysfunction and Caspase-Dependent Pathway. Exp. Biol. Med. 2013, 238, 1047–1054. [Google Scholar] [CrossRef]
- Xu, M.; Wang, S.; Song, Y.; Yao, J.; Huang, K.; Zhu, X. Apigenin Suppresses Colorectal Cancer Cell Proliferation, Migration and Invasion via Inhibition of the Wnt/β-Catenin Signaling Pathway. Oncol. Lett. 2016, 11, 3075–3080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chunhua, L.; Donglan, L.; Xiuqiong, F.; Lihua, Z.; Qin, F.; Yawei, L.; Liang, Z.; Ge, W.; Linlin, J.; Ping, Z.; et al. Apigenin Up-Regulates Transgelin and Inhibits Invasion and Migration of Colorectal Cancer through Decreased Phosphorylation of AKT. J. Nutr. Biochem. 2013, 24, 1766–1775. [Google Scholar] [CrossRef] [PubMed]
- Márquez-Flores, Y.K.; Villegas, I.; Cárdeno, A.; Rosillo, M.Á.; Alarcón-de-la-Lastra, C. Apigenin Supplementation Protects the Development of Dextran Sulfate Sodium-Induced Murine Experimental Colitis by Inhibiting Canonical and Non-Canonical Inflammasome Signaling Pathways. J. Nutr. Biochem. 2016, 30, 143–152. [Google Scholar] [CrossRef]
- Ai, X.Y.; Qin, Y.; Liu, H.J.; Cui, Z.H.; Li, M.; Yang, J.H.; Zhong, W.L.; Liu, Y.R.; Chen, S.; Sun, T.; et al. Apigenin Inhibits Colonic Inflammation and Tumorigenesis by Suppressing STAT3-NF-ΚB Signaling. Oncotarget 2017, 8, 100216. [Google Scholar] [CrossRef] [Green Version]
- Shan, B.E.; Wang, M.X.; Li, R.Q. Quercetin Inhibit Human SW480 Colon Cancer Growth in Association with Inhibition of Cyclin D1 and Survivin Expression through Wnt/Beta-Catenin Signaling Pathway. Cancer Investig. 2009, 27, 604–612. [Google Scholar] [CrossRef]
- Park, C.H.; Chang, J.Y.; Hahm, E.R.; Park, S.; Kim, H.K.; Yang, C.H. Quercetin, a Potent Inhibitor against β-Catenin/Tcf Signaling in SW480 Colon Cancer Cells. Biochem. Biophys. Res. Commun. 2005, 328, 227–234. [Google Scholar] [CrossRef]
- Taniguchi, H.; Yoshida, T.; Horinaka, M.; Yasuda, T.; Goda, A.E.; Konishi, M.; Wakada, M.; Kataoka, K.; Yoshikawa, T.; Sakai, T. Baicalein Overcomes Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand Resistance via Two Different Cell-Specific Pathways in Cancer Cells but Not in Normal Cells. Cancer Res 2008, 68, 8918–8945. [Google Scholar] [CrossRef] [Green Version]
- Kauntz, H.; Bousserouel, S.; Gossé, F.; Raul, F. Silibinin Triggers Apoptotic Signaling Pathways and Autophagic Survival Response in Human Colon Adenocarcinoma Cells and Their Derived Metastatic Cells. Apoptosis 2011, 16, 1042–1053. [Google Scholar] [CrossRef]
- Horinaka, M.; Yoshida, T.; Shiraishi, T.; Nakata, S.; Wakada, M.; Sakai, T. The Dietary Flavonoid Apigenin Sensitizes Malignant Tumor Cells to Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand. Mol. Cancer Ther. 2006, 5, 945–951. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Polier, G.; Koḧler, R.; Giaisi, M.; Krammer, P.H.; Li-Weber, M. Wogonin and Related Natural Flavones Overcome Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL) Protein Resistance of Tumors by down-Regulation of c-FLIP Protein and up-Regulation of TRAIL Receptor 2 Expression. J. Biol. Chem. 2012, 287, 641–649. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Xiong, J.; Zhou, Y.; Wu, Y.; Song, Y.; Wang, N.; Chen, L.; Zhang, J. Luteolin Enhances TRAIL Sensitivity in Non-Small Cell Lung Cancer Cells through Increasing DR5 Expression and Drp1-Mediated Mitochondrial Fission. Arch. Biochem. Biophys. 2020, 692, 108539. [Google Scholar] [CrossRef]
- Chen, M.; Wang, X.; Zha, D.; Cai, F.; Zhang, W.; He, Y.; Huang, Q.; Zhuang, H.; Hua, Z.C. Apigenin Potentiates TRAIL Therapy of Non-Small Cell Lung Cancer via Upregulating DR4/DR5 Expression in a P53-Dependent Manner. Sci. Rep. 2016, 6, 35468. [Google Scholar] [CrossRef] [Green Version]
- Min, K.J.; Nam, J.O.; Kwon, T.K. Fisetin Induces Apoptosis through P53-Mediated up-Regulation of Dr5 Expression in Human Renal Carcinoma Caki Cells. Molecules 2017, 22, 1285. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Chemical structure of santin (5,7-dihydroxy-3,6,4′-trimetoxy-flavone).

Figure 2.
Santin enhanced TRAIL-induced apoptosis in colon cancer cells. SW480 and SW620 cells were incubated for 48 h with rhsTRAIL at the concentrations of 25–100 ng/mL and/or with 25–100 μM santin. The percentage of apoptotic cells was determined by flow cytometry using annexin V-FITC staining. The values represent mean ± SD of three independent experiments performed fourfold (n = 3) (*** p < 0.001 compared with control, +++ p < 0.001 compared with santin, ### p < 0.001 compared with TRAIL).
Figure 2.
Santin enhanced TRAIL-induced apoptosis in colon cancer cells. SW480 and SW620 cells were incubated for 48 h with rhsTRAIL at the concentrations of 25–100 ng/mL and/or with 25–100 μM santin. The percentage of apoptotic cells was determined by flow cytometry using annexin V-FITC staining. The values represent mean ± SD of three independent experiments performed fourfold (n = 3) (*** p < 0.001 compared with control, +++ p < 0.001 compared with santin, ### p < 0.001 compared with TRAIL).

Figure 3.
Effects of santin on the expression of TRAIL-R1 and TRAIL-R2 death receptors in SW480 and SW620 cancer cells. Cells were incubated for 24 h with santin at concentrations of 50 μM and 100 μM. The surface expression of death receptors on cancer cells was measured by flow cytometry.
Figure 3.
Effects of santin on the expression of TRAIL-R1 and TRAIL-R2 death receptors in SW480 and SW620 cancer cells. Cells were incubated for 24 h with santin at concentrations of 50 μM and 100 μM. The surface expression of death receptors on cancer cells was measured by flow cytometry.

Figure 4.
Effects of santin on the expression of TRAIL-R1 and TRAIL-R2 death receptors in SW480 and SW620 cancer cells. The values represent the mean ± SD of three independent experiments performed in duplicate (n = 3) (** p < 0.01, *** p < 0.001 compared with control).
Figure 4.
Effects of santin on the expression of TRAIL-R1 and TRAIL-R2 death receptors in SW480 and SW620 cancer cells. The values represent the mean ± SD of three independent experiments performed in duplicate (n = 3) (** p < 0.01, *** p < 0.001 compared with control).

Figure 5.
The effects of TRAIL combined with santin on the mitochondrial membrane potential (ΔΨm) in colon cancer cells. SW480 and SW620 cells were subject to incubation for 48 h with rhsTRAIL (concentration of 25–100 ng/mL) and/or with santin (25–100 μM). The fluorescent microscopic analysis of DePsipher staining was used to assess the ΔΨm loss in cancer cells (*** p < 0.001 compared with control, +++ p < 0.001 compared with santin, ### p < 0.001 compared with TRAIL).
Figure 5.
The effects of TRAIL combined with santin on the mitochondrial membrane potential (ΔΨm) in colon cancer cells. SW480 and SW620 cells were subject to incubation for 48 h with rhsTRAIL (concentration of 25–100 ng/mL) and/or with santin (25–100 μM). The fluorescent microscopic analysis of DePsipher staining was used to assess the ΔΨm loss in cancer cells (*** p < 0.001 compared with control, +++ p < 0.001 compared with santin, ### p < 0.001 compared with TRAIL).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kłósek, M.; Jaworska, D.; Pietsz, G.; Szliszka, E. Santin (5,7-Dihydroxy-3,6,4′-Trimetoxy-Flavone) Enhances TRAIL-Mediated Apoptosis in Colon Cancer Cells. Life 2023, 13, 592. https://doi.org/10.3390/life13020592
AMA Style
Kłósek M, Jaworska D, Pietsz G, Szliszka E. Santin (5,7-Dihydroxy-3,6,4′-Trimetoxy-Flavone) Enhances TRAIL-Mediated Apoptosis in Colon Cancer Cells. Life. 2023; 13(2):592. https://doi.org/10.3390/life13020592
Chicago/Turabian StyleKłósek, Małgorzata, Dagmara Jaworska, Grażyna Pietsz, and Ewelina Szliszka. 2023. "Santin (5,7-Dihydroxy-3,6,4′-Trimetoxy-Flavone) Enhances TRAIL-Mediated Apoptosis in Colon Cancer Cells" Life 13, no. 2: 592. https://doi.org/10.3390/life13020592
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.