
Ruta chalepensis L. In Vitro Cultures as a Source of Bioactive Furanocoumarins and Furoquinoline Alkaloids



1
Department of Pharmaceutical Botany, Faculty of Pharmacy, Jagiellonian University Medical College, 30-688 Krakow, Poland
2
SSG of Medicinal Plants and Mushroom Biotechnology Department of Pharmaceutical Botany, Jagiellonian University Medical College, 30-688 Krakow, Poland
*
Author to whom correspondence should be addressed.
Life 2023, 13(2), 457; https://doi.org/10.3390/life13020457
Submission received: 5 January 2023
/
Revised: 30 January 2023
/
Accepted: 1 February 2023
/
Published: 6 February 2023
(This article belongs to the Collection The Role of Bioactive Natural Compounds in the Treatment of Diseases)
Abstract
:Simple Summary
Of late, the interest in alternative sources of bioactive compounds to field crops has been increasing due to decreasing crop acreages and climate change. Plant biotechnology offers the possibility of cultivating valuable medicinal plant species using in vitro methods. In vitro cultures are characterized by a significant increase in biomass, and one of their primary advantages is that they are independent of climate and soil conditions. This study aimed to investigate the ability of Ruta chalepensis bioreactor cultures to produce the following valuable bioactive substances: linear furanocoumarins and furoquinoline alkaloids. Bioreactor cultures of R. chalepensis may be used as a good raw material for the production of important bioactive compounds from the groups of coumarins and alkaloids.
Abstract
Recently, due to the decreasing areas of cultivation and climate change, the use of biotechnological methods to obtain biomass, which is a source of valuable bioactive metabolites, is becoming more and more interesting. In this study, Ruta chalepensis in vitro cultures were investigated in RITA® temporary immersion bioreactors. Biomass growth and the production of secondary metabolites in 4- and 5-week growth cycles on three variants of the Linsmaier and Skoog (LS) medium (naphthyl-1-acetic acid/6-benzylaminopurine (NAA/BAP): 0.5/1.0, 0.1/0.1, and 1.0/1.0 mg/L) were analyzed. Using high-performance liquid chromatography of methanolic extracts of biomass, the presence of linear furanocoumarins (bergapten, isoimperatorin, isopimpinellin, psoralen, and xanthotoxin) and furoquinoline alkaloids (γ-fagarine, 7-isopentenyloxy-γ-fagarine, and skimmianine) was confirmed. The highest content of linear furanocoumarins (1170 mg/100 g DW (dry weight)) was observed in the LS medium variant containing 0.5/1.0 mg/L NAA/BAP (4-week growth cycle). The highest content of furoquinoline alkaloids (449 mg/100 g DW) was observed in the LS medium variant containing 0.1/0.1 mg/L NAA/BAP (5-week growth cycle). Hence, R. chalepensis bioreactor cultures may be used as a biotechnological source of linear furanocoumarins (xanthotoxin and bergapten) and furoquinoline alkaloids (skimmianine and γ-fagarine).
1. Introduction
Besides Ruta graveolens, the most valuable species of the Ruta genus is Ruta chalepensis. R. chalepensis L. (Rutaceae) is native to the Mediterranean region but widely distributed and cultivated worldwide in temperate and tropical climates, e.g., Latin America, Africa (Tunisia, Algeria, Libya), and Asia (Israel, Iran, Turkey, Syria) [1,2,3]. R. chalepensis contains secondary metabolites from various chemical groups as follows: alkaloids (acridine, quinoline, furoquinoline alkaloids), flavonoids, coumarins (furanocoumarins, dihydrofuranocoumarins, coumarin dimers, simple coumarins), and essential oils (primary compounds: nonan-2-one and undecan-2-one) [4,5,6,7]. Ruta is one of the most frequently used plants for medicinal purposes [1]. R. chalepensis exhibits a wide range of healing effects: antioxidant, anti-inflammatory, antipyretic, spasmolytic, sedative, analgesic, antibacterial, antifungal, cytotoxic, and antidiabetic effects, to name a few [8,9,10,11,12,13,14]. The herb of R. chalepensis has linear furanocoumarins (xanthotoxin and bergapten) that are used in dermatology. Furanocoumarins are responsible for photoreactive properties, so they are used as pigmentation stimulators and antiproliferative factors. The so-called PUVA therapy (psoralens and UVA irradiation) is a kind of photochemotherapy used in the treatment of vitiligo, psoriasis, mycosis, and atopic eczema [15,16,17,18]. The use of furanocoumarins needs to be closely monitored as they can also lead to side effects (photodermatoses and kidney and liver damage) [18]. There are also reports of other applications of furanocoumarins. Bergapten shows activity as a selective blocker of axolemmal potassium channels and may be useful in the treatment of demyelinating diseases [1,19]. Furanocoumarins exhibit anticancer activity and are currently considered novel mechanism-based drug candidates for the treatment of selected forms of cancer [20]. Commercial production of furanocoumarins via chemical synthesis is quite expensive, and they are obtained from bergamot oil (Citrus bergamia). Unfortunately, the areas of cultivation of this plant are gradually decreasing. Hence, there is a need for new sources of these compounds. The genus Ruta is a promising source, but scaling up field crops is similarly problematic [19]. Species of the genus Ruta are also rich in furoquinoline alkaloids (γ-fagarine and skimmianine). Furoquinoline alkaloids have antifungal and antibacterial activities, and acetylcholinesterase-inhibiting properties, as well as an inhibitory effect on the 5-HT2 receptor [21,22].
In vitro cultures can be used as a source of secondary metabolites. In vitro plant cultures have a significant potential for the commercial production of secondary metabolites. They allow for the synthesis of bioactive metabolites under controlled conditions, thus making producers independent of climate and soil conditions. They can be carried out continuously, and the biomass obtained is homogenous. Cell lines with favorable characteristics, e.g., high production of selected metabolites, can be selected, which are a valuable and reliable source of secondary metabolites [23,24]. The production of secondary metabolites in in vitro cultures is also associated with a few problems. The major problem is scaling up for industrial production. On the one hand, shoot cultures are the most productive as they retain a high degree of tissue differentiation; on the other hand, carrying out this type of culture on a larger scale becomes difficult. Temporary immersion system bioreactors are extremely useful for this purpose. They allow for the rapid multiplication of biomass and are easy to use. Our previous studies on in vitro agitated shoot cultures of R. chalepensis confirmed the high biosynthetic potential of these cultures in terms of linear furanocoumarins and furoquinoline alkaloids [25].
With the encouraging findings of these studies, this study attempted to scale up the cultures carried out using the temporary immersion system and assess the usefulness of this method for the commercial production of the abovementioned metabolites. This type of culture is conducted for the first time for the species R. chalepensis.
2. Materials and Methods
2.1. In Vitro Cultures
The starting material used in this study was the in vitro cultures of R. chalepensis, established in 2018 from the seeds obtained from the Botanical Garden of Maria Skłodowska-Curie University, Lublin, Poland. The initial cultures were carried out as liquid stationary cultures on the Linsmaier and Skoog (LS) medium [26] and the following plant growth and development regulators: naphthyl-1-acetic acid (NAA) and 6-benzylaminopurine (BAP) (1.0/1.0 mg/L).
R. chalepensis shoot cultures were carried out in RITA® bioreactors (VITROPIC, Saint-Mathieu-de-Treviers, France). Of the previously grown plant biomass, 5.0 g was placed in the bioreactors, to which 200 mL of the LS medium was added. Growth and development regulators in three concentration variants (NAA/BAP: 0.5/1.0, 0.1/0.1, and 1.0/1.0 mg/L) were then added to it. Immersion frequency was 5 min every 90 min. Cultivation was carried out for a period of 4 and 5 weeks. Finally, fresh biomass was collected and dried at approximately 38 °C.
2.2. Extraction
Micronized dry biomass (1.0 g) was weighed into 250 mL round-bottom flasks. Biomass was then extracted with methanol (50 mL) at the solvent boiling point (64.7 °C). Following this, the extracts were evaporated, dissolved in 4.0 mL high-performance liquid chromatography (HPLC)-grade methanol, and filtered through Millipore filters (pore size 0.22 µm) for the HPLC analysis.
2.3. HPLC Analyses
HPLC analysis was carried out as previously described [27] on a liquid chromatograph (LaChrom Elite, Hitachi, Tokyo, Japan) using DAD detection (200–400 nm). Quantification was carried out at two wavelengths (λ): 254 and 330 nm.
Reference substances were obtained from bergapten, imperatorin, xanthotoxin, and psoralen from Roth (Karlsruhe, Germany); 5,7-dimethoxycoumarin, 4-hydroxy-6-methylcoumarin, 6-methylcoumarin, osthole, and umbelliferone from Sigma-Aldrich (St Louis, MO, USA); coumarin and scopoletin from Fluka (Bucha, Switzerland); 4-methylumbelliferone, 4,6-dimethoxy-2H-1-benzopyran-2-one, and skimmianine from ChromaDex (Irvine, CA, USA); and isopimpinellin, isoimperatorin, daphnetin 7-methyl ether, rutaretin, daphnetin, osthenol, bergaptol, daphnetin dimethyl ether, γ-fagarine, and 7-isopentenyloxy-γ-fagarine from ChemFaces (Wuhan, China).
2.4. Statistical Analysis
Statistical analysis of the results was carried out using analysis of variance. The NIR post-hoc test was used, and the comparison of homogeneous groups was made, with two independent variables (growth cycle and medium variant). STATISTICA software (version 13.3, StatSoft Inc., Tulsa, OK, USA) was used. Detailed results of the statistical analysis are included in the Supplementary Materials.
3. Results
3.1. In Vitro Cultures
The in vitro cultures of R. chalepensis were grown as shoot cultures in 4- and 5-week growth cycles on three variants of the LS medium (LS NAA/BAP 0.5/1.0, 0.1/0.1, and 1.0/1.0) in RITA® bioreactors. They were characterized by intense green color and grew primarily in the form of shoots with a slight tendency to form callus tissue (Figure 1). The increase in biomass in all cultures was similar. Their dry biomass content ranged between 3.97 (the LS medium containing 0.1/0.1 mg/L NAA/BAP, 4-week growth cycle) and 3.17 g (the LS medium containing 1.0/1.0 mg/L NAA/BAP, 4-week growth cycle). The highest weight was obtained after the 4-week cultivation period (growth cycle) on the LS medium containing 0.1/0.1 mg/L NAA/BAP. No significant differences in biomass growth were observed between 4- and 5-week growth cycles. The obtained dry biomass content is summarized in Table 1.
3.2. HPLC Analyses
3.2.1. Production of Linear Furanocoumarins
HPLC analyses of methanol extracts from the biomass of R. chalepensis bioreactor cultures confirmed the presence of five linear furanocoumarins: bergapten, isoimperatorin, isopimpinellin, psoralen, and xanthotoxin. A sample HPLC chromatogram of the culture extract is included in Figure S1 of the Supplementary Materials.
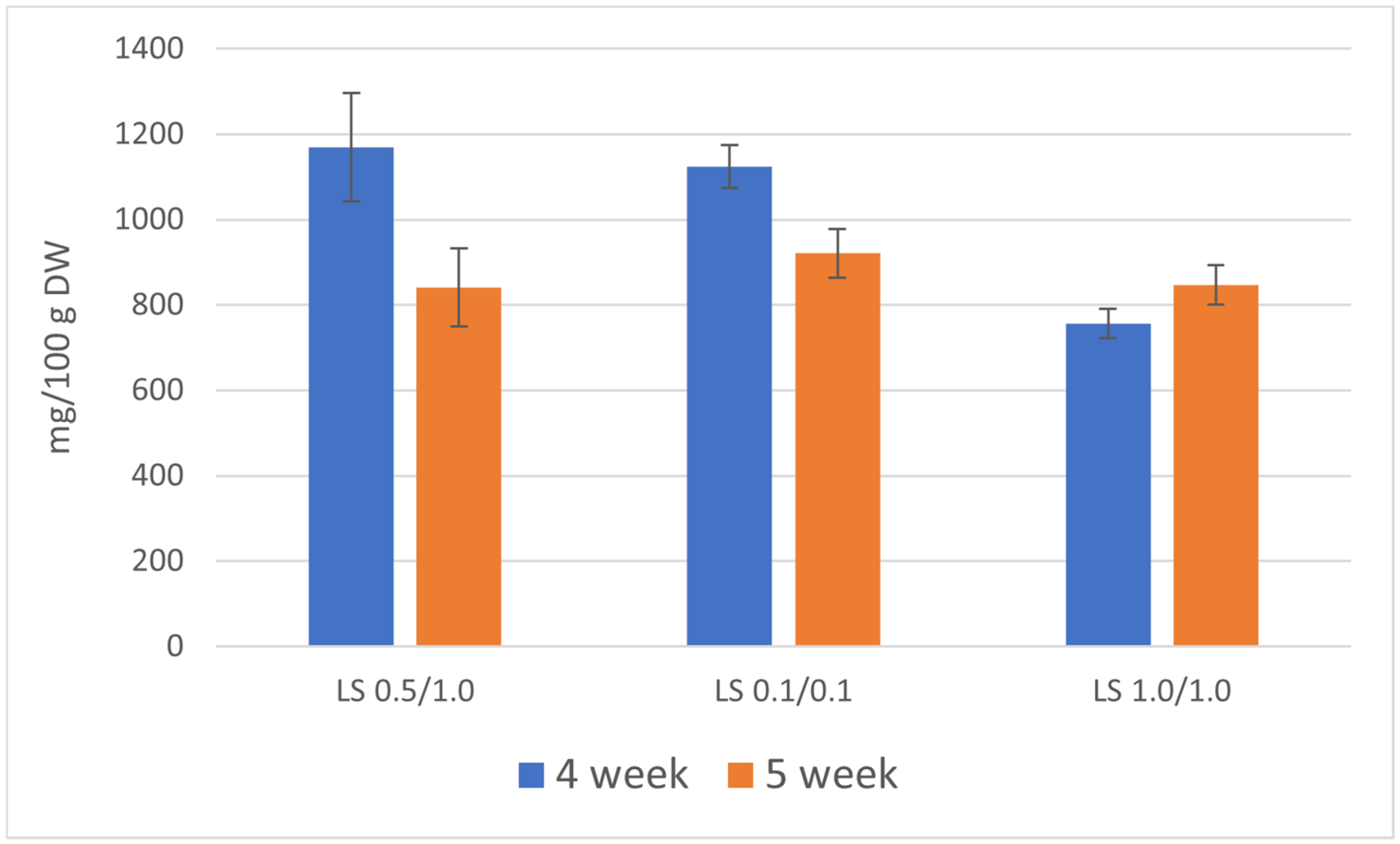
The total content of five linear furanocoumarins determined in the culture extracts during the 4-week growth cycle was quite higher in comparison with the 5-week growth cycle and ranged between 756 and 1170 mg/100 g DW (dry weight). This higher accumulation of furanocoumarins was attributable to the use of a medium containing lower concentrations of plant growth and development regulators (NAA/BAP 0.5/1.0 and 0.1/0.1 mg/L). The primary compound was xanthotoxin, with the highest content of 604 mg/100 g DW on the LS medium variant containing 0.1/0.1 mg/L NAA/BAP. Other major furanocoumarins are bergapten (highest content 245 mg/100 g DW, LS NAA/BAP 0.5/1.0) and psoralen (highest content 222 mg/100 g DW, LS NAA/BAP 0.5/1.0). Isopimpinellin and isoimperatorin were accumulated in lower amounts (76 and 54 mg/100 g DW, respectively).
In the 5-week growth cycle, the total content of linear furanocoumarins in the biomass extracts ranged between 841 and 921 mg/100 g DW. The content of individual furanocoumarins was generally lower in comparison with the 4-week growth cycle (Table 2).
The LS medium containing 0.5/1.0 mg/L NAA/BAP seemed to be the most advantageous for the production of bioactive furanocoumarins. However, there were no significant differences between LS 0.5/1.0 and 0.1/0.1 variants. In principle, both variants can be used to produce bioactive furanocoumarins. The LS medium with the highest concentration of plant growth and development regulators (LS 1.0/1.0) was the least favorable and cannot be considered for the cultivation of R. chalepensis bioreactor cultures. To increase the content of linear furanocoumarins, 4-week growth cycles are preferable (Figure 2).
3.2.2. Production of Furoquinoline Alkaloids
HPLC analyses of methanol extracts from the biomass of R. chalepensis bioreactor cultures confirmed the presence of three furoquinoline alkaloids: γ-fagarine, 7-isopentenyloxy-γ-fagarine, and skimmianine. A sample HPLC chromatogram is included in Figure S1 of the Supplementary Materials.
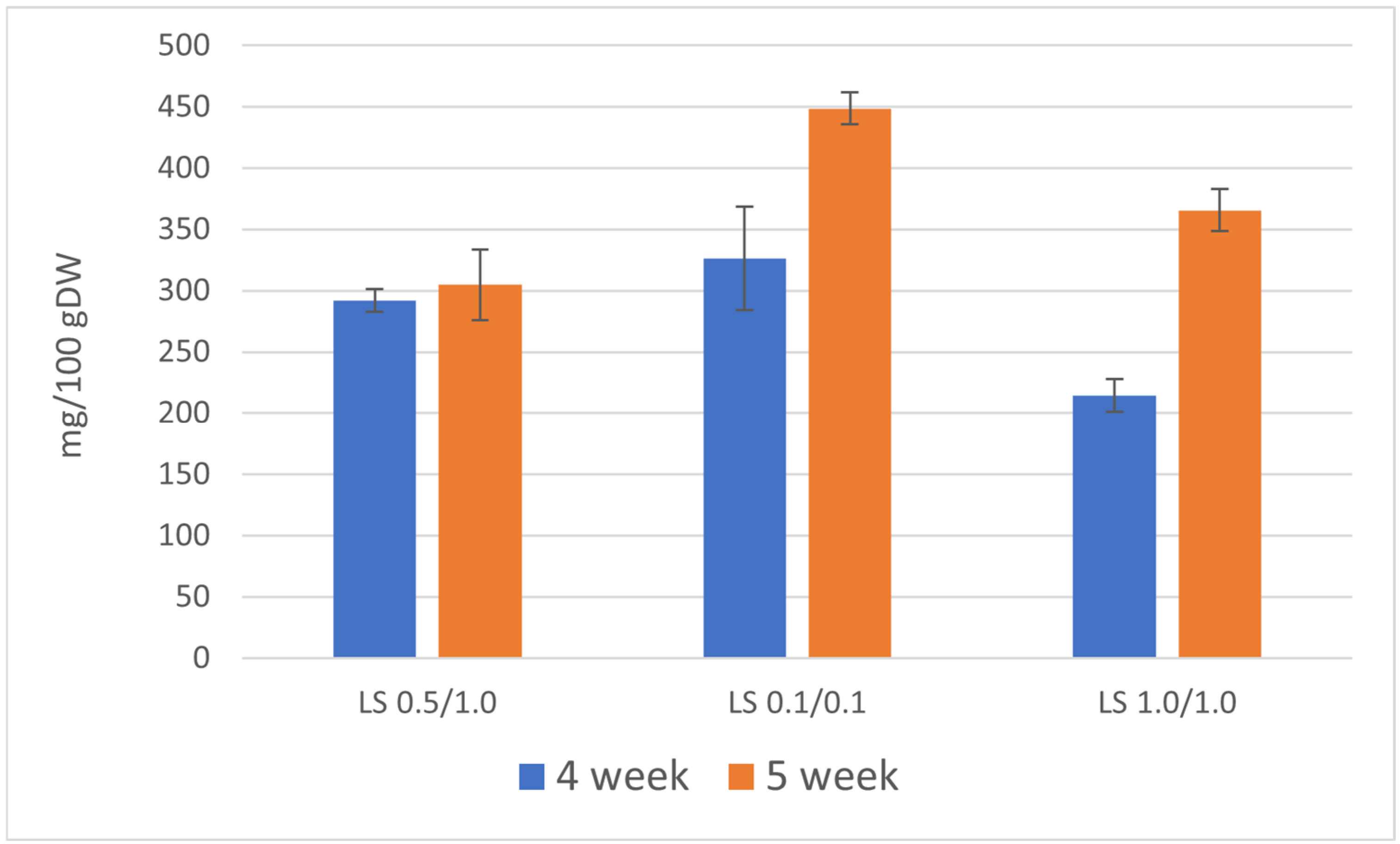
The total content of the three furoquinoline alkaloids determined in the culture extracts during the 4-week growth cycle was lower in comparison with the 5-week growth cycle and ranged between 214 and 326 mg/100 g DW. This higher accumulation of furoquinoline alkaloids was attributable to the use of a medium containing the lowest concentrations of growth and development regulators (NAA/BAP 0.1/0.1 mg/L). During the 5-week growth cycle, the total content of these three furoquinoline alkaloids ranged between 305 and 449 mg/100 g DW.
The primary compound was skimmianine, with the highest content of 292 mg/100 g DW on the LS medium variant containing 0.1/0.1 mg/L NAA/BAP (5-week growth cycle), followed by γ-fagarine (highest content 187 mg/100 g DW, LS NAA/BAP 0.1/0.1, 4-week growth cycle). 7-isopentenyloxy-γ-fagarine was accumulated in lower amounts (highest content 17 mg/100 g DW, LS NAA/BAP 0.1/0.1, 4-week growth cycle) (Table 3).
Higher content of furoquinoline alkaloids can be achieved by using the medium containing 0.1/0.1 mg/L NAA/BAP. LS media containing a higher concentration of plant growth and development regulators (LS 0.5/1.0 and 1.0/1.0) were less favorable for the production of furoquinoline alkaloids. To achieve a higher total content of furoquinoline alkaloids, 5-week growth cycles are preferable (Figure 3). However, analysis of the results for individual compounds showed that only skimmianine is produced in higher amounts during the 5-week growth cycle and the 4-week growth cycle is more favorable for γ-fagarine production (Table 3).
4. Discussion
So far, R. chalepensis in vitro cultures have not been studied in detail. Studies on in vitro callus agar cultures of R. chalepensis showed the presence of furanocoumarins such as bergapten and isopimpinellin. Furthermore, the presence of alkaloids such as skimmianine, γ-fagarine, rutacridone, rutacridone epoxide, gravacridone, and arborinin was reported. However, researchers have not quantified these metabolites [28].
R. chalepensis suspension cultures produced three metabolites: isorutarin, rutarensin, and 3-hydroxy-3-methylglutaric acid. Similarly, these metabolites have not been quantified [29].
Studies on the effect of LED light on the accumulation of the four coumarins—xanthotoxin, bergapten, psoralen, and umbelliferone—have shown that blue light increases the accumulation of these metabolites in R. chalepensis callus cultures. However, the content of these compounds was significantly lower than that of the present study. In addition, quantitative determinations were carried out on the fresh weight (FW) in the abovementioned studies, whereas in the present study, the chemical composition of the DW was analyzed. The highest content of xanthotoxin was 0.95 mg/100 g FW, and the bergapten content was higher than the xanthotoxin content and amounted to 48.9 mg/100 g FW [20].
The abovementioned studies also investigated other types of cultures (callus and suspension cultures). These cultures have a low degree of tissue differentiation, which are characterized by lower levels of secondary metabolite accumulation. In the present study, another type of in vitro culture was used—shoot culture. Due to the higher level of tissue differentiation, a higher biosynthetic potential can be observed in these cultures. Our previous studies on agitated shoot cultures of R. chalepensis (the LS medium variant NAA/BAP 0.1/0.1 mg/L) showed that the metabolism of shoot cultures is directed toward the production of furanocoumarins and furoquinoline alkaloids [25]. The results of the present study also showed a similar ability of cultures grown in bioreactors. In our previous studies, the content of the primary furanocoumarins—xanthotoxin and bergapten—was extremely high (509 and 281 mg/100 g DW, respectively), which is comparable to the results obtained in the present study (highest content 604 and 245 mg/100 g DW, respectively). However, the content of the primary alkaloids followed a different trend. The dominant compound was γ-fagarine, and skimmianine was accumulated in a lower amount, which is contrary to the present study. The content of these alkaloids was lower (γ-fagarine 79 mg/100 g DW and skimmianine 56 mg/100 g DW) than in the present study (187 and 292 mg/100 g DW, respectively) [25].
In vitro cultures may differ from their parent plants in the composition of secondary metabolites. Studies on in vitro cultures of three species of rue—R. graveolens, R. chalepensis, and R. corsica showed that the dominant secondary metabolites were compounds from the group of furanocoumarins and alkaloids, while phenolic compounds were accumulated in small amounts [25]. This observation also applies to the cultures in the present study. The content of some metabolites (furanocoumarin) was extremely high, higher than in the parent plant. On the other hand, other compounds (e.g., phenolics) were accumulated in much lower amounts. In addition, HPLC analyses did not show the presence of flavonoids, including rutoside—a characteristic flavonoid found in the genus Ruta. This may result from damage to the metabolic pathway or inhibition of the expression of certain genes [23].
In this study, R. chalepensis shoot cultures in the temporary immersion bioreactor RITA® were carried out for the first time. RITA® and other temporary immersion bioreactors (Plantform™) have recently been used to produce secondary metabolites. They are highly useful for cultivating shoot cultures. This method allows the large-scale production of secondary metabolites from various groups of compounds. Studies on Schizandra chinensis cultures carried out in various types of bioreactors showed the high production of lignans in cultures carried out in Plantform™ and RITA® bioreactors [30]. Centella asiatica cultures in RITA® and Plantform™ bioreactors were a valuable source of asiaticoside, phenolic acids, and flavonoids [31]. Leucojum aestivum cultures maintained in RITA® bioreactors produced a high content of Amaryllidaceae alkaloids, especially galanthamine and lycorine [32].
The Ruta genus has long been of interest due to its diverse chemical composition and multidirectional healing effect. Therefore, obtaining its bioactive metabolites is of interest [33]. In vitro cultures of the most well-known and studied species Ruta graveolens have been intensively studied for many years. Studies of the chemical composition of R. graveolens cultures resulted in the isolation of many metabolites from the following groups: coumarins (22 compounds), quinoline alkaloids (13 compounds), acridone alkaloids (17 compounds), and almost 50 essential oil components [34]. Many publications have focused on the possibilities of increasing the production of secondary metabolites in in vitro cultures of R. graveolens using various strategies known in plant biotechnology, such as changing the light conditions, elicitation, and feeding with phenylalanine. The most dominant furanocoumarins were xanthotoxin and bergapten, and the most dominant furoquinoline alkaloids were skimmianine and γ-fagarine [35,36,37,38]. Studies on stationary liquid cultures of R. graveolens conducted in various light conditions on LS medium containing 2/2 mg/L NAA/BAP (6-week growth cycle) showed the presence of five furanocoumarins: psoralen, xanthotoxin, isopimpinellin, bergapten, and imperatorin. Additionally, the content of umbelliferon was analyzed. The highest total content of coumarins (1022 mg/100 g DM) was detected in the cultures grown under white constant artificial light. Two main furanocoumarins were xanthotoxin and bergapten with the highest content of 433 and 219.5 mg/100 g DM, respectively [35]. In the subsequent studies on R. graveolens cultures, it was observed that the use of biotic and abiotic elicitors is a promising strategy to increase the production of furanocoumarins. The effect of abiotic elicitors (benzothiazole and saccharin) was studied in agitated shoot cultures of R. graveolens using B5 medium (4-week growth cycle). It was observed that the addition of benzothiazole in a concentration of 5% increased the production of furanocoumarins: xanthotoxin (288.36 mg/100 g DW, 8.5-fold increase, compared to control cultures), bergapten (153.78 mg/100 g DW, 3.7-fold increase), and isopimpinellin (78.9 mg/100 g DW, 14-fold increase); additionally, the cultures accumulated psoralen (82 mg/100 g DW), which was not found in the control samples [36]. Elicitation with another elicitor, chitin, also showed a positive effect on the production of furanocoumarins, but the obtained contents were slightly lower: xanthotoxin (212 mg/100 g DW, 6.3-fold increase, compared to control cultures), bergapten (146 mg/100 g DW, 3.5-fold increase), isopimpinellin (61 mg/100 g DW, 10.9-fold increase), and psoralen (68 mg/100 g DW, not detected in control cultures) [37]. The worst results were obtained using a bacterial (Bacillus sp.) lysate as an elicitor: xanthotoxin (153 mg/100 g DW, 5-fold increase, compared to control cultures), bergapten (90 mg/100 g DW, 2-fold increase), isopimpinellin (49 mg/100 g DW, 9-fold increase), and psoralen (52 mg/100 g DW, not detected in control cultures) [38].
Elicitation also had the effect of increasing the production of furoquinoline alkaloids, but the type of elicitor used had the opposite effect on alkaloid production compared to the accumulation of furanocoumarins. The addition of benzothiazole increased the production of three alkaloids: γ-fagarine (5.8 mg/100 g DW, 12-fold increase, compared to control cultures), kokusaginine (2.8 mg/100 g DW, 5.3-fold increase), and skimmianine (6.4 mg/100 g DW, 15.7-fold increase) [36]. Chitin, in this case, was more effective and increased production of γ-fagarine (12.6 mg/100 g DW, 36-fold increase), kokusaginine (4.4 mg/100 g DW, 9-fold increase), and skimmianine (14.7 mg/100 g DW, 25-fold increase). In addition, the cultures produced dictamnine (1.04 mg/100 g DW), which was not found in the control samples [37]. The best results were obtained using a bacterial lysate from Pectobacterium atrosepticum. The content of furoquinoline alkaloids was as follows: γ-fagarine (68.0 mg/100 g DW), kokusaginine (17.2 mg/100 g DW), skimmianine (48.0 mg/100 g DW), and dictamnine (9.9 mg/100 g DW) [38].
Subsequent research focused on another strategy for increasing the production of secondary metabolites in in vitro cultures of R. graveolens (agitated shoot cultures)—precursor feeding of the metabolic pathway. As a precursor, phenylalanine was used at a concentration of 1.25 g/L. The method used allowed for an increase in the production of phenolic acids and catechins, but no significant effect of phenylalanine on the production of furanocoumarins and furoquinoline alkaloids was observed. The highest concentration of furanocoumarins was as follows: 482.5 mg/100 g DW (xanthotoxin), 392.6 mg/100 g DW (bergapten), and 1059.9 mg/100 g DW (total content of furanocoumarins). Furoquinoline alkaloids were accumulated in the following amounts: 101.4 mg/100 g DW (skimmianine), 94.5 mg/100 g DW (γ-fagarine), and 185.6 mg/100 g DW (total content of furoquinoline alkaloids) [39].
In the study on R. graveolens-agitated cultures, the content of most dominant coumarins was as follows: xanthotoxin (428.3 mg/100 g DW) and bergapten (186.6 mg/100 g DW). The maximum total content (917.2 mg/100 g DW) of linear furanocoumarins (xanthotoxin, bergapten, isoimperatorin, isopimpinellin, and psoralen) was reached after a 5-week growth cycle on LS medium containing 0.1/0.1 mg/L NAA/BAP. The maximum total content of furoquinolic alkaloids was 156 mg/100 g DW. The main compounds were skimmianine (94.6 mg/100 g DW) and γ-fagarine (54.5 mg/100 g DW) [25].
The results of the current research have shown that in vitro cultures of R. chalepensis are characterized by a similar chemical composition to cultures of R. graveolens. However, the highest concentrations of main compounds obtained are higher than those in R. graveolens cultures. This may be due to the technique of cultivating cultures in RITA® bioreactors. Based on the obtained results, R. chalepensis bioreactor cultures can be a good raw material for the production of linear furanocoumarins. Xanthotoxin and bergapten are used in dermatology, and bergapten shows fewer secondary effects than xanthotoxin [19]. These studies have confirmed an extremely high content of xanthotoxin and bergapten in R. chalepensis cultures (above 600 and 245 mg/100 g DW, respectively). For instance, the content of these compounds in the parent plants can reach the highest values of 10 mg/100 g DW (xanthotoxin) and 260 mg/100 g DW (bergapten) [19].
R. chalepensis bioreactor cultures are also rich in the furoquinoline alkaloids γ-fagarine and skimmianine (highest content about 200 and 300 mg/100 g DW, respectively), which is interesting from a practical point of view.
For the production of bioactive furanocoumarins, the LS medium variant containing 0.5/1.0 mg/L NAA/BAP seemed to be the most advantageous. A higher content of alkaloids can be achieved using the variant containing 0.1/0.1 mg/L NAA/BAP. The majority of the metabolites are accumulated in higher amounts in the 4-week growth cycle, except for skimmianine, which was accumulated in higher amounts after the 5-week growth cycle.
5. Conclusions
The results of this study suggest that R. chalepensis bioreactor cultures can be a source of linear furanocoumarins and furoquinoline alkaloids. The bioreactor cultures showed good biomass growth, and the most favorable increase in biomass was observed during the 4-week growth cycle. The optimal medium to obtain biomass was found to be the LS medium containing 0.1/0.1 mg/L NAA/BAP. Variants containing lower concentrations of plant growth and development regulators NAA/BAP (0.5/1.0 and 0.1/0.1 mg/L) promoted a higher accumulation of secondary metabolites. These results encourage further research into the possibilities for the large-scale production of bioactive compounds in R. chalepensis cultures, e.g., in Plantform™ bioreactors. Strategies to increase the content of bioactive metabolites in in vitro cultures, such as elicitation and feeding with metabolic pathway precursors, can also be used. An interesting step in research could also be to investigate the effect of other plant growth and development regulators on the accumulation of secondary metabolites in bioreactor cultures of R. chalepensis.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/life13020457/s1, Figure S1: Sample chromatogram of the extract from Ruta chalepensis in vitro cultures (LS NAA/BAP 0.1/0.1 mg/L medium, 5-week growth cycle): 1. psoralen, 2. xanthotoxin, 3. isopimpinellin, 4. skimmianine, 5. bergapten, 6. γ-fagarine, 7 isoimperatorin, 8. 7-isopentenyloxy-γ-fagarine.
Author Contributions
Conceptualization, A.S.; investigation, A.S., M.G., and D.Z.; data curation, A.S. and M.G.; writing—original draft preparation, A.S.; writing—review and editing, A.S. All authors have read and agreed to the published version of the manuscript.
Funding
The research was realized as a part of the research project N42/DBS/000272 supported by the Polish Ministry of Science and Higher Education. The publication was created using the equipment cofinanced by the qLIFE Priority Research Area under the program “Excellence Initiative—Research University” at Jagiellonian University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gunaydin, K.; Savci, S. Phytochemical studies on Ruta chalepensis (Lam.) Lamarck. Nat. Prod. Res. 2005, 19, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Engler, A. Syllabus der Pflanzenfamilien, 12th ed.; Gebruder Borntraeger Verlag: Berlin, Germany, 1964. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A. Flora Europea; University Press Cambridge: Cambridge, UK, 1968; Volume 2. [Google Scholar]
- Kacem, M.; Kacem, I.; Simon, G.; Mansourd, A.B.; Chaabouni, S.; Elfeki, A.; Bouaziz, M. Phytochemicals and biological activities of Ruta chalepensis L. growing in Tunisia. Food Biosci. 2015, 12, 73–83. [Google Scholar] [CrossRef]
- Ouerghemmi, I.; Bettaieb, R.I.; Rahali, F.Z.; Bourgou, S.; Pistelli, L.; Ksouri, R.; Marzouk, B.; Tounsi, M.S. Antioxidant and antimicrobial phenolic compounds from extracts of cultivated and wild-grown Tunisian Ruta chalepensis. J. Food Drug Anal. 2017, 25, 19. [Google Scholar] [CrossRef] [PubMed]
- Tounsi, M.S.; Wannes, W.A.; Ouerghemmi, I.; Msaada, K.; Smaoui, A.; Marzouk, B. Variation in essential oil and fatty acid composition in different organs of cultivated and growing wild Ruta chalepensis L. Ind. Crops Prod. 2011, 33, 617–623. [Google Scholar] [CrossRef]
- Mejri, J.; Abderrabba, M.; Mejri, M. Chemical composition of the essential oil of Ruta chalepensis L: Influence of drying, hydro-distillation duration and plant parts. Ind. Crops Prod. 2010, 32, 671–673. [Google Scholar] [CrossRef]
- Pollio, A.; De Natale, A.; Appetiti, E.; Aliotta, G.; Touwaide, A. Continuity and change in the Mediterranean medical tradition: Ruta spp. (Rutaceae) in Hippocratic medicine and present practices. J. Ethnopharmacol. 2008, 116, 469–488. [Google Scholar] [CrossRef]
- Fakhfakh, N.; Zouari, S.; Zouari, M.; Loussayef, C.; Zouari, N. Chemical composition of volatile compounds and antioxidant activities of essential oil, aqueous and ethanol extracts of wild Tunisian Ruta chalepensis L. (Rutacea). J. Med. Plants Res. 2012, 6, 593–600. [Google Scholar]
- Gali, L.; Bedjou, F. Antioxidant and anticholinesterase effects of the ethanol extract, ethanol extract fractions and total alkaloids from the cultivated Ruta chalepensis. S. Afr. J. Bot. 2019, 120, 163–169. [Google Scholar] [CrossRef]
- Khadhri, A.; Bouali, I.; Belkhir, S.; Mokded, R.; Smiti, S.; Falé, P.; Araújo, M.E.M.; Serralheiro, M.L.M. In vitro digestion, antioxidant and antiacetylcholinesterase activities of two species of Ruta: Ruta chalepensis and Ruta montana. Pharm. Biol. 2017, 55, 101–107. [Google Scholar] [CrossRef]
- Althaher, A.R.; Oran, S.A.; Bustanji, Y.K. Phytochemical Analysis, In vitro Assessment of Antioxidant Properties and Cytotoxic Potential of Ruta chalepensis L. Essential Oil. J. Essent. Oil Bear. Plants 2020, 23, 1409–1421. [Google Scholar] [CrossRef]
- Alotaibi, S.M.; Saleem, M.S.; Al-humaidi, J.G. Phytochemical contents and biological evaluation of Ruta chalepensis L. growing in Saudi Arabia. Saudi Pharm. J. 2018, 26, 504–508. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Falco, T.; Bonesi, M.; Sicari, V.; Tundis, R.; Bruno, M. Ruta chalepensis L. (Rutaceae) leaf extract: Chemical composition, antioxidant and hypoglycemic activities. Nat. Prod. Res. 2018, 32, 521–528. [Google Scholar] [CrossRef]
- Roelandts, R. Photo(chemo) therapy for vitiligo. Photodermatol Photoimmunol Photomed 2003, 19, 1–4. [Google Scholar] [CrossRef]
- Wolf, P. Psoralen-ultraviolet A endures as one of the most powerful treatments in dermatology: Reinforcement of this ‘triple-product therapy’ by the 2016 British guidelines. Br. J. Dermatol. 2016, 174, 11–14. [Google Scholar] [CrossRef]
- Wollenberg, A.; Kinberger, M.; Arents, B.; Aszodi, N.; Avila Valle, G.; Barbarot, S.; Bieber, T.; Brough, H.A.; Calzavara Pinton, P.; Christen-Zäch, S.; et al. European guideline (EuroGuiDerm) on atopic eczema–Part II: Non-systemic treatments and treatment recommendations for special AE patient populations. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 1904–1926. [Google Scholar] [CrossRef]
- Damiani, G.; Pacifico, A.; Chu, S.; Chi, C.; Young Dermatologists Italian Network (YDIN). Frequency of phototherapy for treating psoriasis: A systematic review. Ital. J. Dermatol. Venereol. 2021, 157, 215–219. [Google Scholar] [CrossRef]
- Milesi, S.; Massot, B.; Gontier, E.; Bourgaud, F.; Guckert, A. Ruta graveolens L.: A promising species for the production of furanocoumarins. Plant Sci. 2001, 161, 189–199. [Google Scholar] [CrossRef]
- Juneja, K.; Beuerle, T.; Sircar, D. Enhanced Accumulation of Biologically Active Coumarin and Furanocoumarins in Callus Culture and Field-grown Plants of Ruta chalepensis Through LED Light-treatment. Photochem. Photobiol. 2022, 98, 1100–1109. [Google Scholar] [CrossRef]
- Murugan, N.; Srinivasan, R.; Murugan, A.; Kim, M.; Natarajan, D. Glycosmis pentaphylla (Rutaceae): A Natural Candidate for the Isolation of Potential Bioactive Arborine and Skimmianine Compounds for Controlling Multidrug-Resistant Staphylococcus aureus. Front. Public Health 2020, 8, 176. [Google Scholar] [CrossRef]
- Adamska-Szewczyk, A.; Głowniak, K.; Baj, T. Furochinoline alkaloids in plants from Rutaceae family–A review. Curr. Issues Pharm. Med. Sci. 2016, 29, 33–38. [Google Scholar] [CrossRef]
- Karuppusamy, S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J. Med. Plants Res. 2009, 3, 1222–1239. [Google Scholar]
- Shah, H.; Mtewa, A.G.; Egbuna, C.; Godwin, A.; Sesaazi, D.C. Biotechnology Approach to the Production of Phytochemicals: An Introduction. In Phytochemistry, Marine Sources, Industrial Applications, and Recent Advances; Egbuna, C., Ifemeje, J.C., Kumar, S., Sharif, N., Eds.; Apple Academic Press: Oakville, ON, Canada, 2019; Volume 3, pp. 107–130. [Google Scholar]
- Szewczyk, A.; Marino, A.; Molinari, J.; Ekiert, H.; Miceli, N. Phytochemical Characterization, and Antioxidant and Antimicrobial Properties of Agitated Cultures of Three Rue Species: Ruta chalepensis, Ruta corsica, and Ruta graveolens. Antioxidants 2022, 11, 592. [Google Scholar] [CrossRef] [PubMed]
- Linsmaier, E.; Skoog, F. Organic growth factor requirements of tobacco tissue cultures. Physiol. Plant 1965, 18, 100–127. [Google Scholar] [CrossRef]
- Sułkowska-Ziaja, K.; Maślanka, A.; Szewczyk, A.; Muszyńska, B. Physiologically active compounds in four species of Phellinus. Nat. Prod. Commun. 2017, 12, 363–366. [Google Scholar] [CrossRef]
- Baumert, A.; Gröger, D.; Kuzovkina, I.N.; Reisch, J. Secondary metabolites produced by callus cultures of various Ruta species. Plant Cell Tissue Organ Cult. 1992, 28, 159–162. [Google Scholar] [CrossRef]
- Fischer, H.; Römer, A.; Ulbrich, B.; Arens, H. A new biscoumarin glucoside ester from Ruta chalepensis cell cultures. Planta Med. 1988, 54, 398–400. [Google Scholar] [CrossRef]
- Szopa, A.; Kokotkiewicz, A.; Luczkiewicz, M.; Ekiert, H. Schisandra lignans production regulated by different bioreactor type. J. Biotechnol. 2017, 247, 11–17. [Google Scholar] [CrossRef]
- Skrzypczak-Pietraszek, E.; Piska, K.; Pietraszek, J. Enhanced production of the pharmaceutically important polyphenolic compounds in Vitex agnus castus L. shoot cultures by precursor feeding strategy. Eng. Life Sci. 2018, 18, 287–297. [Google Scholar] [CrossRef]
- Ptak, A.; Morańska, E.; Skrzypek, E.; Warchoł, M.; Spina, R.; Laurain-Mattar, D.; Simlat, M. Carbohydrates stimulated Amaryllidaceae alkaloids biosynthesis in Leucojum aestivum L. plants cultured in RITA® bioreactor. PeerJ 2020, 8, e8688. [Google Scholar] [CrossRef]
- Coimbra, A.T.; Ferreira, S.; Duarte, A.P. Genus Ruta: A natural source of high value products with biological and pharmacological properties. J. Ethnopharmacol. 2020, 260, 113076. [Google Scholar] [CrossRef]
- Petit-Paly, G.; Ramawat, K.G.; Chenieux, J.C.; Rideau, M. Ruta graveolens: In Vitro Production of Alkaloids and Medicinal Compounds. In: Bajaj, Y.P.S. (eds) Medicinal and Aromatic Plants II. Biotechnol. Agric. For. 1989, 7, 488–505. [Google Scholar] [CrossRef]
- Ekiert, H.; Gomółka, E. Effect of light on contents of coumarin compounds in shoots of Ruta graveolens L. cultivated in vitro. Acta Soc. Bot. Pol. 1999, 68, 197–200. [Google Scholar] [CrossRef]
- Orlita, A.; Sidwa-Gorycka, M.; Kumirska, J. Identification of Ruta graveolens L. metabolites accumulated in the presence of abiotic elicitors. Biotechnol. Prog. 2008, 24, 128–133. [Google Scholar] [CrossRef]
- Orlita, A.; Sidwa-Gorycka, M.; Paszkiewicz, M.; Maliński, E.; Kumirska, J.; Siedlecka, E.M.; Łojkowska, E.; Stepnowski, P. Applicationof chitin and chitosan as elicitors of coumarins and furoquinolone alkaloids in Ruta graveolens L. (common rue). Biotechnol. Appl. Biochem. 2008, 51, 91–96. [Google Scholar] [CrossRef]
- Orlita, A.; Sidwa-Gorycka, M.; Malinski, E. Effective biotic elicitation of Ruta graveolens L. shoot cultures by lysates from Pectobacterium atrosepticum and Bacillus sp. Biotechnol. Lett. 2007, 30, 541–545. [Google Scholar] [CrossRef]
- Szewczyk, A.; Paździora, W.; Ekiert, H. The Influence of Exogenous Phenylalanine on the Accumulation of Secondary Metabolites in Agitated Shoot Cultures of Ruta graveolens L. Molecules 2023, 28, 727. [Google Scholar] [CrossRef]
Figure 1.
Ruta chalepensis bioreactor culture (LS NAA/BAP 0.1/0.1 mg/L, 4-week growth cycle).

Figure 2.
Total content of furanocoumarins [mg/100 g DW] in the biomass of R. chalepensis maintained in RITA® bioreactors, depending on the duration of the growth cycle of the culture (4 and 5 weeks) and LS medium variant NAA/BAP (LS 0.5/1.0, 0.1/0.1, and 1.0/1.0).
Figure 2.
Total content of furanocoumarins [mg/100 g DW] in the biomass of R. chalepensis maintained in RITA® bioreactors, depending on the duration of the growth cycle of the culture (4 and 5 weeks) and LS medium variant NAA/BAP (LS 0.5/1.0, 0.1/0.1, and 1.0/1.0).

Figure 3.
Total content of furoquinoline alkaloids [mg/100 g DW] in the biomass of R. chalepensis maintained in RITA® bioreactors, depending on the duration of the growth cycle of culture (4 and 5 weeks) and LS medium variant NAA/BAP (LS 0.5/1.0, 0.1/0.1, 1.0/1.0).
Figure 3.
Total content of furoquinoline alkaloids [mg/100 g DW] in the biomass of R. chalepensis maintained in RITA® bioreactors, depending on the duration of the growth cycle of culture (4 and 5 weeks) and LS medium variant NAA/BAP (LS 0.5/1.0, 0.1/0.1, 1.0/1.0).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Dry weight [g] obtained from R. chalepensis cultures maintained in RITA® bioreactors, depending on the duration of the growth cycle of culture (4 and 5 weeks) and LS medium variant NAA/BAP (0.5/1.0, 0.1/0.1, and 1.0/1.0 mg/L). Mean of three replications ± SD.
Table 1.
Dry weight [g] obtained from R. chalepensis cultures maintained in RITA® bioreactors, depending on the duration of the growth cycle of culture (4 and 5 weeks) and LS medium variant NAA/BAP (0.5/1.0, 0.1/0.1, and 1.0/1.0 mg/L). Mean of three replications ± SD.
| LS Medium Variant | Dry Weight [g] | |
|---|---|---|
| NAA/BAP mg/L | 4-Week Growth Cycle | 5-Week Growth Cycle |
| 0.5/1.0 | 3.56 ± 0.26 | 3.66 ± 0.12 |
| 0.1/0.1 | 3.97 ± 0.46 | 3.93 ± 0.12 |
| 1.0/1.0 | 3.17 ± 0.14 | 3.46 ± 0.16 |
Table 2.
Contents of furanocoumarins [mg/100 g DW] obtained from R. chalepensis cultures maintained in RITA® bioreactors, depending on the duration of the growth cycle of culture (4 and 5 weeks) and LS medium variant NAA/BAP (0.5/1.0, 0.1/0.1, 1.0/1.0 mg/L). Means of three repetitions ± SD. Letters denote homogeneous groups. Different letters indicate significant differences (p < 0.05).
Table 2.
Contents of furanocoumarins [mg/100 g DW] obtained from R. chalepensis cultures maintained in RITA® bioreactors, depending on the duration of the growth cycle of culture (4 and 5 weeks) and LS medium variant NAA/BAP (0.5/1.0, 0.1/0.1, 1.0/1.0 mg/L). Means of three repetitions ± SD. Letters denote homogeneous groups. Different letters indicate significant differences (p < 0.05).
| Metabolite | LS Medium Variant | Content [mg/100 g DW] | |
|---|---|---|---|
| NAA/BAP mg/L | 4-Week Growth Cycle | 5-Week Growth Cycle | |
| Xanthotoxin | 0.5/1.0 | 592.57 ± 68.05 a | 425.23 ± 45.48 bcd |
| 0.1/0.1 | 603.96 ± 22.33 a | 486.72 ± 34.18 cd | |
| 1.0/1.0 | 375.43 ± 25.46 bc | 444.87 ± 28.55 bcd | |
| Bergapten | 0.5/1.0 | 244.76 ± 25.73 a | 136.17 ± 12.94 c |
| 0.1/0.1 | 196.89 ± 6.08 b | 129.81 ± 6.67 c | |
| 1.0/1.0 | 135.81 ± 7.09 c | 147.85 ± 3.28 c | |
| Isopimpinellin | 0.5/1.0 | 54.97 ± 4.81 a | 29.09 ± 4.23 c |
| 0.1/0.1 | 55.66 ± 11.44 a | 36.37 ± 2.66 c | |
| 1.0/1.0 | 75.58 ± 7.58 b | 84.48 ± 13.22 b | |
| Psoralen | 0.5/1.0 | 222.29 ± 32.73 a | 208.31 ± 52.47 a |
| 0.1/0.1 | 217.83 ± 12.56 a | 218.54 ± 9.97 a | |
| 1.0/1.0 | 125.88 ± 12.14 b | 119.58 ± 5.14 b | |
| Isoimperatorin | 0.5/1.0 | 54.46 ± 7.97 ab | 42.09 ± 6.32 bc |
| 0.1/0.1 | 50.28 ± 6.69 abc | 49.61 ± 7.78 abc | |
| 1.0/1.0 | 43.59 ± 2.41 abc | 50.56 ± 7.44 abc | |
| Total furanocoumarins | 0.5/1.0 | 1169.05 ± 127.51 a | 840.89 ± 90.67 bcd |
| 0.1/0.1 | 1124.62 ± 49.85 a | 921.05 ± 57.29 cd | |
| 1.0/1.0 | 756.29 ± 33.94 bc | 847.34 ± 46.07 bcd | |
Table 3.
Contents of furoquinoline alkaloids [mg/100 g DW] obtained from R. chalepensis cultures maintained in RITA® bioreactors, depending on the duration of the growth cycle of culture (4 and 5 weeks) and LS medium variant NAA/BAP (0.5/1.0, 0.1/0.1, and 1.0/1.0 mg/L). Means of three replications ± SD. Letters denote homogeneous groups. Different letters indicate significant differences (p < 0.05).
Table 3.
Contents of furoquinoline alkaloids [mg/100 g DW] obtained from R. chalepensis cultures maintained in RITA® bioreactors, depending on the duration of the growth cycle of culture (4 and 5 weeks) and LS medium variant NAA/BAP (0.5/1.0, 0.1/0.1, and 1.0/1.0 mg/L). Means of three replications ± SD. Letters denote homogeneous groups. Different letters indicate significant differences (p < 0.05).
| Metabolite | LS Medium Variant | Content [mg/100 g DW] | |
|---|---|---|---|
| NAA/BAP mg/L | 4-Week Growth Cycle | 5-Week Growth Cycle | |
| Skimmianine | 0.5/1.0 | 143.18 ± 4.89 a | 192.64 ± 19.26 d |
| 0.1/0.1 | 122.44 ± 16.27 b | 291.59 ± 3.95 e | |
| 1.0/1.0 | 74.30 ± 4.99 c | 213.53 ± 3.41 f | |
| γ-fagarine | 0.5/1.0 | 135.44 ± 9.47 ac | 98.97 ± 11.48 cd |
| 0.1/0.1 | 186.88 ± 34.50 b | 143.84 ± 10.90 ac | |
| 1.0/1.0 | 124.47 ± 14.09 acd | 136.73 ± 15.36 ac | |
| 7-isopentenyloxy-γ-fagarine | 0.5/1.0 | 13.25 ± 2.94 ac | 13.04 ± 0.37 ac |
| 0.1/0.1 | 16.79 ± 1.11 bc | 13.23 ± 2.47 ac | |
| 1.0/1.0 | 15.45 ± 0.49 abc | 15.45 ± 1.00 abc | |
| Total furoquinoline alkaloids | 0.5/1.0 | 291.88 ± 9.01 ab | 304.64 ± 28.91 ab |
| 0.1/0.1 | 326.10 ± 42.07 abe | 448.66 ± 12.86 d | |
| 1.0/1.0 | 214.22 ± 13.52 c | 365.70 ± 17.41 be | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Szewczyk, A.; Grabowski, M.; Zych, D. Ruta chalepensis L. In Vitro Cultures as a Source of Bioactive Furanocoumarins and Furoquinoline Alkaloids. Life 2023, 13, 457. https://doi.org/10.3390/life13020457
AMA Style
Szewczyk A, Grabowski M, Zych D. Ruta chalepensis L. In Vitro Cultures as a Source of Bioactive Furanocoumarins and Furoquinoline Alkaloids. Life. 2023; 13(2):457. https://doi.org/10.3390/life13020457
Chicago/Turabian StyleSzewczyk, Agnieszka, Mariusz Grabowski, and Dominika Zych. 2023. "Ruta chalepensis L. In Vitro Cultures as a Source of Bioactive Furanocoumarins and Furoquinoline Alkaloids" Life 13, no. 2: 457. https://doi.org/10.3390/life13020457
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.