

The Topological Characteristics of Biological Ratio-Sensing Networks


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
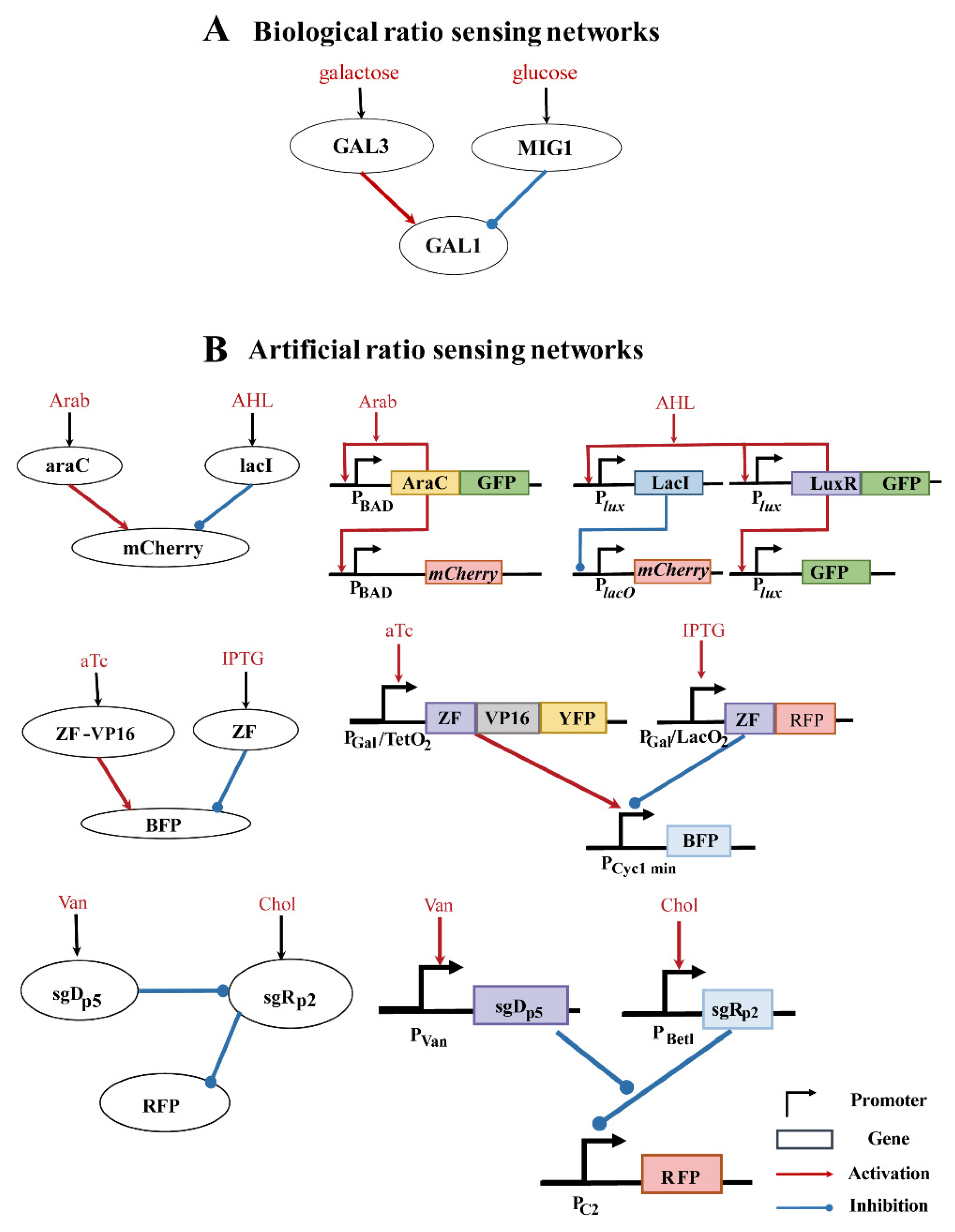
:1. Introduction
2. Materials and Methods
2.1. The Coarse-Grained Computational Enumeration of 3-Node Networks
2.2. Modeling Network Behavior with Ordinary Differential Equations
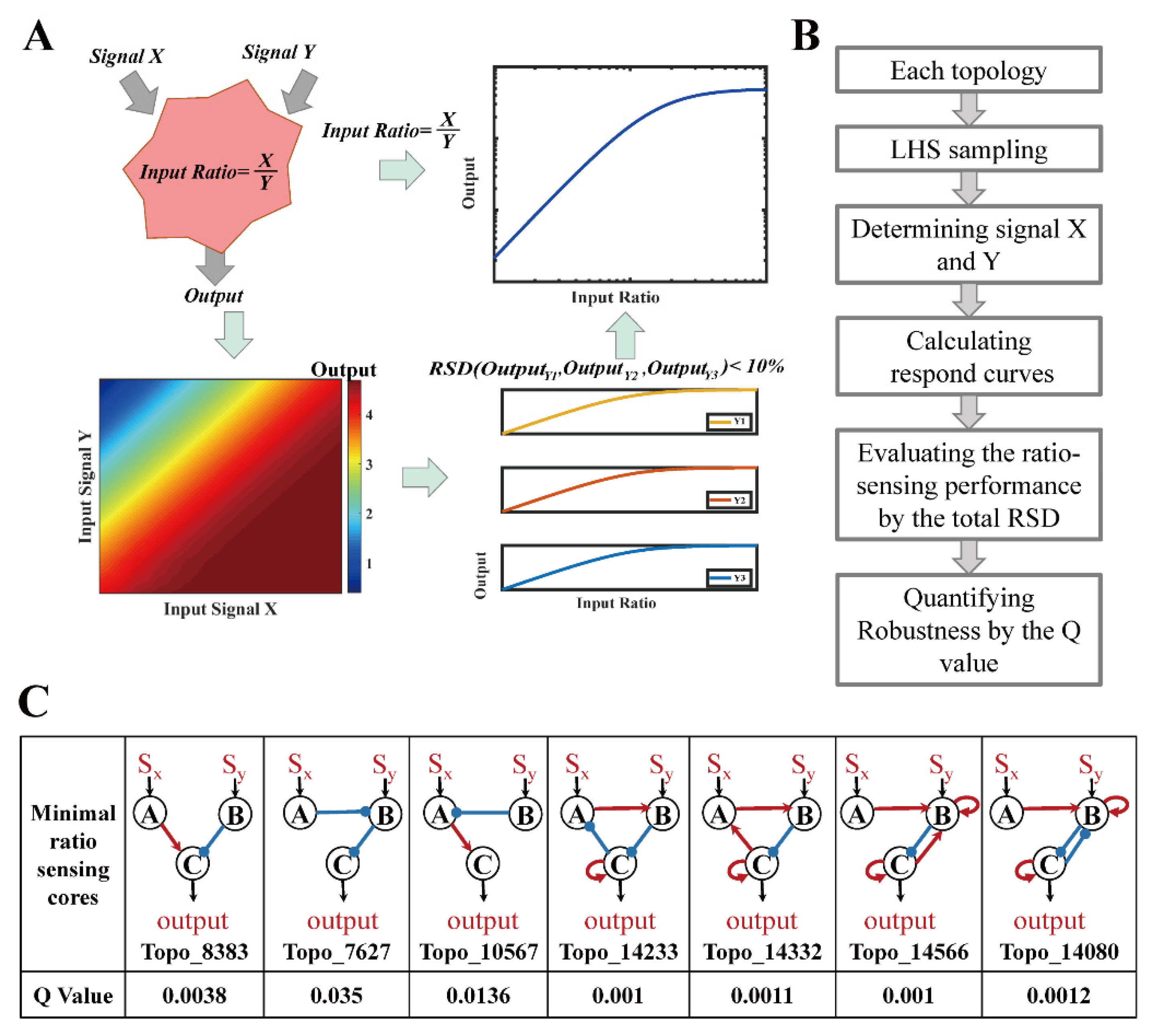
2.3. Definition and Quantification of Ratio-Sensing Performance
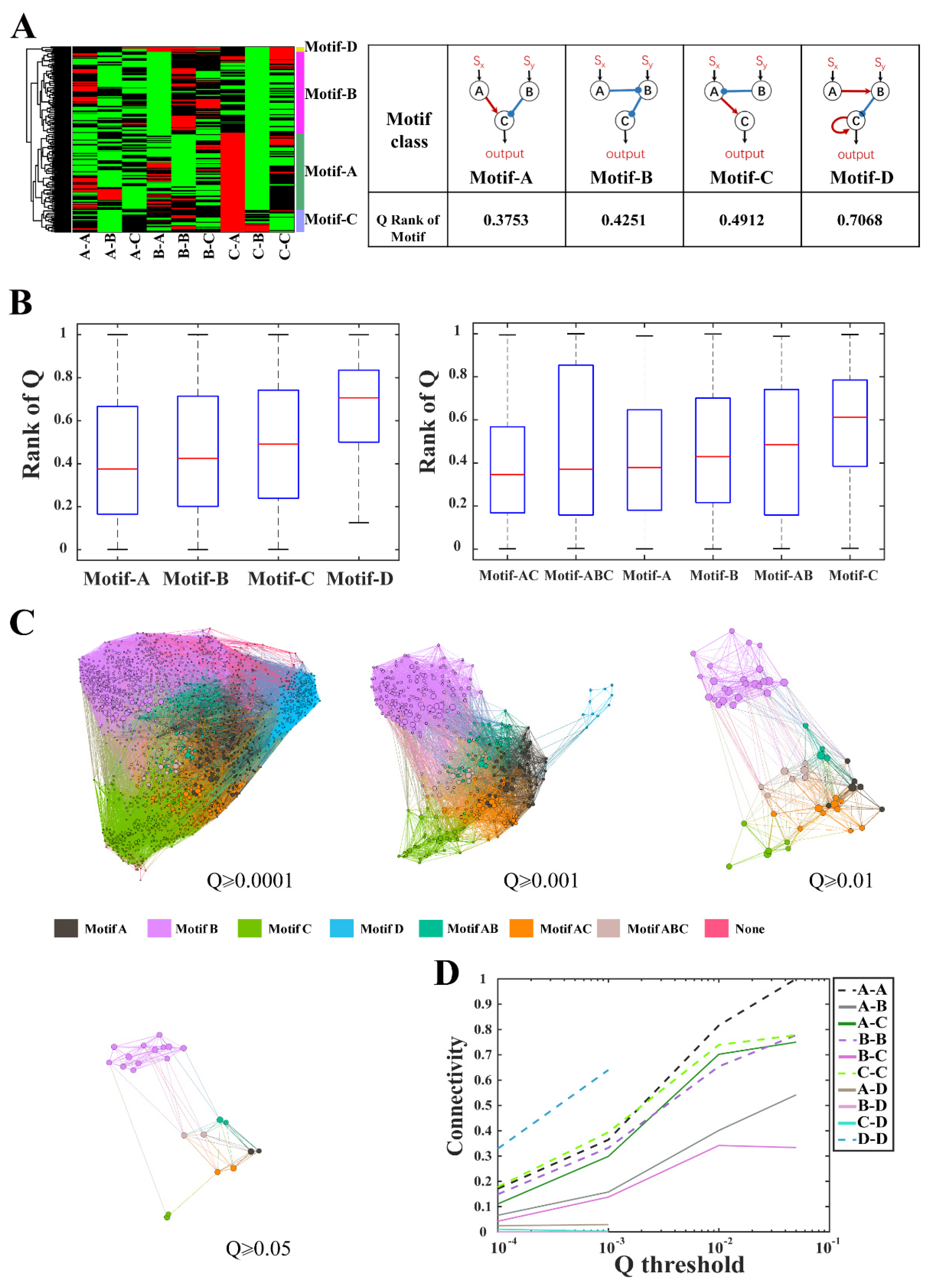
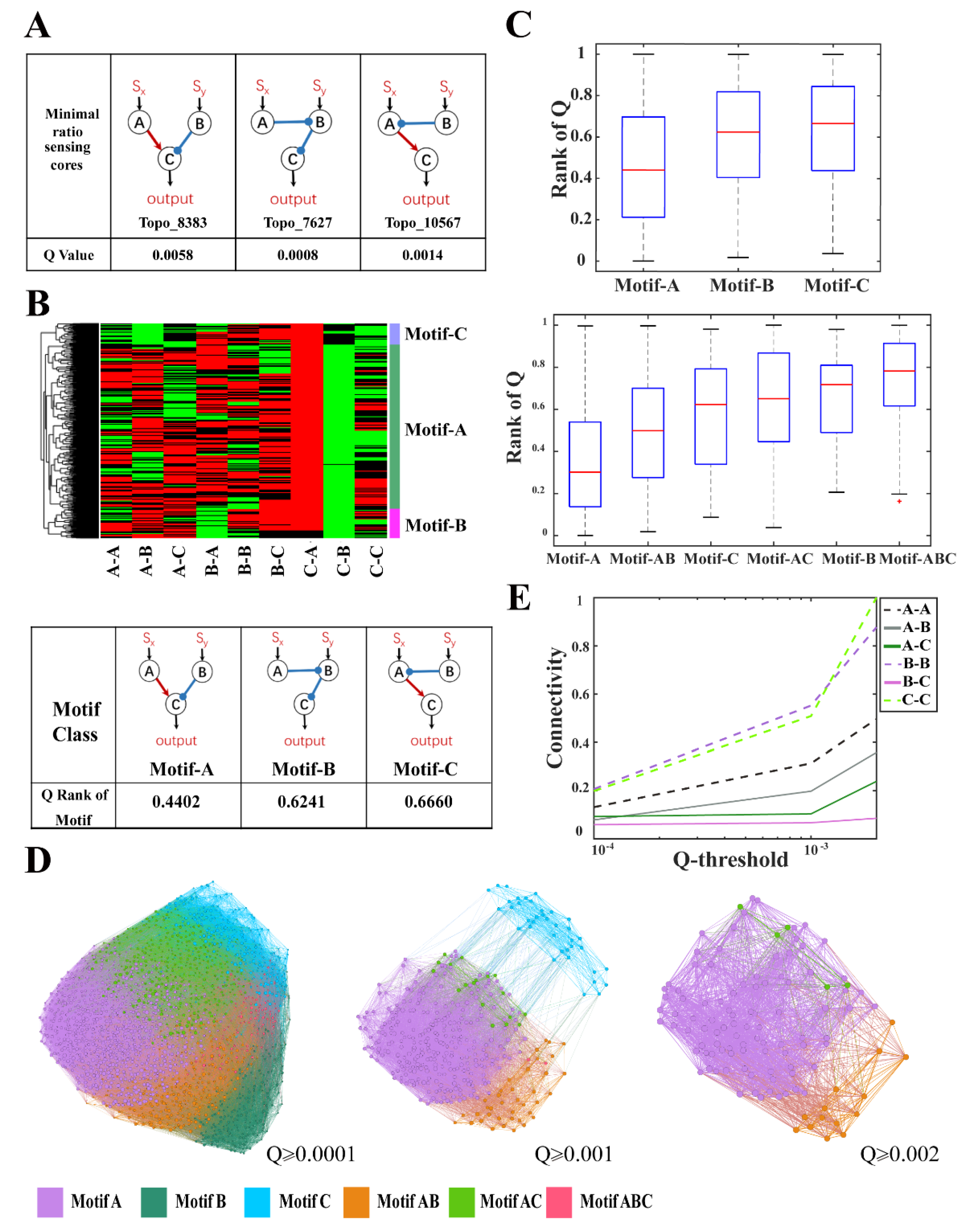
2.4. The Minimal Core Topologies and Motif Selection
2.5. Evolutionary Metagraph
3. Results
3.1. Ratio-Sensing Definition and Three-Node Network Enumeration
3.2. Structural Characteristics of Ratio-Sensing Topologies for Enzymatic Networks
3.3. Robustness and Evolutionary Connectivity
3.4. Structural Characteristics of Ratio-Sensing Topologies for Transcriptional Networks
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gancedo, J.M. Yeast carbon catabolite repression. Microbiol. Mol. Biol. Rev. 1998, 62, 334–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohr, D.; Venkov, P.; Zlatanova, J. Transcriptional regulation in the yeast GAL gene family: A complex genetic network. FASEB J. 1995, 9, 777–787. [Google Scholar] [CrossRef]
- Wang, J.; Atolia, E.; Hua, B.; Savir, Y.; Escalante-Chong, R.; Springer, M. Natural variation in preparation for nutrient depletion reveals a cost-benefit tradeoff. PLoS Biol. 2015, 13, e1002041. [Google Scholar] [CrossRef] [Green Version]
- Deutscher, J. The mechanisms of carbon catabolite repression in bacteria. Curr. Opin. Microbiol. 2008, 11, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Escalante-Chong, R.; Savir, Y.; Carroll, S.M.; Ingraham, J.B.; Wang, J.; Marx, C.J.; Springer, M. Galactose metabolic genes in yeast respond to a ratio of galactose and glucose. Proc. Natl. Acad. Sci. USA 2015, 112, 1636–1641. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Hua, B.; Springer, M.; Tang, C. Computational study on ratio-sensing in yeast galactose utilization pathway. PLoS Comput. Biol. 2020, 16, e1007960. [Google Scholar] [CrossRef]
- Hong, J.; Palme, J.; Hua, B.; Springer, M. Computational analysis of GAL pathway pinpoints mechanisms underlying natural variation. PLoS Comput. Biol. 2021, 17, e1008691. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiong, W.; Zhang, E.E. The ratio of intracellular CRY proteins determines the clock period length. Biochem. Biophys. Res. Commun. 2016, 472, 531–538. [Google Scholar] [CrossRef]
- Raisova, M.; Hossini, A.M.; Eberle, J.; Riebeling, C.; Wieder, T.; Sturm, I.; Daniel, P.T.; Orfanos, C.E.; Geilen, C.C. The Bax/Bcl-2 ratio determines the susceptibility of human melanoma cells to CD95/Fas-mediated apoptosis. J. Investig. Dermatol. 2001, 117, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Berg, J.; Hung, Y.P.; Yellen, G. A genetically encoded fluorescent reporter of ATP:ADP ratio. Nat. Methods 2009, 6, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Tantama, M.; Martínez-François, J.R.; Mongeon, R.; Yellen, G. Imaging energy status in live cells with a fluorescent biosensor of the intracellular ATP-to-ADP ratio. Nat. Commun. 2013, 4, 2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Landick, R.; Raman, S. A Regulatory NADH/NAD+ Redox Biosensor for Bacteria. ACS Synth. Biol. 2019, 8, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Antebi, Y.E.; Linton, J.M.; Klumpe, H.; Bintu, B.; Gong, M.; Su, C.; MaCardell, R.; Elowitz, M.B. Combinatorial Signal Perception in the BMP Pathway. Cell 2017, 170, 1184–1196.e24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, R.; Rubens, J.R.; Sarpeshkar, R.; Lu, T.K. Synthetic analog computation in living cells. Nature. 2013, 497, 619–623. [Google Scholar] [CrossRef]
- Perli, S.D.; Lu, T.K. Ratiometric logic in living cells via competitive binding of synthetic transcription factors. In Proceedings of the 4th ACM International Conference on Nanoscale Computing and Communicatio, Washington, DC, USA, 27–29 September 2017; pp. 1–4. [Google Scholar] [CrossRef]
- Anderson, D.A.; Voigt, C.A. Competitive dCas9 binding as a mechanism for transcriptional control. Mol. Syst. Biol. 2021, 17, e10512. [Google Scholar] [CrossRef] [PubMed]
- Alon, U. Network motifs: Theory and experimental approaches. Nat. Rev. Genet. 2007, 8, 450–461. [Google Scholar] [CrossRef]
- Lim, W.A.; Lee, C.M.; Tang, C. Design principles of regulatory networks: Searching for the molecular algorithms of the cell. Mol Cell. 2013, 49, 202–212. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Trusina, A.; El-Samad, H.; Lim, W.A.; Tang, C. Defining network topologies that can achieve biochemical adaptation. Cell 2009, 138, 760–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.; Lai, L.; Ouyang, Q.; Tang, C. Robustness and modular design of the Drosophila segment polarity network. Mol. Syst. Biol. 2006, 2, 70. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Ouyang, Q.; Wang, H. Dose-response aligned circuits in signaling systems. PLoS ONE 2012, 7, e34727. [Google Scholar] [CrossRef] [Green Version]
- Chau, A.H.; Walter, J.M.; Gerardin, J.; Tang, C.; Lim, W.A. Designing synthetic regulatory networks capable of self-organizing cell polarization. Cell. 2012, 151, 320–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, W.; Du, P.; Lou, Q.; Wu, L.; Zhang, H.M.; Lou, C.; Wang, H.; Ouyang, Q. Rational Design of an Ultrasensitive Quorum-Sensing Switch. ACS Synth. Biol. 2017, 6, 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.M.; Shao, B.; Ouyang, Q. Identifying network topologies that can generate turing pattern. J. Theor. Biol. 2016, 408, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.Y.; Ouyang, Q. Using Sub-Network Combinations to Scale Up an Enumeration Method for Determining the Network Structures of Biological Functions. PLoS ONE 2016, 11, e0168214. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Ouyang, Q.; Wang, H. Robust network topologies for generating oscillations with temperature-independent periods. PLoS ONE 2017, 12, e0171263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo-Hair, S.M.; Villota, E.R.; Coronado, A.M. Design principles for robust oscillatory behavior. Syst. Synth. Biol. 2015, 9, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Liu, S.; Yang, Q. Incoherent Inputs Enhance the Robustness of Biological Oscillators. Cell Syst. 2017, 5, 72–81.e4. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Wang, H.; Ouyang, Q. Constructing network topologies for multiple signal-encoding functions. BMC Syst. Biol. 2019, 13, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerardin, J.; Reddy, N.R.; Lim, W.A. The Design Principles of Biochemical Timers: Circuits that Discriminate between Transient and Sustained Stimulation. Cell Syst. 2019, 9, 297–308.e2. [Google Scholar] [CrossRef] [PubMed]
- Milo, R.; Shen-Orr, S.; Itzkovitz, S.; Kashtan, N.; Chklovskii, D.; Alon, U. Network motifs: Simple building blocks of complex networks. Science. 2002, 298, 824–827. [Google Scholar] [CrossRef] [Green Version]
- Alon, U. An Introduction to Systems Biology: Design Principles of Biological Circuits; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Zong, Y.; Zhang, H.M.; Lyu, C.; Ji, X.; Hou, J.; Guo, X.; Ouyang, Q.; Lou, C. Insulated transcriptional elements enable precise design of genetic circuits. Nat. Commun. 2017, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Moon, T.S.; Lou, C.; Tamsir, A.; Stanton, B.C.; Voigt, C.A. Genetic programs constructed from layered logic gates in single cells. Nature. 2012, 491, 249–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, P.; Zhao, H.; Zhang, H.; Wang, R.; Huang, J.; Tian, Y.; Luo, X.; Luo, X.; Wang, M.; Xiang, Y.; et al. De novo design of an intercellular signaling toolbox for multi-channel cell-cell communication and biological computation. Nat. Commun. 2020, 11, 4226. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Du, C.; Chen, H.; Simha, R.; Rong, Y.; Xiao, Y.; Zeng, C. Process-based network decomposition reveals backbone motif structure. Proc. Natl. Acad. Sci. USA 2010, 107, 10478–10483. [Google Scholar] [CrossRef] [Green Version]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. In Proceedings of the International AAAI Conference on Web and Social Media, San Jose, CA, USA, 17–20 May 2009; pp. 361–362. [Google Scholar]
- Jacomy, M.; Venturini, T.; Heymann, S.; Bastian, M. ForceAtlas2, a continuous graph layout algorithm for handy network visualization designed for the Gephi software. PLoS ONE 2014, 9, e98679. [Google Scholar] [CrossRef]
- Ciliberti, S.; Martin, O.C.; Wagner, A. Robustness can evolve gradually in complex regulatory gene networks with varying topology. PLoS Comput. Biol. 2007, 3, e15. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Wang, T.; Guan, Y.; Ouyang, Q.; Lou, C.; Qian, L. The Topological Characteristics of Biological Ratio-Sensing Networks. Life 2023, 13, 351. https://doi.org/10.3390/life13020351
Chen X, Wang T, Guan Y, Ouyang Q, Lou C, Qian L. The Topological Characteristics of Biological Ratio-Sensing Networks. Life. 2023; 13(2):351. https://doi.org/10.3390/life13020351
Chicago/Turabian StyleChen, Xinmao, Tianze Wang, Ying Guan, Qi Ouyang, Chunbo Lou, and Long Qian. 2023. "The Topological Characteristics of Biological Ratio-Sensing Networks" Life 13, no. 2: 351. https://doi.org/10.3390/life13020351