
Potential Health Benefits of Banana Phenolic Content during Ripening by Implementing Analytical and In Silico Techniques

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Standards and Reagents
2.2. Sample Collection
2.3. Extraction of Phenolic Compounds and Spectrophotometric Assays
2.4. LC-ESI(−)-MS/MS Analysis
2.5. Computational Studies
2.5.1. Phenolic Compounds Preparation
2.5.2. Protein Target Prediction and Molecular Docking Studies
2.6. Statistical Analysis
3. Results and Discussion
3.1. Spectrophotometric Assays in Banana Samples
3.2. Phenolic Profile of the Examined Banana Samples by LC-MS/MS Analysis
3.3. Computational Studies
Target Prediction and Molecular Docking Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phillips, K.M.; McGinty, R.C.; Couture, G.; Pehrsson, P.R.; McKillop, K.; Fukagawa, N.K. Dietary Fiber, Starch, and Sugars in Bananas at Different Stages of Ripeness in the Retail Market. PLoS ONE 2021, 16, e0253366. [Google Scholar] [CrossRef]
- da Costa Nascimento, R.; de Oliveira Freire, O.; Ribeiro, L.S.; Araújo, M.B.; Finger, F.L.; Soares, M.A.; Wilcken, C.F.; Zanuncio, J.C.; Souto Ribeiro, W. Ripening of Bananas Using Bowdichia Virgilioides Kunth Leaves. Sci. Rep. 2019, 9, 3548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marta, H.; Cahyana, Y.; Djali, M.; Pramafisi, G. The Properties, Modification, and Application of Banana Starch. Polymers 2022, 14, 3092. [Google Scholar] [CrossRef] [PubMed]
- FAO. Banana Market Review 2019; FAO: Rome, Italy, 2020. [Google Scholar]
- Zou, F.; Tan, C.; Zhang, B.; Wu, W.; Shang, N. The Valorization of Banana By-Products: Nutritional Composition, Bioactivities, Applications, and Future Development. Foods 2022, 11, 3170. [Google Scholar] [CrossRef]
- Bashmil, Y.M.; Ali, A.; Bk, A.; Dunshea, F.R.; Suleria, H.A.R. Screening and Characterization of Phenolic Compounds from Australian Grown Bananas and Their Antioxidant Capacity. Antioxidants 2021, 10, 1521. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.F.; Khalid, W.; Akram, S.; Khalid, M.A.; Zubair, M.; Kauser, S.; Mohamedahmed, K.A.; Aziz, A.; Siddiqui, S.A. Bioactive profile and functional food applications of banana in food sectors and health: A review. Int. J. Food Prop. 2022, 25, 2286–2300. [Google Scholar] [CrossRef]
- Sidhu, J.S.; Zafar, T.A. Chemical Composition and Nutritional Profile of Raw and Processed Banana Products. Handb. Banan. Prod. Postharvest Sci. Process. Technol. Nutr. First Ed. 2020, 11, 207–225. [Google Scholar] [CrossRef]
- Borges, C.V.; Maraschin, M.; Coelho, D.S.; Leonel, M.; Gomez, H.A.G.; Belin, M.A.F.; Diamante, M.S.; Amorim, E.P.; Gianeti, T.; Castro, G.R.; et al. Nutritional Value and Antioxidant Compounds during the Ripening and after Domestic Cooking of Bananas and Plantains. Food Res. Int. 2020, 132, 109061. [Google Scholar] [CrossRef]
- Arias, A.; Feijoo, G.; Moreira, M.T. Exploring the potential of antioxidants from fruits and vegetables and strategies for their recovery. Innov. Food Sci. Emerg. Technol. 2022, 77, 102974. [Google Scholar] [CrossRef]
- Dong, C.; Wang, J.; Hu, Y.; Xiao, H.; Xie, J. Analyses of key gene networks controlling carotenoid metabolism in Xiangfen 1 banana. BMC Plant Biol. 2022, 22, 34. [Google Scholar] [CrossRef]
- Rad, N.K.; Mohri, M.; Seifi, H.A.; Haghparast, A. Supplementation of overripe pulp extract and green peel extract or powder of banana fruit peel (musa. cavendish) to diets of neonatal dairy calves: Effects on haematological, immunological and performance characteristics. Med. Sci. 2021, 7, 876–887. [Google Scholar] [CrossRef]
- Passo Tsamo, C.V.; Herent, M.-F.; Tomekpe, K.; Happi Emaga, T.; Quetin-Leclercq, J.; Rogez, H.; Larondelle, Y.; Andre, C. Phenolic Profiling in the Pulp and Peel of Nine Plantain Cultivars (Musa sp.). Food Chem. 2015, 167, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Gedük, A.Ş.; Zengin, F. LC–MS/MS Characterization, Antidiabetic, Antioxidative, and Antibacterial Effects of Different Solvent Extracts of Anamur Banana (Musa Cavendishii). Food Sci. Biotechnol. 2021, 30, 1183–1193. [Google Scholar] [CrossRef]
- Kongkachuichai, R.; Charoensiri, R.; Sungpuag, P. Carotenoid, Flavonoid Profiles and Dietary Fiber Contents of Fruits Commonly Consumed in Thailand. Int. J. Food Sci. Nutr. 2010, 61, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Jaganath, I.B.; Ludwig, I.A.; Crozier, A. Chlorogenic Acids and the Acyl-Quinic Acids: Discovery, Biosynthesis, Bioavailability and Bioactivity. Nat. Prod. Rep. 2017, 34, 1391–1421. [Google Scholar] [CrossRef] [Green Version]
- Maduwanthi, S.D.T.; Marapana, R.A.U.J. Total phenolics, flavonoids and antioxidant activity following simulated gastro-intestinal digestion and dialysis of banana (Musa acuminata, AAB) as affected by induced ripening agents. Food Chem. 2021, 339, 127909. [Google Scholar] [CrossRef]
- Sidhu, J.S.; Zafar, T.A. Bioactive Compounds in Banana Fruits and Their Health Benefits. Food Qual. Saf. 2018, 2, 183–188. [Google Scholar] [CrossRef]
- Sarma, P.P.; Gurumayum, N.; Verma, A.K.; Devi, R. A Pharmacological Perspective of Banana: Implications Relating to Therapeutic Benefits and Molecular Docking. Food Funct. 2021, 12, 4749–4767. [Google Scholar] [CrossRef]
- Mondal, A.; Banerjee, S.; Bose, S.; Das, P.P.; Sandberg, E.N.; Atanasov, A.G.; Bishayee, A. Cancer Preventive and Therapeutic Potential of Banana and Its Bioactive Constituents: A Systematic, Comprehensive, and Mechanistic Review. Front. Oncol. 2021, 11, 697143. [Google Scholar] [CrossRef]
- Andreou, V.; Strati, I.F.; Fotakis, C.; Liouni, M.; Zoumpoulakis, P.; Sinanoglou, V.J. Herbal Distillates: A New Era of Grape Marc Distillates with Enriched Antioxidant Profile. Food Chem. 2018, 253, 171–178. [Google Scholar] [CrossRef]
- Lantzouraki, D.Z.; Sinanoglou, V.J.; Zoumpoulakis, P.G.; Glamočlija, J.; Ćirić, A.; Soković, M.; Heropoulos, G.; Proestos, C. Antiradical–Antimicrobial Activity and Phenolic Profile of Pomegranate (Punica Granatum L.) Juices from Different Cultivars: A Comparative Study. RSC Adv. 2014, 5, 2602–2614. [Google Scholar] [CrossRef]
- Lantzouraki, D.Z.; Sinanoglou, V.J.; Zoumpoulakis, P.; Proestos, C. Comparison of the Antioxidant and Antiradical Activity of Pomegranate (Punica Granatum L.) by Ultrasound-Assisted and Classical Extraction. Anal. Lett. 2016, 49, 969–978. [Google Scholar] [CrossRef]
- Tsiaka, T.; Kritsi, E.; Lantzouraki, D.Z.; Christodoulou, P.; Tsigrimani, D.; Strati, I.F.; Sinanoglou, V.J.; Zoumpoulakis, P. Assessing the Phytochemical Profile and Potential of Traditional Herbal Infusions against Aldose Reductase through In Silico Studies and LC-MS/MS Analysis. Appl. Sci. 2022, 12, 8361. [Google Scholar] [CrossRef]
- Kavga, A.; Strati, I.F.; Sinanoglou, V.J.; Fotakis, C.; Sotiroudis, G.; Christodoulou, P.; Zoumpoulakis, P. Evaluating the Experimental Cultivation of Peppers in Low-Energy-Demand Greenhouses. An Interdisciplinary Study. J. Sci. Food Agric. 2019, 99, 781–789. [Google Scholar] [CrossRef]
- Schrödinger Release 2020-3, LigPrep; Schrödinger, LLC: New York, NY, USA, 2020.
- Schrödinger Release 2020-3, Maestro; Schrödinger, LLC: New York, NY, USA, 2020.
- Lee, K.; Lee, M.; Kim, D. Utilizing Random Forest QSAR Models with Optimized Parameters for Target Identification and Its Application to Target-Fishing Server. BMC Bioinform. 2017, 18, 567. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated Data and New Features for Efficient Prediction of Protein Targets of Small Molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrödinger Release 2020-3, Protein Preparation Wizard; Schrödinger, LLC: New York, NY, USA, 2020.
- Schrödinger Release 2020-3, Glide; Schrödinger, LLC: New York, NY, USA, 2020.
- Fernando, H.; Srilaong, V.; Pongprasert, N.; Boonyaritthongchai, P.; Jitareerat, P. Changes in Antioxidant Properties and Chemical Composition during Ripening in Banana Variety “Hom Thong” (AAA Group) and “Khai” (AA Group). Int. Food Res. J. 2013, 21, 749–754. [Google Scholar]
- Aquino, C.F.; Salomão, L.C.C.; Ribeiro, S.M.R.; Siqueira, D.L.D.; Cecon, P.R. Carbohydrates, Phenolic Compounds and Antioxidant Activity in Pulp and Peel of 15 Banana Cultivars. Rev. Bras. Frutic. 2016, 38, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bilgin, Ö.F.; Nogay, G.; Kafkas, E.; Kafkas, S. Some Fruit Quality Characteristics of ‘Grand Naine’ Banana Fruits during Various Ripening Stages. Int. J. Agric. For. Life Sci. 2022, 6, 24–27. [Google Scholar]
- S Reihani, S.F.; Ramli, S.; Alkarkhi, A.; Easa, A. Total Phenolics, Flavonoids and Antioxidant Activity of Banana Pulp and Peel Flours: Influence of Variety and Stage of Ripeness. Int. Food Res. J. 2012, 19, 1041–1046. [Google Scholar]
- Yadav, A. Banana (Musa Acuminata): Most Popular and Common Indian Plant with Multiple Pharmacological Potentials. World J. Biol. Pharm. Health Sci. 2021, 7, 36–44. [Google Scholar] [CrossRef]
- Wongwaiwech, D.; Kamchonemenukool, S.; Ho, C.-T.; Li, S.; Thongsook, T.; Majai, N.; Premjet, D.; Sujipuli, K.; Weerawatanakorn, M. Nutraceutical Difference between Two Popular Thai Namwa Cultivars Used for Sun Dried Banana Products. Molecules 2022, 27, 5675. [Google Scholar] [CrossRef]
- Li, X.; Sun, J.; Chen, Z.; Jiang, J.; Jackson, A. Characterization of Carotenoids and Phenolics during Fruit Ripening of Chinese Raspberry (Rubus Chingii Hu). RSC Adv. 2021, 11, 10804–10813. [Google Scholar] [CrossRef] [PubMed]
- Pothavorn, P.; Kitdamrongsont, K.; Swangpol, S.; Wongniam, S.; Atawongsa, K.; Svasti, J.; Somana, J. Sap phytochemical compositions of some bananas in Thailand. J. Agric. Food Chem. 2010, 58, 8782–8787. [Google Scholar] [CrossRef]
- Zhu, C.; Wu, S.; Sun, T.; Zhou, Z.; Hu, Z.; Yu, J. Rosmarinic Acid Delays Tomato Fruit Ripening by Regulating Ripening-Associated Traits. Antioxidants 2021, 10, 1821. [Google Scholar] [CrossRef]
- Siriamornpun, S.; Kaewseejan, N. Quality, Bioactive Compounds and Antioxidant Capacity of Selected Climacteric Fruits with Relation to Their Maturity. Sci. Hortic. 2017, 221, 33–42. [Google Scholar] [CrossRef]
- Cai, J.; Chen, T.; Zhang, Z.; Li, B.; Qin, G.; Tian, S. Metabolic Dynamics During Loquat Fruit Ripening and Postharvest Technologies. Front. Plant Sci. 2019, 10, 619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, C.; Hu, H.; Hu, Y.; Xie, J. Metabolism of Flavonoids in Novel Banana Germplasm during Fruit Development. Front. Plant Sci. 2016, 7, 1291. [Google Scholar] [CrossRef] [Green Version]
- Shamla, L.; Nisha, P. Acrylamide Formation in Plantain (Musa Paradisiaca) Chips Influenced by Different Ripening Stages: A Correlation Study with Respect to Reducing Sugars, Amino Acids and Phenolic Content. Food Chem. 2017, 222, 53–60. [Google Scholar] [CrossRef]
- Vu, H.T.; Scarlett, C.J.; Vuong, Q.V. Phenolic Compounds within Banana Peel and Their Potential Uses: A Review. J. Funct. Foods 2018, 40, 238–248. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Bioactive Compounds in Banana and Their Associated Health Benefits—A Review. Food Chem. 2016, 206, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Structure and Function of Carbonic Anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Saadiq, M.; Uddin, G.; Latif, A.; Ali, M.; Akbar, N.; Ammara; Ali, S.; Ahmad, M.; Zahoor, M.; Khan, A.; et al. Synthesis, Bioactivity Assessment, and Molecular Docking of Non-Sulfonamide Benzimidazole-Derived N-Acylhydrazone Scaffolds as Carbonic Anhydrase-II Inhibitors. ACS Omega 2022, 7, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T.; Scozzafava, A. Carbonic Anhydrases as Targets for Medicinal Chemistry. Bioorganic Med. Chem. 2007, 15, 4336–4350. [Google Scholar] [CrossRef]
- Carta, F.; Dumy, P.; Supuran, C.T.; Winum, J.-Y. Multivalent Carbonic Anhydrases Inhibitors. IJMS 2019, 20, 5352. [Google Scholar] [CrossRef] [Green Version]
- Cuffaro, D.; Nuti, E.; Rossello, A. An Overview of Carbohydrate-Based Carbonic Anhydrase Inhibitors. J. Enzym. Inhib. Med. Chem. 2020, 35, 1906–1922. [Google Scholar] [CrossRef] [PubMed]
- Lomelino, C.; McKenna, R. Carbonic Anhydrase Inhibitors: A Review on the Progress of Patent Literature (2011–2016). Expert Opin. Ther. Pat. 2016, 26, 947–956. [Google Scholar] [CrossRef]
- Aggul, A.G.; Uzun, N.; Kuzu, M.; Taslimi, P.; Gulcin, I. Some Phenolic Natural Compounds as Carbonic Anhydrase Inhibitors: An in Vitro and in Silico Study. Arch. der Pharm. 2022, 355, 2100476. [Google Scholar] [CrossRef]
- Huang, H.; Pan, X.; Ji, C.; Zeng, G.; Jiang, L.; Fu, X.; Liu, J.; Hao, X.; Zhang, Y.; Tan, N. Screening and Docking Studies of Natural Phenolic Inhibitors of Carbonic Anhydrase II. Sci. China Ser. B-Chem. 2009, 52, 332–337. [Google Scholar] [CrossRef]
- Rauf, A.; Raza, M.; Saleem, M.; Ozgen, U.; Karaoglan, E.S.; Renda, G.; Palaska, E.; Orhan, I.E. Carbonic Anhydrase and Urease Inhibitory Potential of Various Plant Phenolics Using in Vitro and in Silico Methods. Chem. Biodivers. 2017, 14, e1700024. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Steps of Elution (min) | Water-0.2% v/v Formic Acid | Acetonitrile-0.1% v/v Formic Acid | Flow Rate (μL/min) |
|---|---|---|---|
| 0.0 | 90 | 10 | 300 |
| 0.5 | 80 | 20 | 300 |
| 4.0 | 70 | 30 | 300 |
| 4.1 | 50 | 50 | 350 |
| 4.5 | 50 | 50 | 350 |
| 5.1 | 35 | 65 | 350 |
| 7.0 | 0 | 100 | 350 |
| 8.0 | 0 | 100 | 350 |
| 9.0 | 0 | 100 | 300 |
| 9.1 | 90 | 10 | 300 |
| 15 | 90 | 10 | 300 |
| Correlation Coefficients | TPC | ABTS●+ | FRAP |
|---|---|---|---|
| TPC | 1 | 0.53 | 0.52 |
| ABTS●+ | 0.53 | 1 | 0.95 |
| FRAP | 0.52 | 0.95 | 1 |
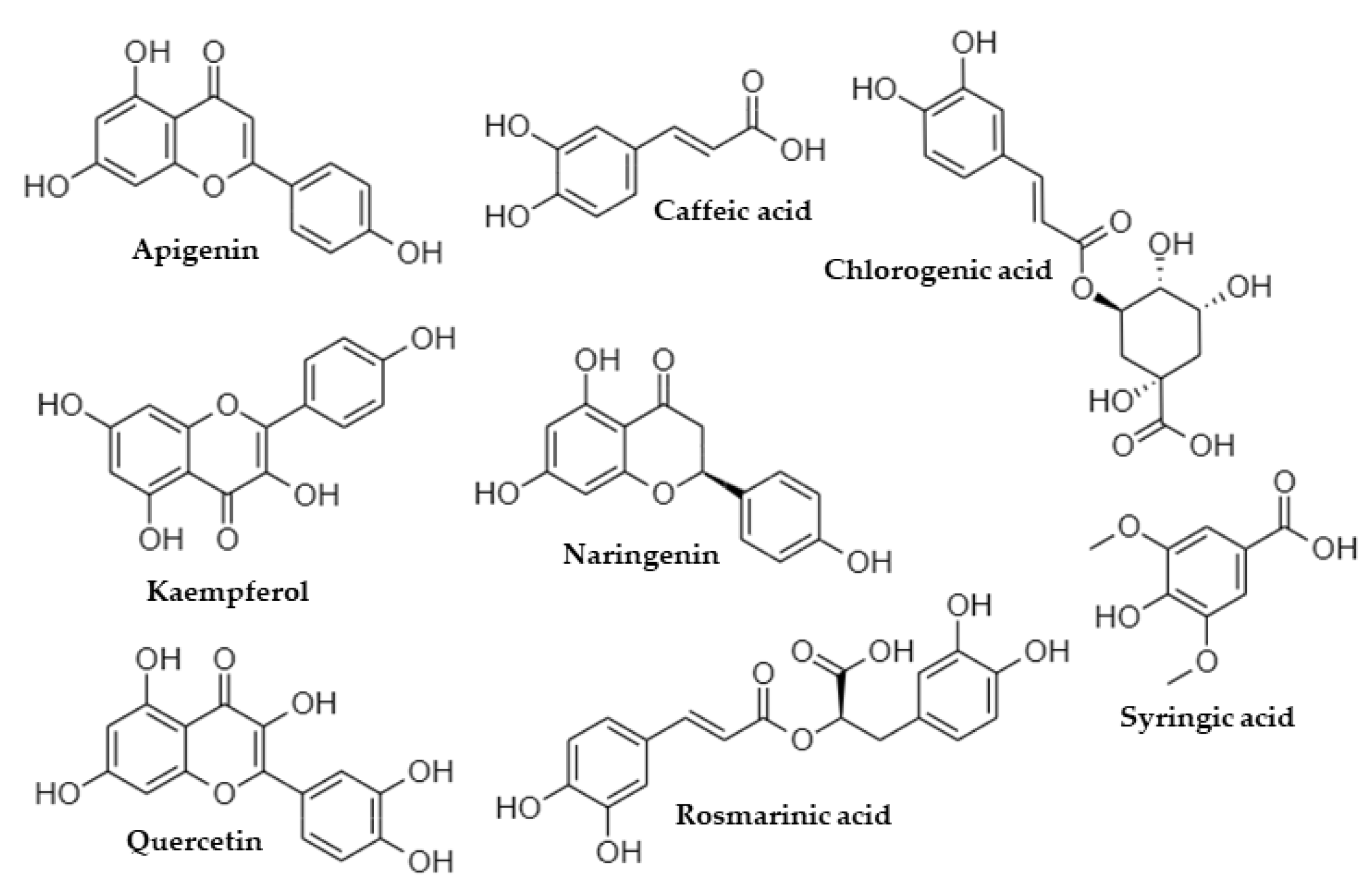
| Phenolic Compound | Retention Time (min) | Parent Ion [M-H]− | Product Ions (MS/MS) |
|---|---|---|---|
| Apigenin | 6.60 | 269.1 | 151.4, 149.3, 117.2 * |
| Caffeic acid | 2.75 | 179.1 | 135.4 *, 107.3 |
| Chlorogenic acid | 1.72 | 353.2 | 191.5 *, 179.6, 161.5 |
| Kaempferol | 6.66 | 285.3 | 257.6 *, 229.6, 187.5 |
| Naringenin | 6.64 | 271.1 | 177.4, 151.4, 119.3 *, 107.3, 93.2 |
| Quercetin | 6.20 | 301.1 | 273.5, 179.3, 151.4 * |
| Rosmarinic acid | 4.95 | 359.3 | 197.5, 179.5, 161.5 *, 135.4 |
| Syringic acid | 2.62 | 197.0 | 182.5 *, 167.5, 123.2, 95.2 |
| Storage Period (Days) | Apigenin | Caffeic Acid | Chlorogenic Acid | Kaempferol | Naringenin | Quercetin | Rosmarinic Acid | Syringic Acid |
|---|---|---|---|---|---|---|---|---|
| 2 | √ | √ | √ | √ | ||||
| 4 | √ | √ | √ | √ | ||||
| 7 | √ | √ | √ | |||||
| 9 | √ | √ | √ | √ | √ | |||
| 11 | √ | √ | √ | √ | √ | √ | ||
| 14 | √ | √ | √ | √ | √ | |||
| 17 | √ | √ | √ | √ | √ | |||
| 21 | √ | √ | √ | √ |
| Phenolic Compounds | hCA-II | hCA-XII | ||
|---|---|---|---|---|
| Docking Score (kcal · mol−1) | Distance Zn2+ (Å) | Docking Score (kcal · mol−1) | Distance Zn2+ (Å) | |
| 2-(But-2-yn-1-ylsulfamoyl)-4-sulfamoylbenzoic acid | −4.86 | 1.76 | 1 NT | NT |
| 2,3,5,6-Tetrafluoro-4-(propylthio)benzenesulfonamide | NT | NT | −3.67 | 1.76 |
| Apigenin | −5.70 | 2.23 | −5.64 | 2.01 |
| Caffeic acid | −4.46 | 2.05 | −5.26 | 2.12 |
| Chlorogenic acid | −5.73 | 2.06 | −5.95 | 2.17 |
| Kaempferol | −5.31 | 2.06 | −4.90 | 2.04 |
| Naringenin | −5.32 | 2.05 | −5.02 | 2.00 |
| Quercetin | −5.13 | 2.23 | −5.29 | 2.17 |
| Rosmarinic acid | −4.72 | 2.05 | −5.73 | 2.12 |
| Syringic acid | −5.48 | 2.05 | −4.89 | 2.15 |
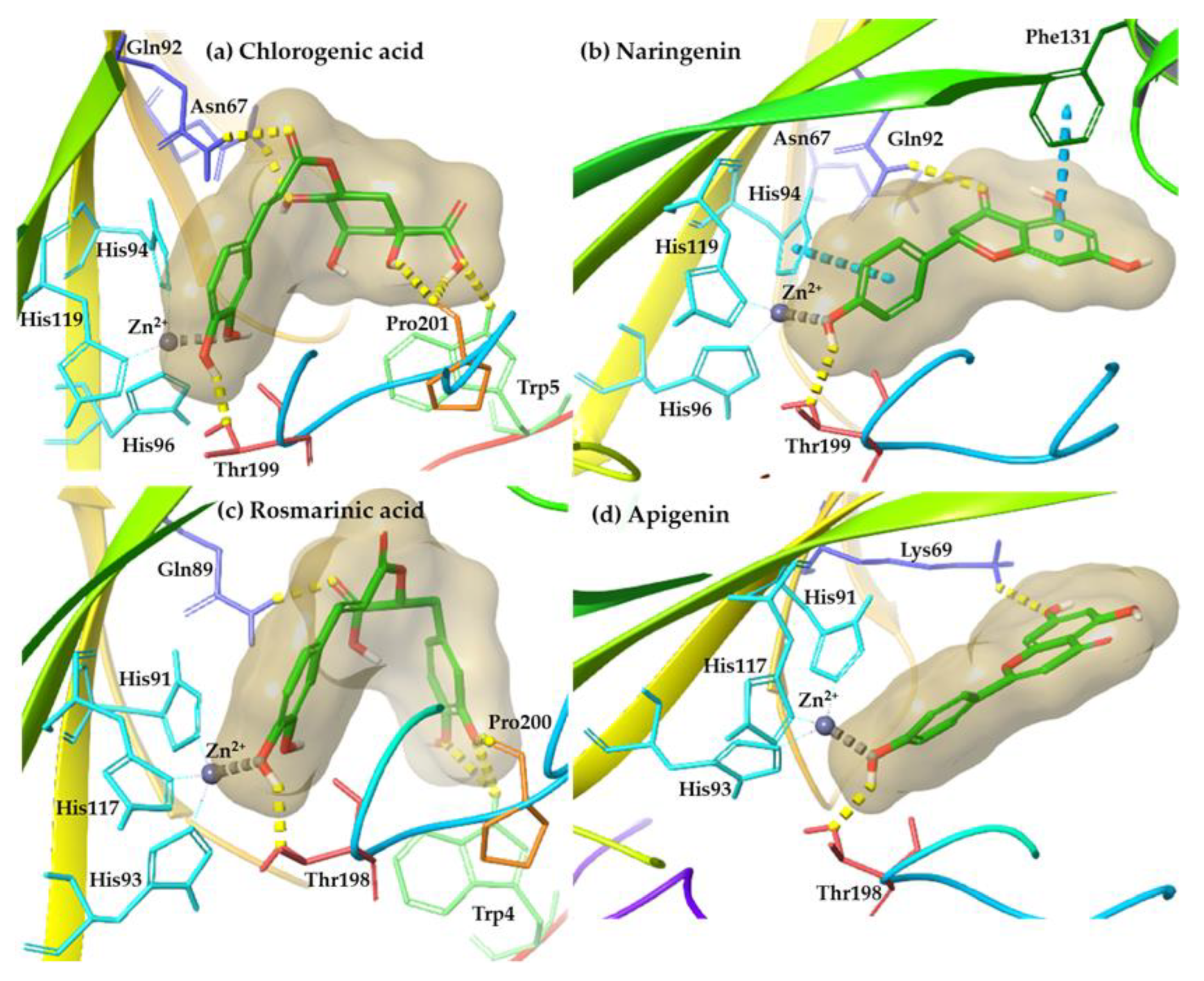
| Phenolic Compounds | hCA-II (PDB ID: 5AML) | hCA-XII (PDB ID: 5MSA) |
|---|---|---|
| Interactions | ||
| 2-(But-2-yn-1-ylsulfamoyl)-4-sulfamoylbenzoic acid | 1 HB: Gln92, Thr199 & 2 mc | |
| 2,3,5,6-Tetrafluoro-4-(propylthio)benzenesulfonamide | HB: Thr198 & mc | |
| Apigenin | HB: Thr199 & mc | HB: Lys69, Thr198 & mc |
| Caffeic acid | HB: Thr199, Asn67 & mc | HB: Thr198 & mc |
| Chlorogenic acid | HB: Trp5, Asn67, Gln92, Thr199, Pro201 & mc | HB: Thr88, Gln89 & mc |
| Kaempferol | HB: Thr199 & mc | mc |
| Naringenin | HB: Gln92, His64, Thr199 & pi-pi Phe131 & mc | HB: Thr198 & mc |
| Quercetin | pi-pi: His94, Phe131 & mc | HB: Thr198, Thr199 & mc |
| Rosmarinic acid | HB: Trp5, Asn67, Gln92, Thr199, Pro201 & mc | HB: Trp4, Gln89, Thr198, Pro200 & mc |
| Syringic acid | HB: Thr199, Thr200 & pi-pi His94 & mc | HB: Gln89, Thr198 & mc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kritsi, E.; Tsiaka, T.; Sotiroudis, G.; Mouka, E.; Aouant, K.; Ladika, G.; Zoumpoulakis, P.; Cavouras, D.; Sinanoglou, V.J. Potential Health Benefits of Banana Phenolic Content during Ripening by Implementing Analytical and In Silico Techniques. Life 2023, 13, 332. https://doi.org/10.3390/life13020332
Kritsi E, Tsiaka T, Sotiroudis G, Mouka E, Aouant K, Ladika G, Zoumpoulakis P, Cavouras D, Sinanoglou VJ. Potential Health Benefits of Banana Phenolic Content during Ripening by Implementing Analytical and In Silico Techniques. Life. 2023; 13(2):332. https://doi.org/10.3390/life13020332
Chicago/Turabian StyleKritsi, Eftichia, Thalia Tsiaka, Georgios Sotiroudis, Elizabeth Mouka, Konstantinos Aouant, Georgia Ladika, Panagiotis Zoumpoulakis, Dionisis Cavouras, and Vassilia J. Sinanoglou. 2023. "Potential Health Benefits of Banana Phenolic Content during Ripening by Implementing Analytical and In Silico Techniques" Life 13, no. 2: 332. https://doi.org/10.3390/life13020332