
Eugenol Induces Apoptosis in Tongue Squamous Carcinoma Cells by Mediating the Expression of Bcl-2 Family

 , , and
, , and 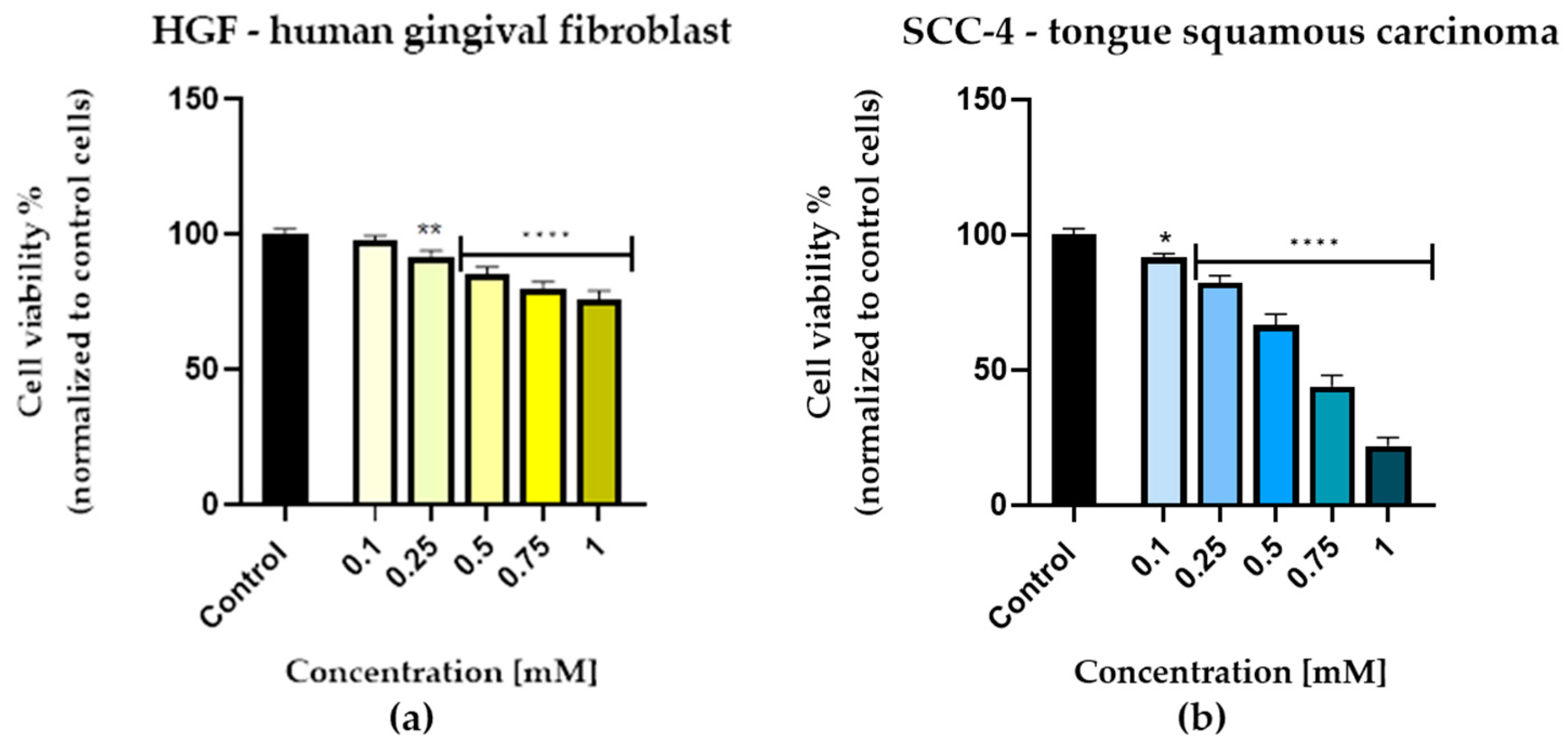{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Culture
2.2. Cell Culture
2.3. Cellular Viability Evaluation
2.4. Cellular, Nuclear, and Cytoskeletal Morphology Evaluation
2.5. Gene Expression
2.6. Statistical Analysis
3. Results
3.1. Cell Viability Evaluation
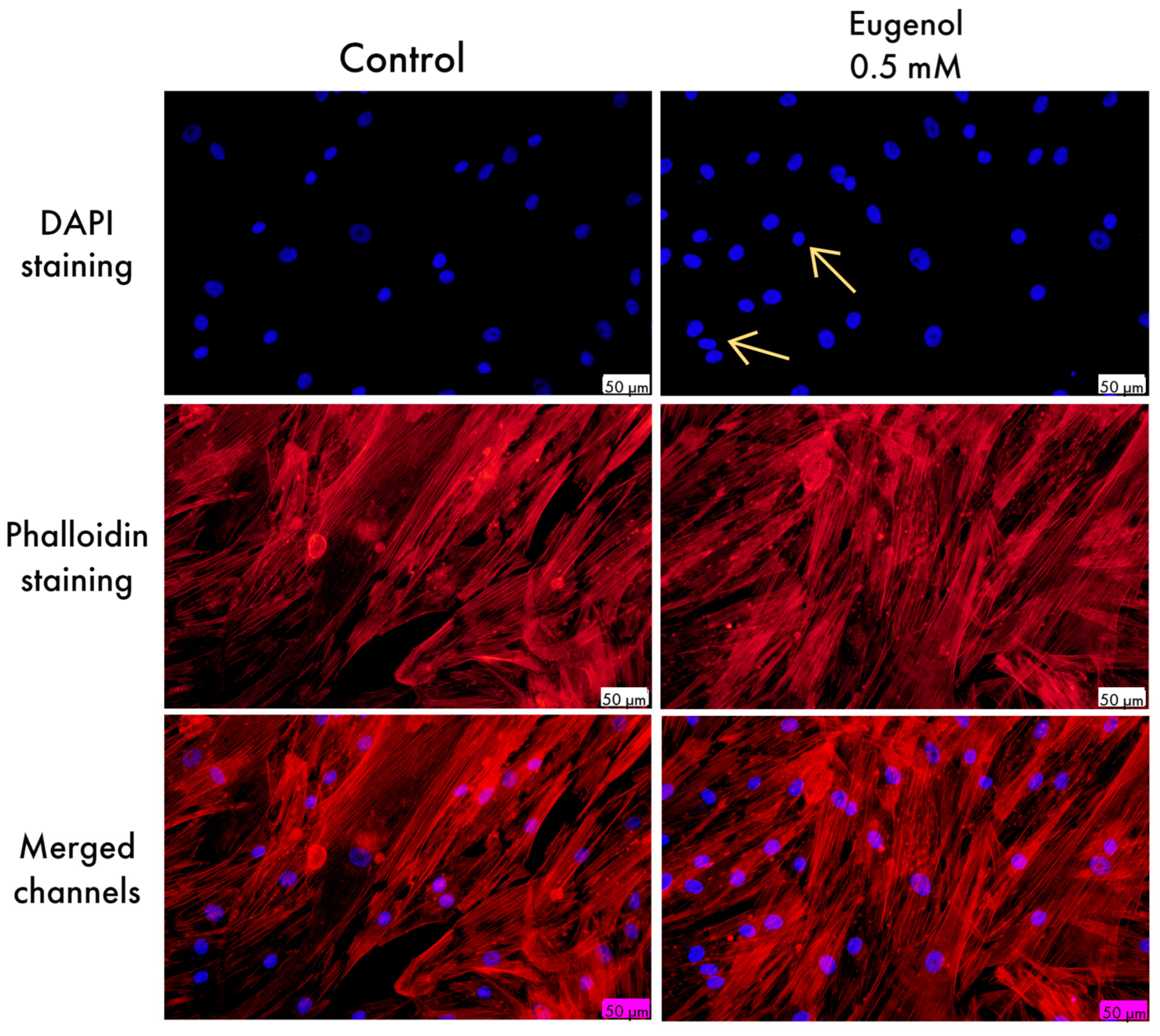
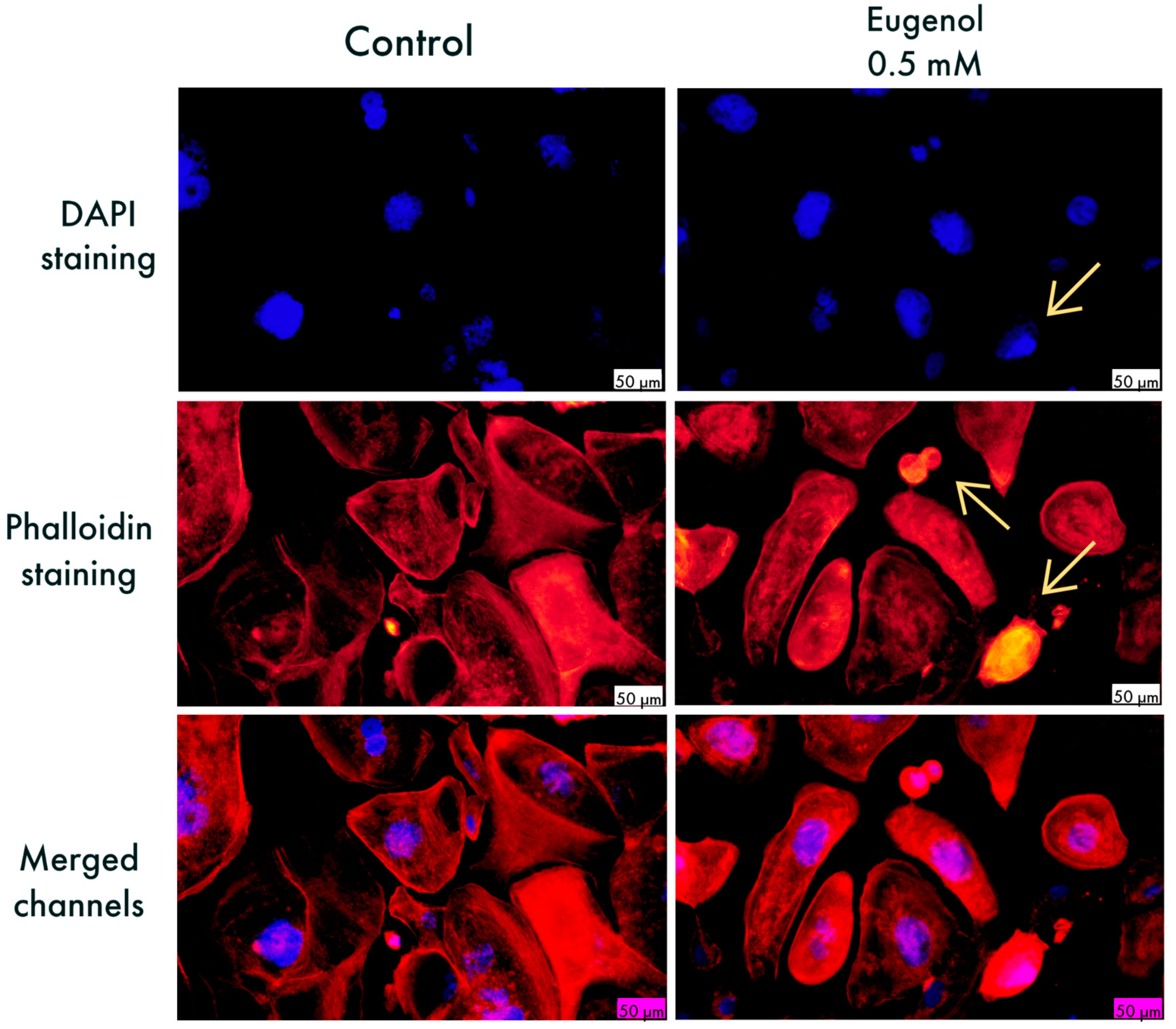
3.2. Cellular, Nuclear, and Cytoskeletal Morphology Evaluation
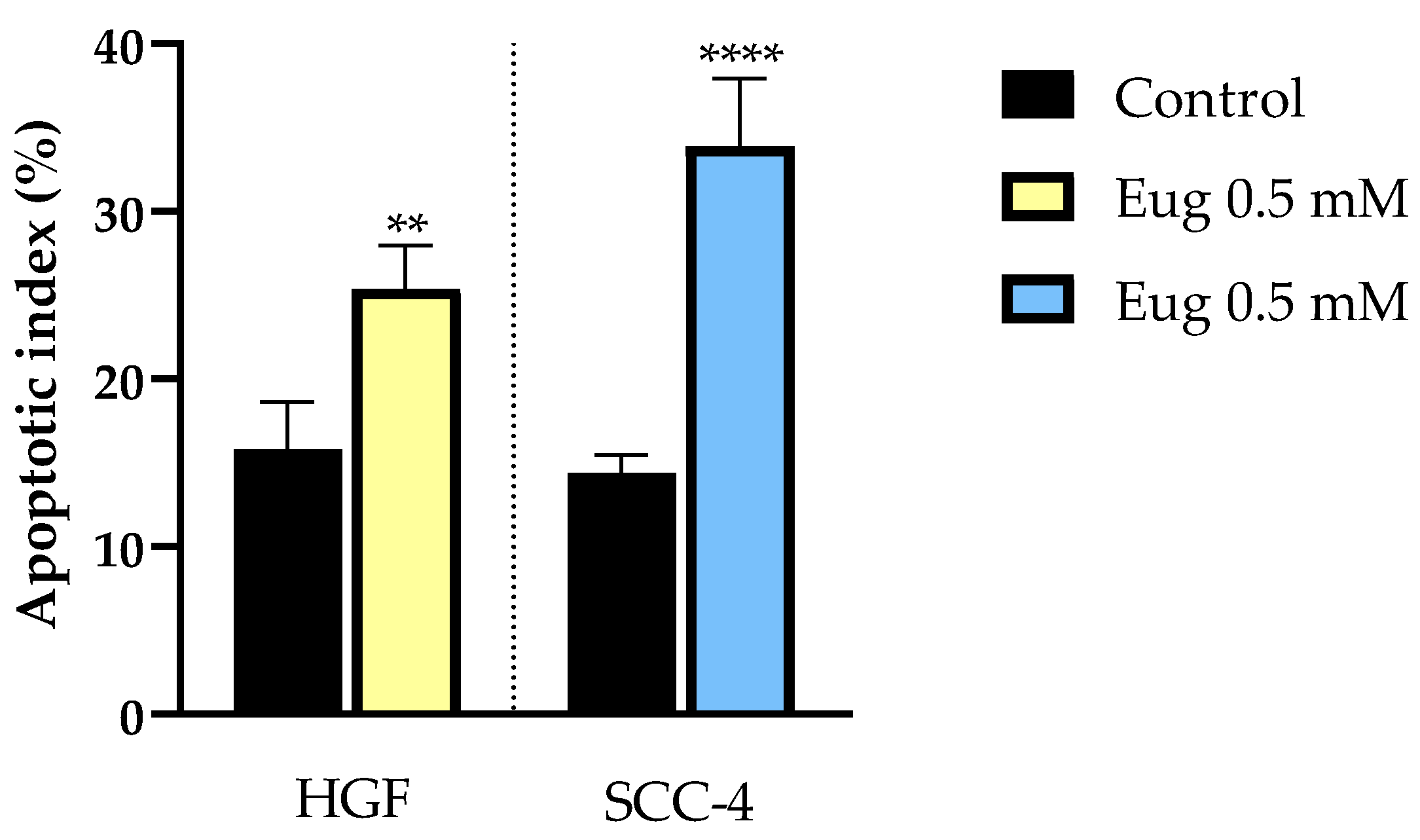
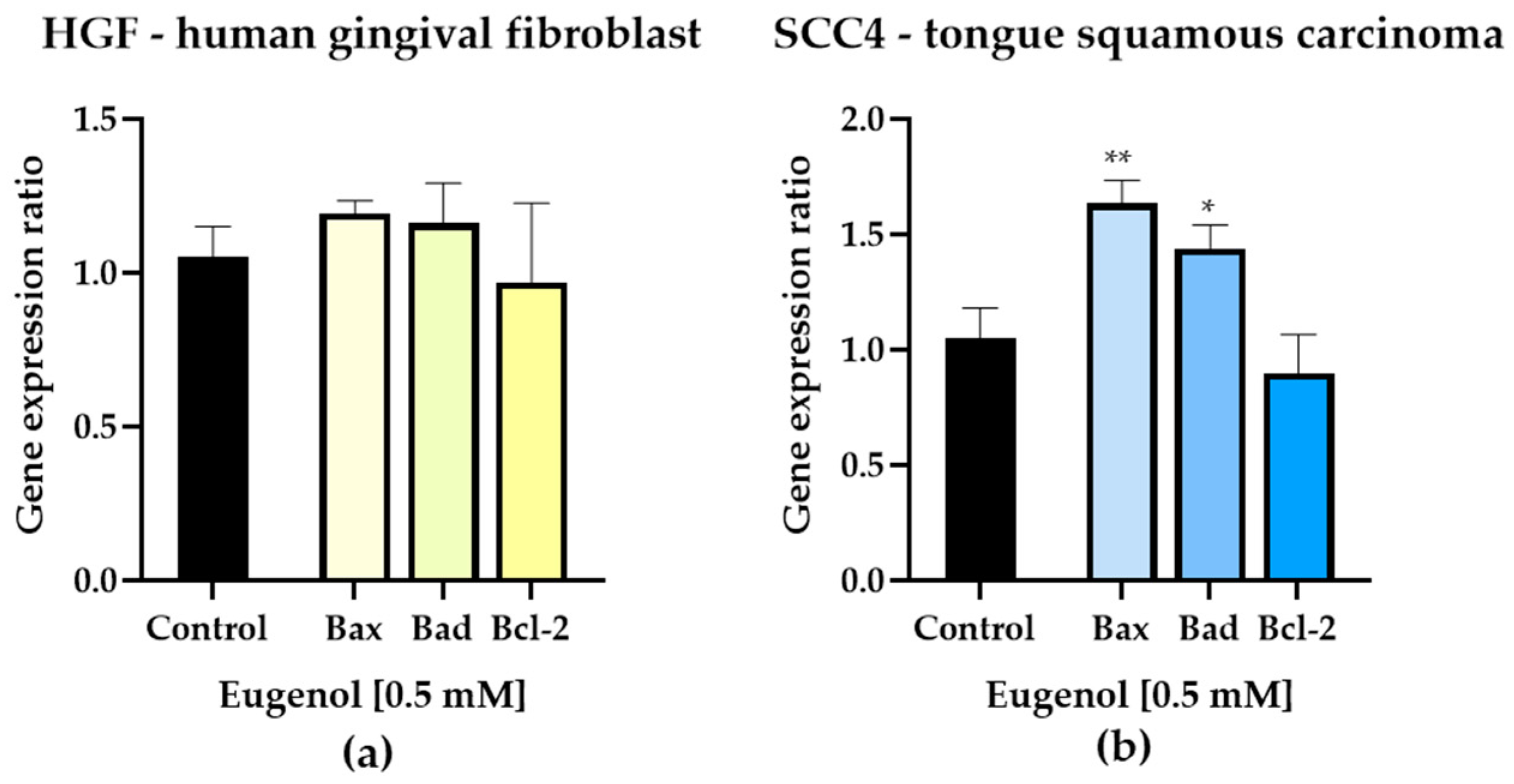
3.3. Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and Neck Squamous Cell Carcinoma. Nat. Rev. Dis. Prim. 2020, 6, 92. [Google Scholar] [CrossRef]
- Pai, S.I.; Westra, W.H. Molecular Pathology of Head and Neck Cancer: Implications for Diagnosis, Prognosis, and Treatment. Annu. Rev. Pathol. 2009, 4, 49–70. [Google Scholar] [CrossRef] [Green Version]
- Woodford, D. An Inflammatory Cytokine Milieu Is Prominent in Premalignant Oral Lesions, but Subsides When Lesions Progress to Squamous Cell Carcinoma. J. Clin. Cell. Immunol. 2014, 5, 230. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.; Lim, J.H.; Lee, H.R.; Jang, J.Y.; Shin, Y.S.; Kim, D.; Lim, J.Y.; Park, Y.M.; Koh, Y.W.; Ahn, S.H.; et al. Deep Learning Model for Tongue Cancer Diagnosis Using Endoscopic Images. Sci. Rep. 2022, 12, 1–10. [Google Scholar] [CrossRef]
- Mannelli, G.; Arcuri, F.; Agostini, T.; Innocenti, M.; Raffaini, M.; Spinelli, G. Classification of Tongue Cancer Resection and Treatment Algorithm. J. Surg. Oncol. 2018, 117, 1092–1099. [Google Scholar] [CrossRef]
- Alsahafi, E.; Begg, K.; Amelio, I.; Raulf, N.; Lucarelli, P.; Sauter, T.; Tavassoli, M. Clinical Update on Head and Neck Cancer: Molecular Biology and Ongoing Challenges. Cell Death Dis. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Andreadis, C.; Vahtsevanos, K.; Sidiras, T.; Thomaidis, I.; Antoniadis, K.; Mouratidou, D. 5-Fluorouracil and Cisplatin in the Treatment of Advanced Oral Cancer. Oral. Oncol. 2003, 39, 380–385. [Google Scholar] [CrossRef]
- Kaidar-Person, O.; Gil, Z.; Billan, S. Precision Medicine in Head and Neck Cancer. Drug Resist. Updat. 2018, 40, 13–16. [Google Scholar] [CrossRef]
- Scotte, F.; Ratta, R.; Beuzeboc, P. Side Effects of Immunotherapy: A Constant Challenge for Oncologists. Curr. Opin. Oncol. 2019, 31, 280–285. [Google Scholar] [CrossRef]
- Talib, W.H.; Awajan, D.; Hamed, R.A.; Azzam, A.O.; Mahmod, A.I.; AL-Yasari, I.H. Combination Anticancer Therapies Using Selected Phytochemicals. Molecules 2022, 27, 5452. [Google Scholar] [CrossRef]
- Ulanowska, M.; Olas, B. Biological Properties and Prospects for the Application of Eugenol—A Review. Int. J. Mol. Sci. 2021, 22, 3671. [Google Scholar] [CrossRef]
- Zari, A.T.; Zari, T.A.; Hakeem, K.R. Anticancer Properties of Eugenol: A Review. Molecules 2021, 26, 7407. [Google Scholar] [CrossRef]
- Barboza, J.N.; da Silva Maia Bezerra Filho, C.; Silva, R.O.; Medeiros, J.V.R.; de Sousa, D.P. An Overview on the Anti-Inflammatory Potential and Antioxidant Profile of Eugenol. Oxid. Med. Cell. Longev. 2018, 2018, 3957262. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative Stress, Inflammation, and Cancer: How Are They Linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Herrera, A.; Pozos-Guillén, A.; Ruiz-Rodríguez, S.; Garrocho-Rangel, A.; Vértiz-Hernández, A.; Escobar-García, D.M. Effect of 4-Allyl-1-Hydroxy-2-Methoxybenzene (Eugenol) on Inflammatory and Apoptosis Processes in Dental Pulp Fibroblasts. Mediat. Inflamm. 2016, 2016, 9371403. [Google Scholar] [CrossRef]
- Marya, C.M.; Satija, G.; Avinash, J.; Nagpal, R.; Kapoor, R.; Ahmad, A. In Vitro Inhibitory Effect of Clove Essential Oil and Its Two Active Principles on Tooth Decalcification by Apple Juice. Int. J. Dent. 2012, 2012, 759618. [Google Scholar] [CrossRef] [Green Version]
- Jeng, J.H.; Hahn, L.J.; Lu, F.J.; Wang, Y.J. Eugenol Triggers Different Pathobiological Effects on Human Oral Mucosal Fibroblasts 1. J. Dent. Res. 1994, 73, 1050–1055. [Google Scholar] [CrossRef]
- Dehelean, C.A.; Coricovac, D.; Pinzaru, I.; Marcovici, I.; Macasoi, I.G.; Semenescu, A.; Lazar, G.; Cinta Pinzaru, S.; Radulov, I.; Alexa, E.; et al. Rutin Bioconjugates as Potential Nutraceutical Prodrugs: An in Vitro and in Ovo Toxicological Screening. Front. Pharmacol. 2022, 13, 1–20. [Google Scholar] [CrossRef]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Cell Viability Assays. Assay Guid. Man. 2016. [Google Scholar]
- Atale, N.; Gupta, S.; Yadav, U.C.S.; Rani, V. Cell-Death Assessment by Fluorescent and Nonfluorescent Cytosolic and Nuclear Staining Techniques. J. Microsc. 2014, 255, 7–19. [Google Scholar] [CrossRef]
- Mazloom-Farsibaf, H.; Farzam, F.; Fazel, M.; Wester, M.J.; Meddens, M.B.M.; Lidke, K.A. Comparing Lifeact and Phalloidin for Superresolution Imaging of Actin in Fixed Cells. PLoS ONE 2021, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pancu, D.F.; Racea, R.C.; Macasoi, I.; Sarau, C.A.; Pinzaru, I.; Poenaru, M.; Rusu, L.C.; Dehelean, C.A.; Dinu, S. Assessment of the In Vitro Cytotoxic Profile of Two Broad-Spectrum Antibiotics—Tetracycline and Ampicillin—On Pharyngeal Carcinoma Cells. Medicina 2022, 58, 1289. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Sun, G.; Yuan, G.; Wang, R.; Sun, X. Effects of Platycodin D on Proliferation, Apoptosis and PI3K/Akt Signal Pathway of Human Glioma U251 Cells. Molecules 2014, 19, 21411–21423. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- ANSI/AAMI/ISO 10993-5:2009; Biological Evaluation of Medical Devices—Part 5: Tests for In Vitro Cytotoxicity. AAMI: Arlington, VA, USA, 2009.
- Kamiloglu, S.; Sari, G.; Ozdal, T.; Capanoglu, E. Guidelines for Cell Viability Assays. Food Front. 2020, 1, 332–349. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Park, B.-S. The Effect of Eugenol on the Induction of Apoptosis in HSC-2 Human Oral Squamous Cell Carcinoma. J. Korean Soc. Dent. Hyg. 2015, 15, 523–529. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.; Park, J.; Kim, G.; Park, B.; Gil, Y.; Kim, C. The Mechanism of Apoptosis Induced by Eugenol in Human Osteosarcoma Cells. J. Korean Assoc. Oral Maxillofac. Surg. 2007, 33, 20–27. [Google Scholar]
- Jaganathan, S.K.; Mazumdar, A.; Mondhe, D.; Mandal, M. Apoptotic Effect of Eugenol in Human Colon Cancer Cell Lines. Cell Biol. Int. 2011, 35, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Ghodousi-Dehnavi, E.; Hosseini, R.H.; Arjmand, M.; Nasri, S.; Zamani, Z. A Metabolomic Investigation of Eugenol on Colorectal Cancer Cell Line HT-29 by Modifying the Expression of APC, P53, and KRAS Genes. Evid. Based Complement. Altern. Med. 2021, 2021, 1448206. [Google Scholar] [CrossRef]
- Begum, S.N.; Ray, A.S.; Rahaman, C.H. A Comprehensive and Systematic Review on Potential Anticancer Activities of Eugenol: From Pre-Clinical Evidence to Molecular Mechanisms of Action. Phytomedicine 2022, 107, 154456. [Google Scholar] [CrossRef]
- Ren, W.; Zhao, W.; Cao, L.; Huang, J. Involvement of the Actin Machinery in Programmed Cell Death. Front. Cell Dev. Biol. 2021, 8, 634849. [Google Scholar] [CrossRef]
- Júnior, P.L.D.S.; Câmara, D.A.D.; Costa, A.S.; Ruiz, J.L.M.; Levy, D.; Azevedo, R.A.; Pasqualoto, K.F.M.; De Oliveira, C.F.; De Melo, T.C.; Pessoa, N.D.S.; et al. Apoptotic Effect of Eugenol Envolves G2/M Phase Abrogation Accompanied by Mitochondrial Damage and Clastogenic Effect on Cancer Cell in Vitro. Phytomedicine 2016, 23, 725–735. [Google Scholar] [CrossRef]
- Ndozangue-Touriguine, O.; Hamelin, J.; Bréard, J. Cytoskeleton and Apoptosis. Biochem. Pharmacol. 2008, 76, 11–18. [Google Scholar] [CrossRef]
- Xu, X.; Lai, Y.; Hua, Z.C. Apoptosis and Apoptotic Body: Disease Message and Therapeutic Target Potentials. Biosci. Rep. 2019, 39, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Doncel, J.P.; de la Cruz Ojeda, P.; OropesaÁvila, M.; Paz, M.V.; De Lavera, I.; La Mata, M.D.; Córdoba, M.Á.; Hidalgo, R.L.; Rivero, J.M.S.; Cotán, D.; et al. Cytoskeleton Rearrangements during the Execution Phase of Apoptosis. Cytoskelet. Struct. Dyn. Funct. Dis. 2017, 151–169. [Google Scholar] [CrossRef] [Green Version]
- Abraham, M.T.; Kuriakose, M.A.; Sacks, P.G.; Yee, H.; Chiriboga, L.; Bearer, E.L.; Delacure, M.D. Motility-Related Proteins as Markers for Head and Neck Squamous Cell Cancer. Laryngoscope 2001, 111, 1285–1289. [Google Scholar] [CrossRef] [Green Version]
- Povea-Cabello, S.; Oropesa-Ávila, M.; de la Cruz-Ojeda, P.; Villanueva-Paz, M.; De La Mata, M.; Suárez-Rivero, J.M.; Álvarez-Córdoba, M.; Villalón-García, I.; Cotán, D.; Ybot-González, P.; et al. Dynamic Reorganization of the Cytoskeleton during Apoptosis: The Two Coffins Hypothesis. Int. J. Mol. Sci. 2017, 18, 2393. [Google Scholar] [CrossRef] [Green Version]
- An, W.; Lai, H.; Zhang, Y.; Liu, M.; Lin, X.; Cao, S. Apoptotic Pathway as the Therapeutic Target for Anticancer Traditional Chinese Medicines. Front. Pharmacol. 2019, 10, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Ketelut-Carneiro, N.; Fitzgerald, K.A. Apoptosis, Pyroptosis, and Necroptosis—Oh My! The Many Ways a Cell Can Die. J. Mol. Biol. 2022, 434, 167378. [Google Scholar] [CrossRef]
- Kale, J.; Osterlund, E.J.; Andrews, D.W. BCL-2 Family Proteins: Changing Partners in the Dance towards Death. Cell Death Differ. 2018, 25, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Fathy, M.; Fawzy, M.A.; Hintzsche, H.; Nikaido, T.; Dandekar, T.; Othman, E.M. Eugenol Exerts Apoptotic Effect and Modulates the Sensitivity of HeLa Cells to Cisplatin and Radiation. Molecules 2019, 24, 3979. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surducan, D.-A.; Racea, R.C.; Cabuta, M.; Olariu, I.; Macasoi, I.; Rusu, L.C.; Chiriac, S.D.; Chioran, D.; Dinu, S.; Pricop, M.O. Eugenol Induces Apoptosis in Tongue Squamous Carcinoma Cells by Mediating the Expression of Bcl-2 Family. Life 2023, 13, 22. https://doi.org/10.3390/life13010022
Surducan D-A, Racea RC, Cabuta M, Olariu I, Macasoi I, Rusu LC, Chiriac SD, Chioran D, Dinu S, Pricop MO. Eugenol Induces Apoptosis in Tongue Squamous Carcinoma Cells by Mediating the Expression of Bcl-2 Family. Life. 2023; 13(1):22. https://doi.org/10.3390/life13010022
Chicago/Turabian StyleSurducan, Dan-Alexandru, Robert Cosmin Racea, Madalina Cabuta, Iustin Olariu, Ioana Macasoi, Laura Cristina Rusu, Sorin Dan Chiriac, Doina Chioran, Stefania Dinu, and Marius Octavian Pricop. 2023. "Eugenol Induces Apoptosis in Tongue Squamous Carcinoma Cells by Mediating the Expression of Bcl-2 Family" Life 13, no. 1: 22. https://doi.org/10.3390/life13010022