
Newly Synthesized Creatine Derivatives as Potential Neuroprotective and Antioxidant Agents on In Vitro Models of Parkinson’s Disease

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Materials

2.3. Isolation of Rat Brain Synaptosomes and Mitochondria
2.4. Synaptosomal Viability
2.5. Determination of GSH Levels in Synaptosomes and Mitochondria
2.6. Isolation of Brain Microsomes
2.7. Determination of MDA in Brain Mitochondria and Microsomes
2.8. Statistical Analysis
3. Results
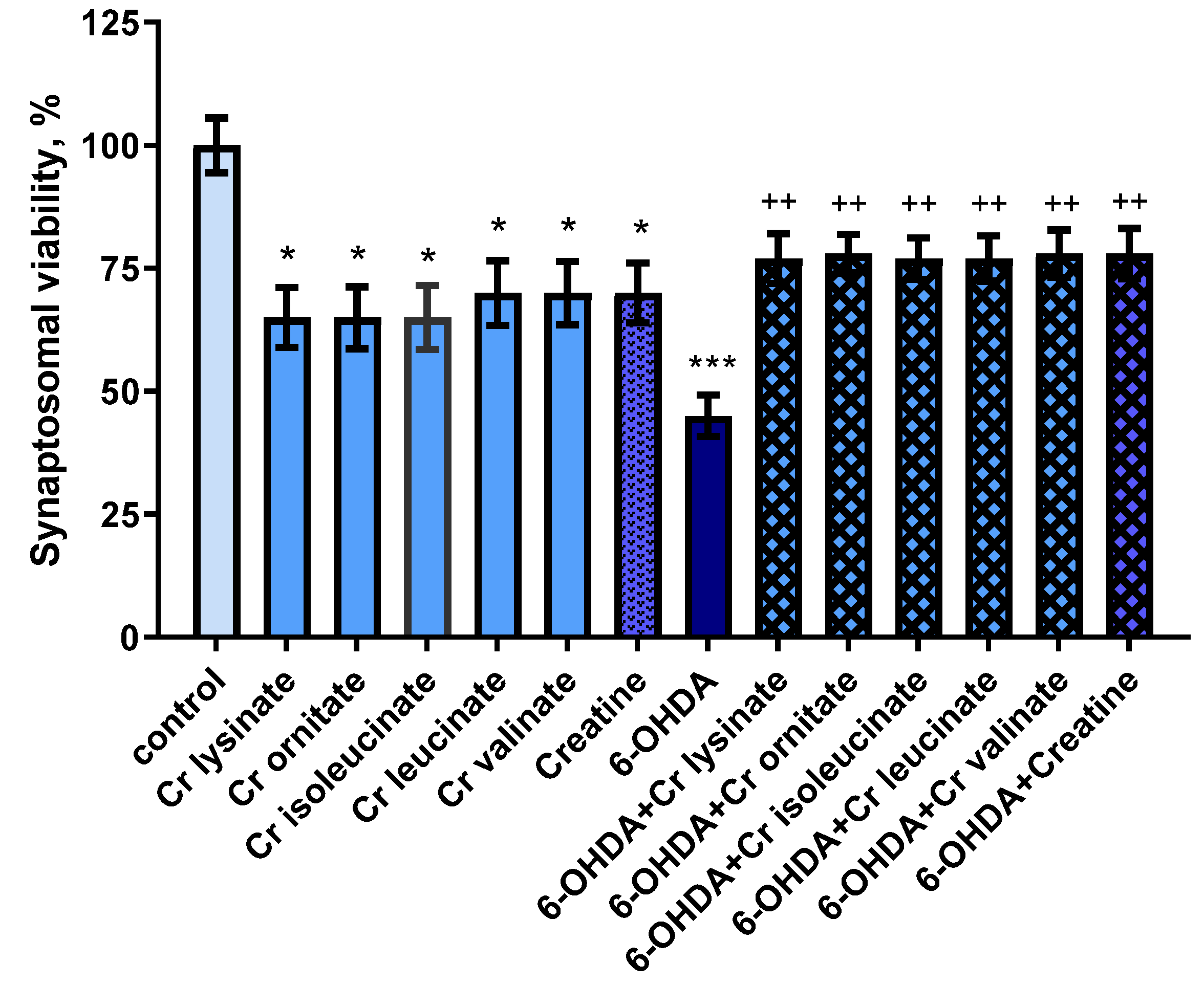
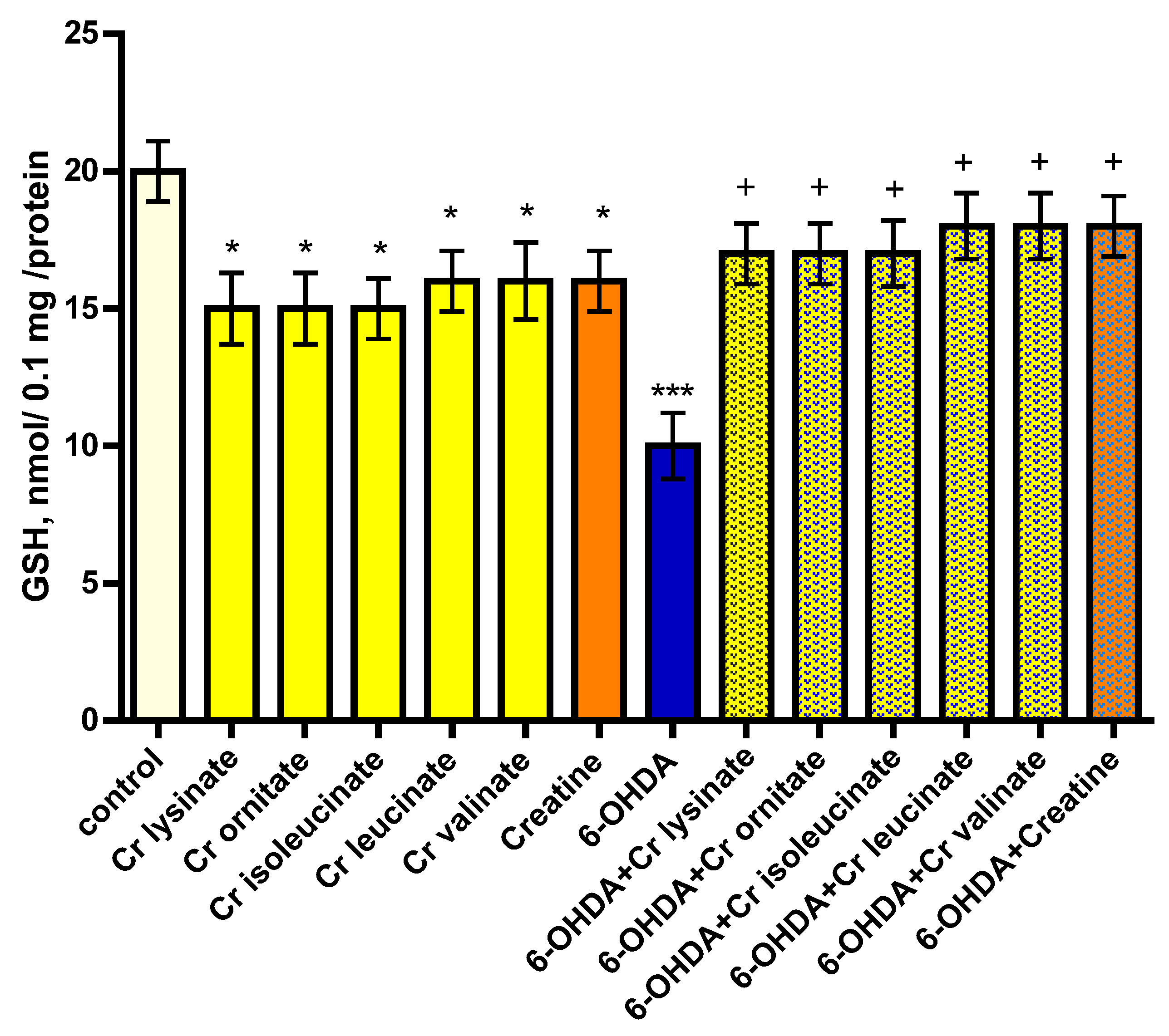
3.1. Effects of Creatine Derivatives (at Concentration 38 µM) on Isolated Rat Brain Synaptosomes
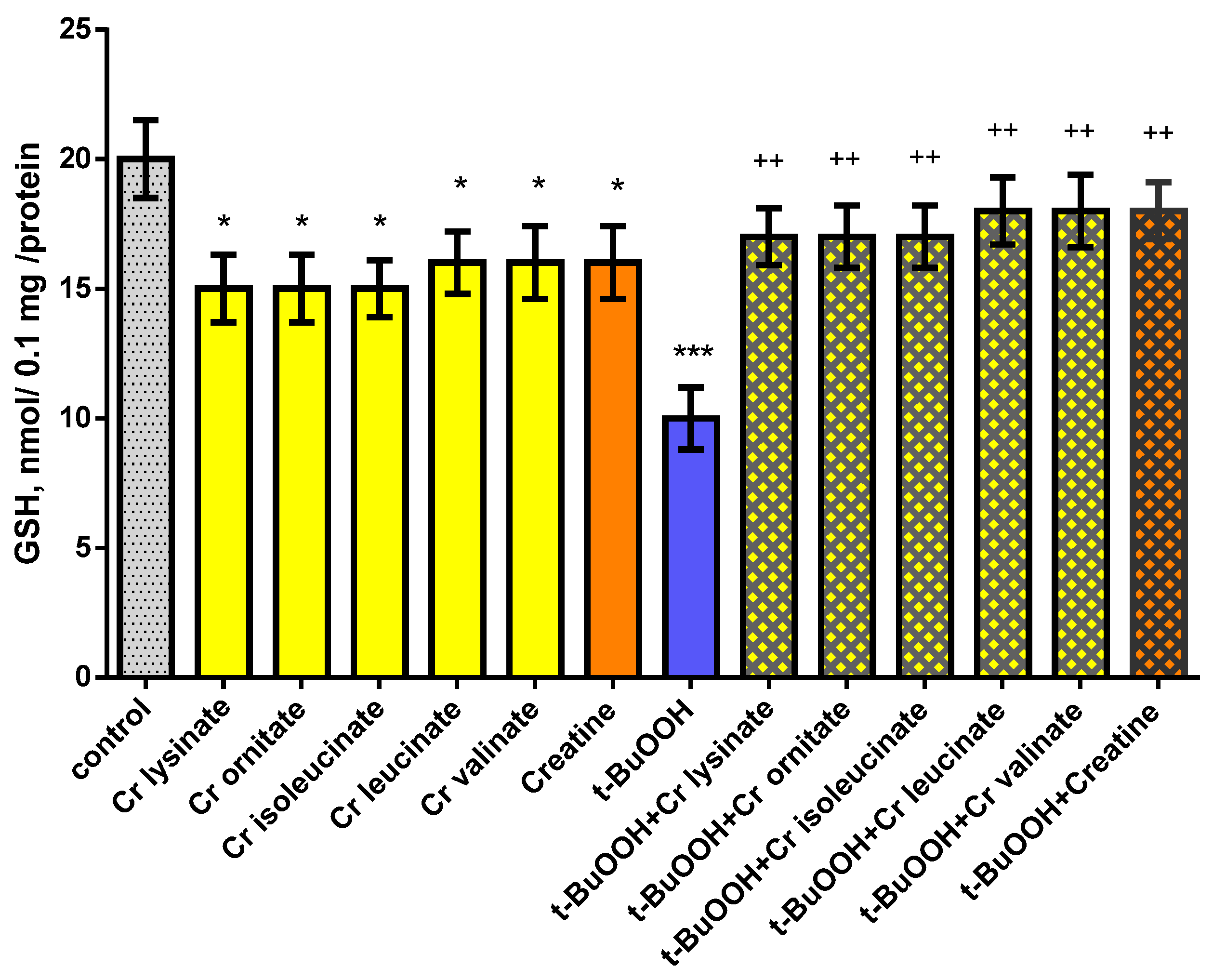
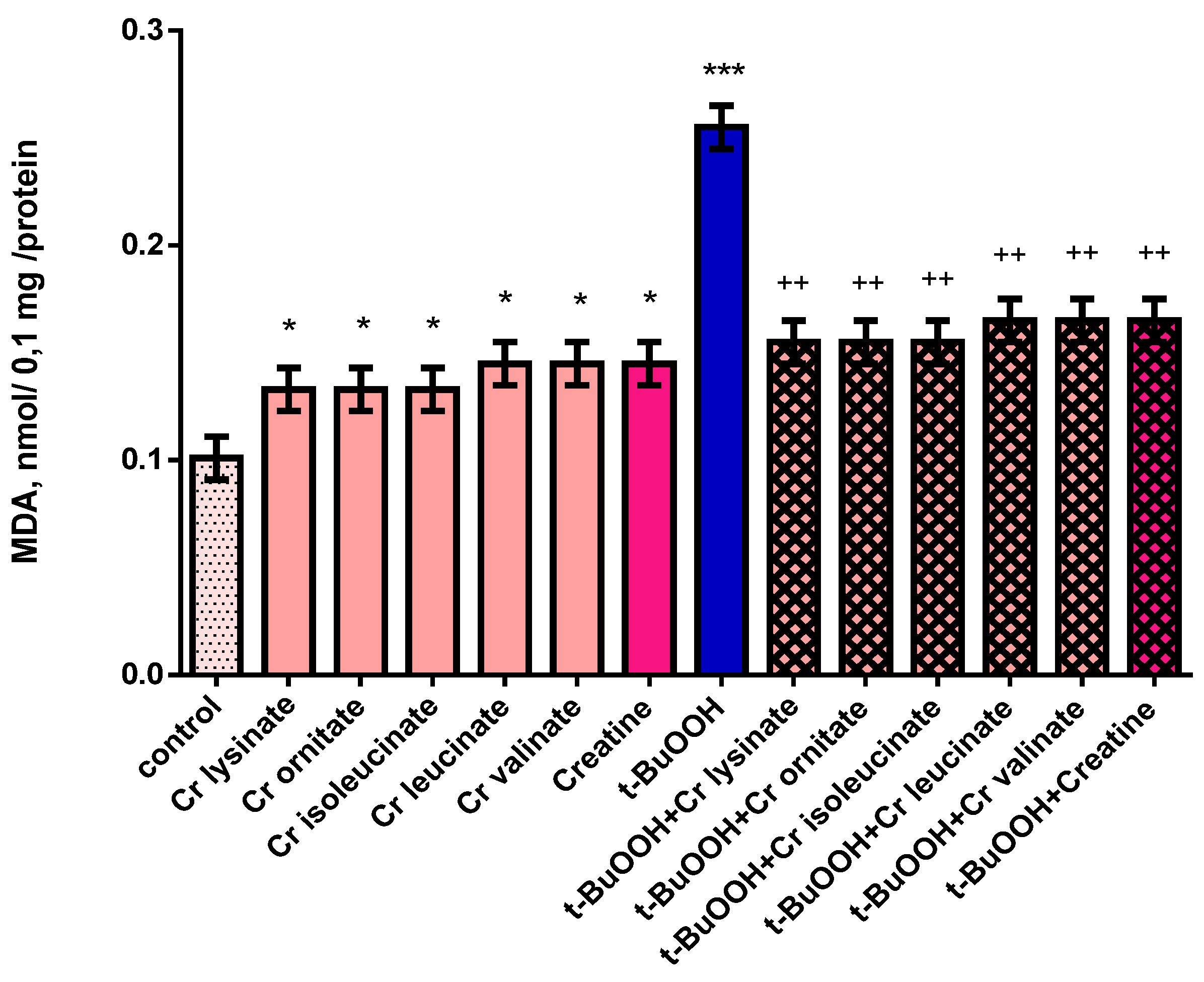
3.2. Effects of Creatine Derivatives (at Concentration 38 µM) on Isolated Brain Mitochondria
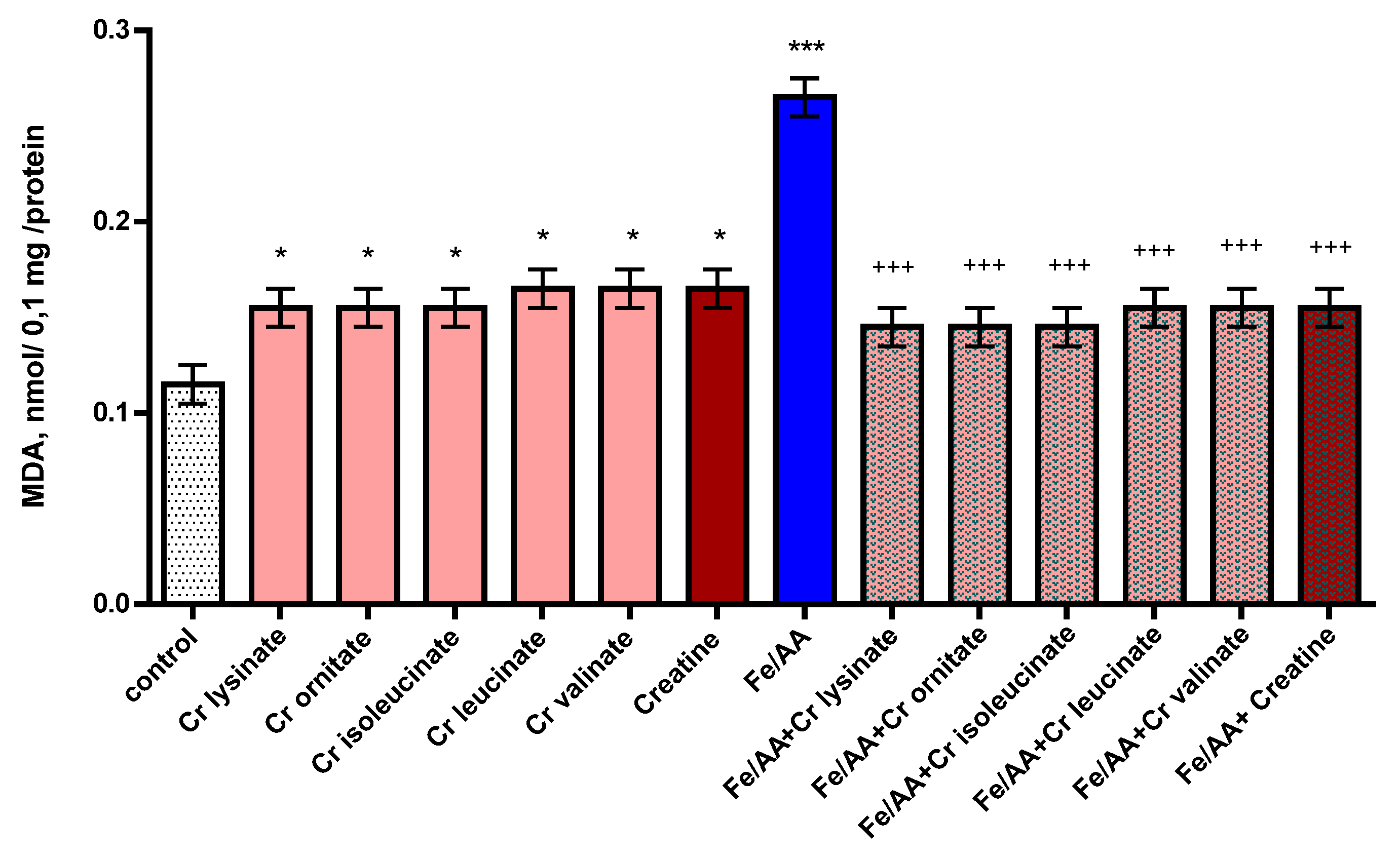
3.3. Effects of Creatine Derivatives (at Concentration 38 µM) on Isolated Brain Microsomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brooks, D.J. The early diagnosis of Parkinsons disease. Ann. Neurol. 1998, 44, S10–S18. [Google Scholar] [CrossRef] [PubMed]
- Bergman, H.; Deuschl, G. Pathophysiology of Parkinson’s disease: From clinical neurology to basic neuroscience and back. Mov. Disord. 2002, 17, S28–S40. [Google Scholar] [CrossRef] [PubMed]
- Lo Bianco, C.; Schneider, B.L.; Bauer, M.; Sajadi, A.; Brice, A.; Iwatsubo, T.; Aebischer, P. Lentiviral vector delivery of parkin prevents dopaminergic degeneration in an alpha-synuclein rat model of Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2004, 101, 17510–17515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, R.L.; James-Kracke, M.; Sun, G.Y.; Sun, A.Y. Oxidative and inflammatory pathways in Parkinson’s disease. Neurochem. Res. 2009, 34, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Tobón-Velasco, J.C.; Carmona-Aparicio, L.; Ali, S.F.; Santamaría, A. Biomarkers of cell damage induced by oxidative stress in Parkinson’s disease and related models. Cent. Nerv. Syst. Agents Med. Chem. 2010, 10, 278–286. [Google Scholar] [CrossRef]
- Leal, P.C.; Lins, L.C.; de Gois, A.M.; Marchioro, M.; Santos, J.R. Commentary: Evaluation of Models of Parkinson’s Disease. Front. Neurosci. 2016, 10, 283. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.C.; Li, J.Y.; Tan, H.H.; Du, C.X.; Xie, W.; Zhang, Y.M.; Ma, W.L.; Zhang, L. Serotonin6 receptors in the dorsal hippocampus regulate depressive-like behaviors in unilateral 6-hydroxydopamine-lesioned Parkinson’s rats. Neuropharmacology 2015, 95, 290–298. [Google Scholar] [CrossRef]
- Massari, C.M.; Castro, A.A.; Dal-Cim, T.; Lanznaster, D.; Tasca, C.I. In vitro 6-hydroxydopamine-induced toxicity in striatal, cere-brocortical and hippocampal slices is attenuated by atorvastatin and MK-801. Toxicol. In Vitro 2016, 37, 162–168. [Google Scholar] [CrossRef]
- Hu, M.; Li, F.; Wang, W. Vitexin protects dopaminergic neurons in MPTP-induced Parkinson’s disease through PI3K/Akt signaling pathway. Drug Des. Devel. Ther. 2018, 12, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Xicoy, H.; Wieringa, B.; Martens, G.J. The SH-SY5Y cell line in Parkinson’s disease research: A systematic review. Mol. Neurodegener. 2017, 12, 10. [Google Scholar] [CrossRef]
- Choi, S.H.; Kim, Y.H.; Hebisch, M.; Sliwinski, C.; Lee, S.; D’Avanzo, C.; Chen, H.; Hooli, B.; Asselin, C.; Muffat, J.; et al. A three-dimensional human neural cell culture model of Alzheimer’s disease. Nature 2014, 515, 274–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechler, M.E.; Byrne, L.; Ffrench-Constant, C. CNS Myelin Sheath Lengths Are an Intrinsic Property of Oligodendrocytes. Curr. Biol. 2015, 25, 2411–2416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blandini, F.; Armentero, M.T. Animal models of Parkinson’s disease. FEBS J. 2012, 279, 1156–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, D.; Torch, S.; Lambeng, N.; Nissou, M.; Benabid, A.L.; Sadoul, R.; Verna, J.-M. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: Contribution to the apoptotic theory in Parkinson’s disease. Prog. Neurobiol. 2001, 65, 135–172. [Google Scholar] [CrossRef]
- Chiba, K.; Trevor, A.; Castagnoli, N., Jr. Metabolism of the neurotoxic tertiary amine, MPTP, by brain monoamine oxidase. Biochem. Biophys. Res. Commun. 1984, 120, 574–578. [Google Scholar] [CrossRef]
- Jenner, P.; Schapira, A.H.; Marsden, C.D. New insights into the cause of Parkinson’s disease. Neurology 1992, 42, 2241–2250. [Google Scholar] [CrossRef]
- Salimi, A.; Jamali, Z.; Shabani, M. Antioxidant Potential and Inhibition of Mitochondrial Permeability Transition Pore by Myricetin Reduces Aluminium Phosphide-Induced Cytotoxicity and Mitochondrial Impairments. Front. Pharmacol. 2021, 12, 719081. [Google Scholar] [CrossRef]
- Mansuy, D.; Sassi, A.; Dansette, P.M.; Plat, M. A new potent inhibitor of lipid peroxidation in vitro and in vivo, the hepatoprotective drug anisyldithiolthione. Biochem. Biophys. Res. Commun. 1986, 135, 1015–1021. [Google Scholar] [CrossRef]
- Harrison, J.F.; Hollensworth, S.B.; Spitz, D.R.; Copeland, W.C.; Wilson, G.L.; LeDoux, S.P. Oxidative stress-induced apoptosis in neurons correlates with mitochondrial DNA base excision repair pathway imbalance. Nucleic Acids Res. 2005, 33, 4660–4671. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, S.; Dikshit, M. Involvement of the mitochondrial apoptotic pathway and nitric oxide synthase in dopaminergic neuronal death induced by 6-hydroxydopamine and lipopolysaccharide. Redox Rep. 2010, 15, 115–122. [Google Scholar] [CrossRef]
- Arazi, H.; Eghbali, E.; Suzuki, K. Creatine Supplementation, Physical Exercise and Oxidative Stress Markers: A Review of the Mechanisms and Effectiveness. Nutrients 2021, 13, 869. [Google Scholar] [CrossRef] [PubMed]
- Taupin, P.; Zini, S.; Cesselin, F.; Ben-Ari, Y.; Roisin, M.P. Subcellular fractionation on Percoll gradient of mossy fiber synaptosomes: Morphological and biochemical characterization in control and degranulated rat hippocampus. J. Neurochem. 1994, 62, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Sims, N.R.; Anderson, M.F. Isolation of mitochondria from rat brain using Percoll density gradient centrifugation. Nat. Protoc. 2008, 3, 1228–1239. [Google Scholar] [CrossRef]
- Kondeva-Burdina, M.; Voynova, M.; Aluani, D.; Tzankova, V.; Krasteva, I. Effects of Amanita muscaria extract on different in vitro neurotoxicity models at sub-cellular and cellular levels. Food Chem. Toxicol. 2019, 132, 110687. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, J.; Emgard, M.; Brundin, P.; Burkitt, M.J. Trans-Resveratrol protects embryonic mesencephalic cells from tert-butyl hydroperoxide: Electron paramagnetic resonance spin trapping evidence for radical scavenging mechanism. J. Neurochem. 2000, 75, 141–150. [Google Scholar] [CrossRef]
- Mungarro-Menchaca, X.; Ferrera, P.; Morán, J.; Arias, C. β-Amyloid peptide induces ultrastructural changes in synaptosomes and potentiates mitochondrial dysfunction in the presence of ryanodine. J. Neurosci. Res. 2002, 68, 89–96. [Google Scholar] [CrossRef]
- Robyt, J.; Ackerman, R.; Chittenden, C. Reaction of protein disulfide groups with Ellman’s reagent: A case study of the number of sulfhydryl and disulfide groups in Aspergillus oryzae α-amylase, papain, and lysozyme. Arch. Biochem. Biophys. 1971, 147, 262–269. [Google Scholar] [CrossRef]
- Shirani, M.; Alizadeh, S.; Mahdavinia, M.; Dehghani, M.A. The ameliorative effect of quercetin on bisphenol A-induced toxicity in mitochondria isolated from rats. Environ. Sci. Pollut. Res. 2019, 26, 7688–7696. [Google Scholar] [CrossRef]
- Ravindranath, V.; Anandatheerthavarada, H.K. Preparation of brain microsomes with cytochrome P450 activity using calcium aggregation method. Anal. Biochem. 1990, 187, 310–313. [Google Scholar] [CrossRef]
- Cunha, M.P.; Lieberknecht, V.; Ramos-Hryb, A.B.; Olescowicz, G.; Ludka, F.K.; Tasca, C.I.; Gabilan, N.H.; Rodrigues, A.L. Creatine affords protection against glutamate-induced nitrosative and oxidative stress. Neurochem. Int. 2016, 95, 4–14. [Google Scholar] [CrossRef]
- Brewer, G.J.; Wallimann, T.W. Protective effect of the energy precursor creatine against toxicity of glutamate and beta-amyloid in rat hippocampal neurons. J. Neurochem. 2000, 74, 1968–1978. [Google Scholar] [CrossRef]
- Andreassen, O.A.; Dedeoglu, A.; Ferrante, R.J.; Jenkins, B.G.; Ferrante, K.L.; Thomas, M.; Friedlich, A.; Browne, S.E.; Schilling, G.; Borchelt, D.R.; et al. Creatine increase survival and delays motor symptoms in a transgenic animal model of Huntington’s disease. Neurobiol. Dis. 2001, 8, 479–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, M.P.; Machado, D.G.; Capra, J.C.; Jacinto, J.; Bettio, L.E.; Rodrigues, A.L. Antidepressant-like effect of creatine in mice involves dopaminergic activation. J. Psychopharmacol. 2012, 26, 1489–1501. [Google Scholar] [CrossRef] [PubMed]
- Gualano, B.; Roschel, H.; Lancha, A.H., Jr.; Brightbill, C.E.; Rawson, E.S. In sickness and in health: The widespread application of creatine supplementation. Amino Acids 2012, 43, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Percário, S.; Domingues, S.P.D.T.; Teixeira, L.F.M.; Vieira, J.L.F.; de Vasconcelos, F.; Ciarrocchi, D.M.; Almeida, E.D.; Conte, M. Effects of creatine supplementation on oxidative stress profile of athletes. J. Int. Soc. Sports Nutr. 2012, 9, 56. [Google Scholar] [CrossRef] [Green Version]
- Tirmenstein, M.A.; Hu, C.X.; Scicchitano, M.S.; Narayanan, P.K.; McFarland, D.C.; Thomas, H.C.; Schwartz, L.W. Effects of 6-hydroxydopamine on mitochondrial function and glutathione status in SH-SY5Y human neuroblastoma cells. Toxicol. In Vitro 2005, 19, 471–479. [Google Scholar] [CrossRef]
- Drahota, Z.; Křiváková, P.; Červinková, Z.; Kmoníčková, E.; Lotková, H.; Kučera, O.; Houštěk, J. Tert-butyl hydroperoxide selectively inhibits mitochondrial respiratory-chain enzymes in isolated rat hepatocytes. Physiol. Res. 2005, 54, 67–72. [Google Scholar] [CrossRef]
- Guidi, C.; Potenza, L.; Sestili, P.; Martinelli, C.; Guescini, M.; Stocchi, L.; Zeppa, S.; Polidori, E.; Annibalini, G.; Stocchi, V. Differential effect of creatine on oxidatively-injured mitochondrial and nuclear DNA. Biochim. Biophys. Acta 2008, 1780, 16–26. [Google Scholar] [CrossRef]
- Beaton, L.J.; Tarnopolsky, M.A.; Phillips, S.M. Contraction-induced muscle damage in humans following calcium channel blocker administration. J. Physiol. 2002, 544, 849–859. [Google Scholar] [CrossRef]
- Cooke, M.B.; Rybalka, E.; Williams, A.D.; Cribb, P.J.; Hayes, A. Creatine supplementation enhances muscle force recovery after eccentrically-induced muscle damage in healthy individuals. J. Int. Soc. Sports Nutr. 2009, 6, 13. [Google Scholar] [CrossRef]
- Korge, P.; Byrd, S.K.; Campbell, K.B. Functional coupling between sarcoplasmic-reticulum-bound creatine kinase and Ca(2+)-ATPase. Eur. J. Biochem. 1993, 213, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Valerio, A.; D’Antona, G.; Nisoli, E. Branched-chain amino acids, mitochondrial biogenesis, and healthspan: An evolutionary perspective. Aging 2011, 3, 464–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefani, G.P.; Nunes, R.B.; Dornelles, A.Z.; Alves, J.P.; Piva, M.O.; Domenico, M.D.; Rhoden, C.R.; Lago, P.D. Effects of creatine supplementation associated with resistance training on oxidative stress in different tissues of rats. J. Int. Soc. Sport Nutr. 2014, 11, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sestili, P.; Martinelli, C.; Bravi, G.; Piccoli, G.; Curci, R.; Battistelli, M.; Falcieri, E.; Agostini, D.; Gioacchini, A.M.; Stocchi, V. Creatine supplementation affords cytoprotection in oxidatively injured cultured mammalian cells via direct antioxidant activity. Free Radic. Biol. Med. 2006, 40, 837–849. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostadinova, I.; Kondeva-Burdina, M.; Marinov, L.; Vezenkov, L.L.; Simeonova, R. Newly Synthesized Creatine Derivatives as Potential Neuroprotective and Antioxidant Agents on In Vitro Models of Parkinson’s Disease. Life 2023, 13, 139. https://doi.org/10.3390/life13010139
Kostadinova I, Kondeva-Burdina M, Marinov L, Vezenkov LL, Simeonova R. Newly Synthesized Creatine Derivatives as Potential Neuroprotective and Antioxidant Agents on In Vitro Models of Parkinson’s Disease. Life. 2023; 13(1):139. https://doi.org/10.3390/life13010139
Chicago/Turabian StyleKostadinova, Ivanka, Magdalena Kondeva-Burdina, Lyubomir Marinov, Lubomir L. Vezenkov, and Rumyana Simeonova. 2023. "Newly Synthesized Creatine Derivatives as Potential Neuroprotective and Antioxidant Agents on In Vitro Models of Parkinson’s Disease" Life 13, no. 1: 139. https://doi.org/10.3390/life13010139