
Role of SaPCR2 in Zn Uptake in the Root Elongation Zone of the Zn/Cd Hyperaccumulator Sedum alfredii

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Culture Conditions
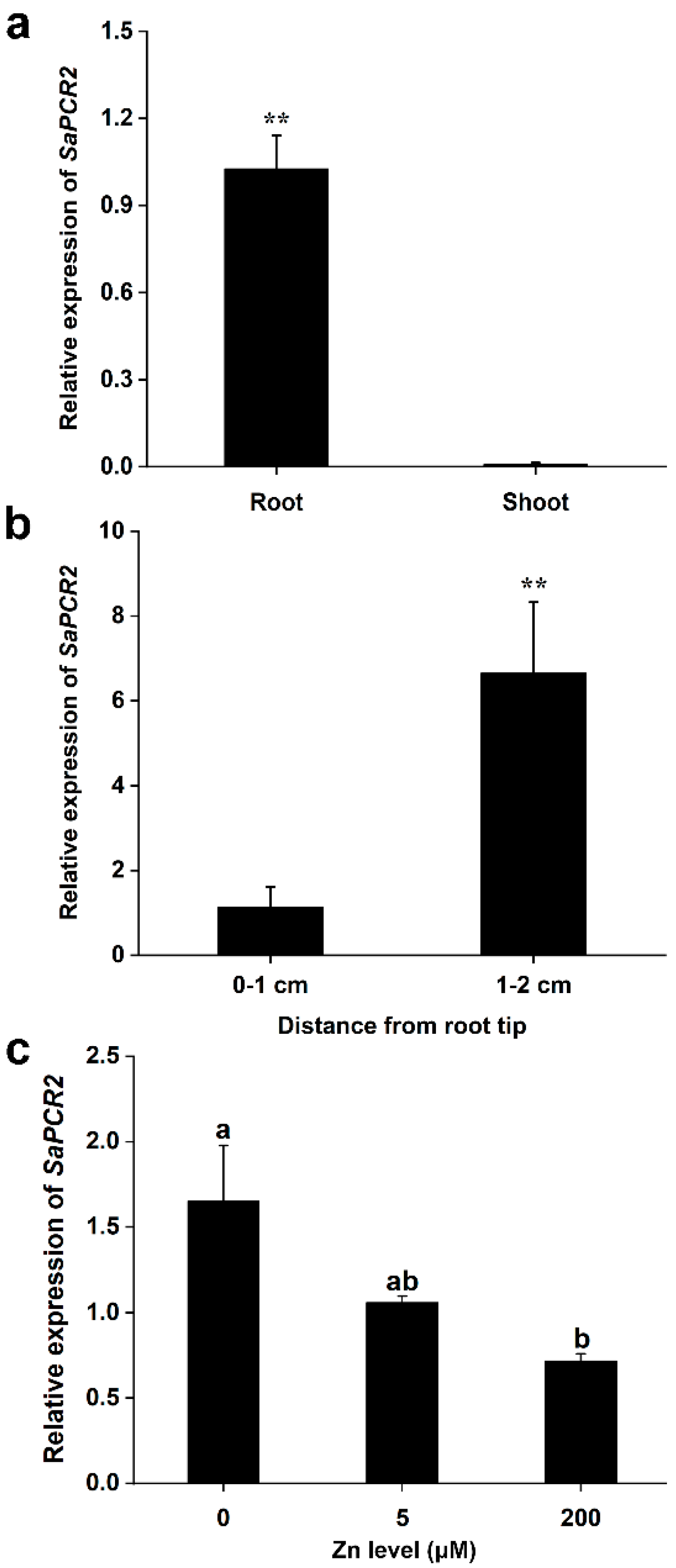
2.2. Expression Pattern of SaPCR2 in S. alfredii
2.3. Yeast Expression Analysis
2.4. Heavy Metal Concentration Analysis in the SaPCR2 Overexpression of NHE S. alfredii
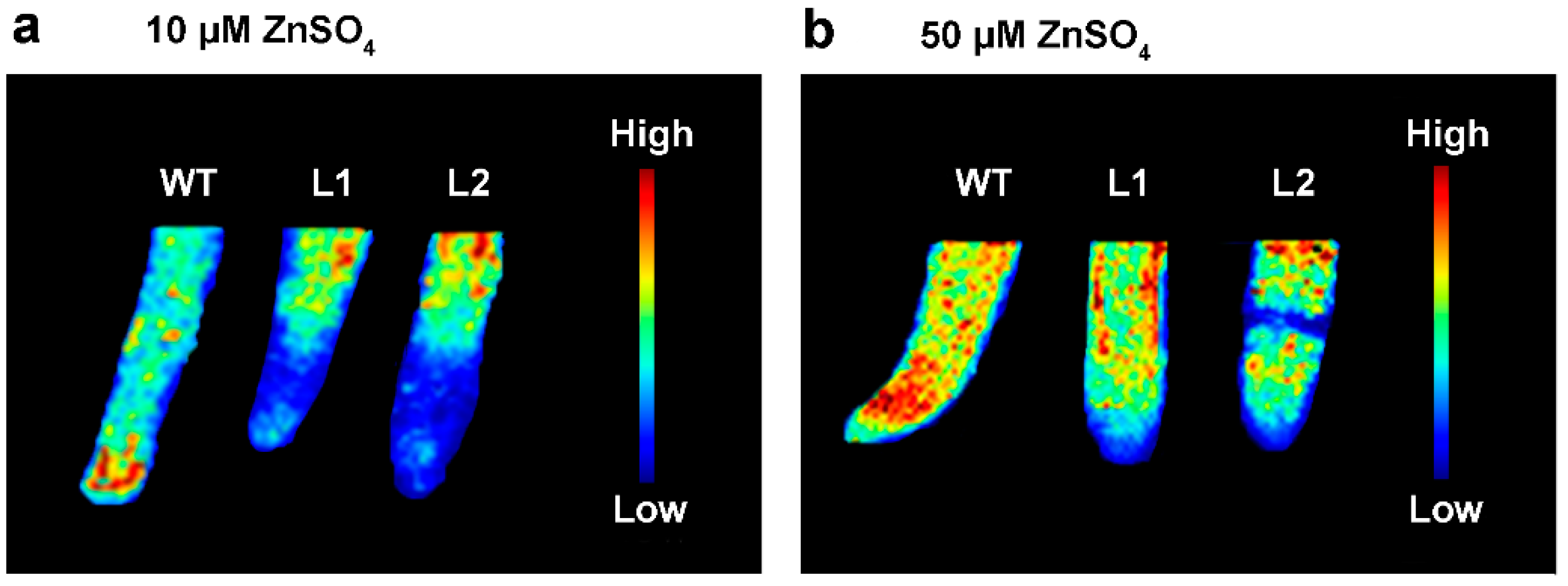
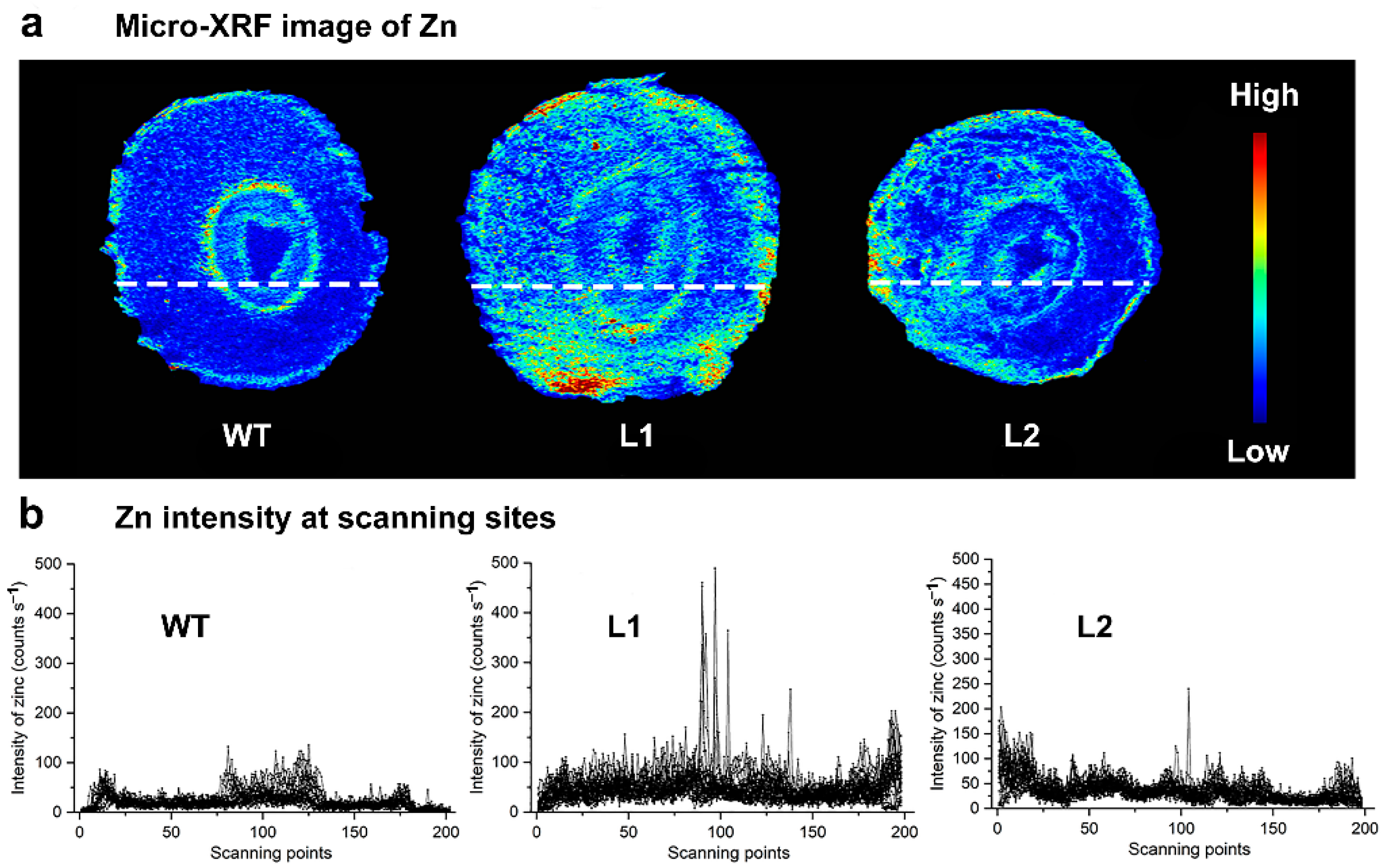
2.5. SR-μ-XRF Analysis Revealed the Zinc Distribution in the SaPCR2-Overexpressing NHE S. alfredii
2.6. Root Morphology in the SaPCR2-Overexpressing NHE S. alfredii
2.7. Statistical Analysis
3. Results
3.1. Expression Pattern of SaPCR2 in the HE and NHE S. alfredii
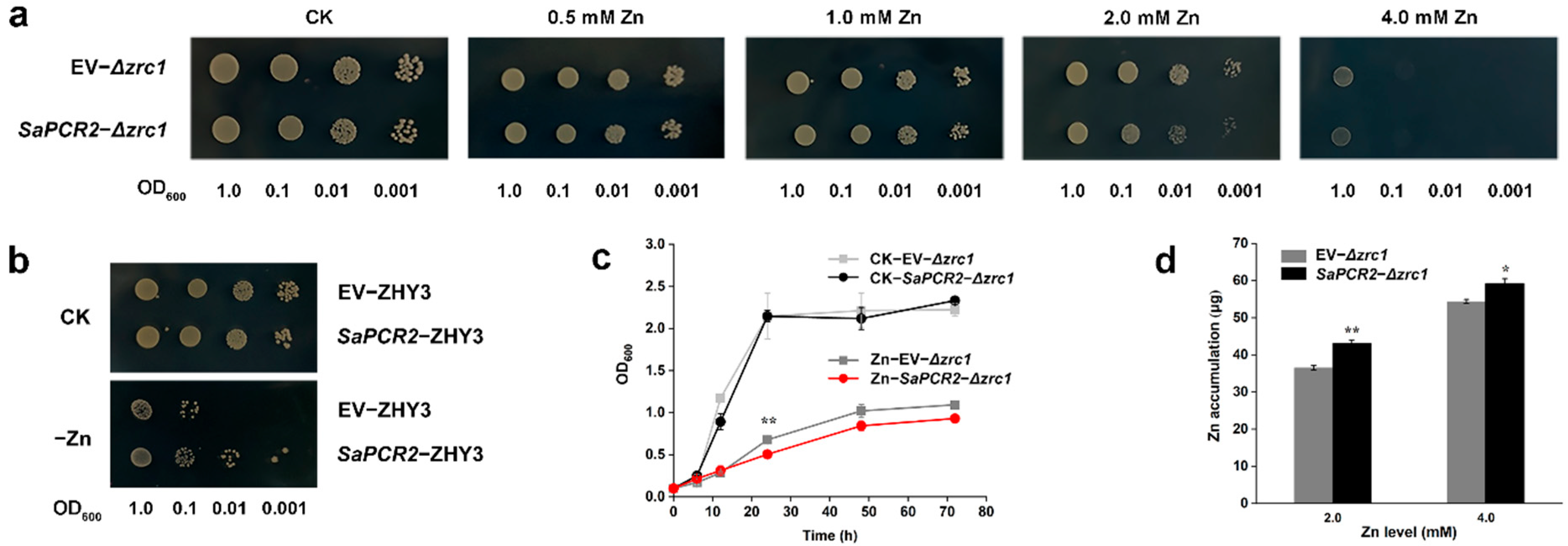
3.2. Functional Analysis of SaPCR2 in Yeast
3.3. Effect of SaPCR2 Overexpression on Zn Concentration in the NHE S. alfredii
3.4. Effect of SaPCR2 Overexpression on Zn Distribution in the NHE S. alfredii
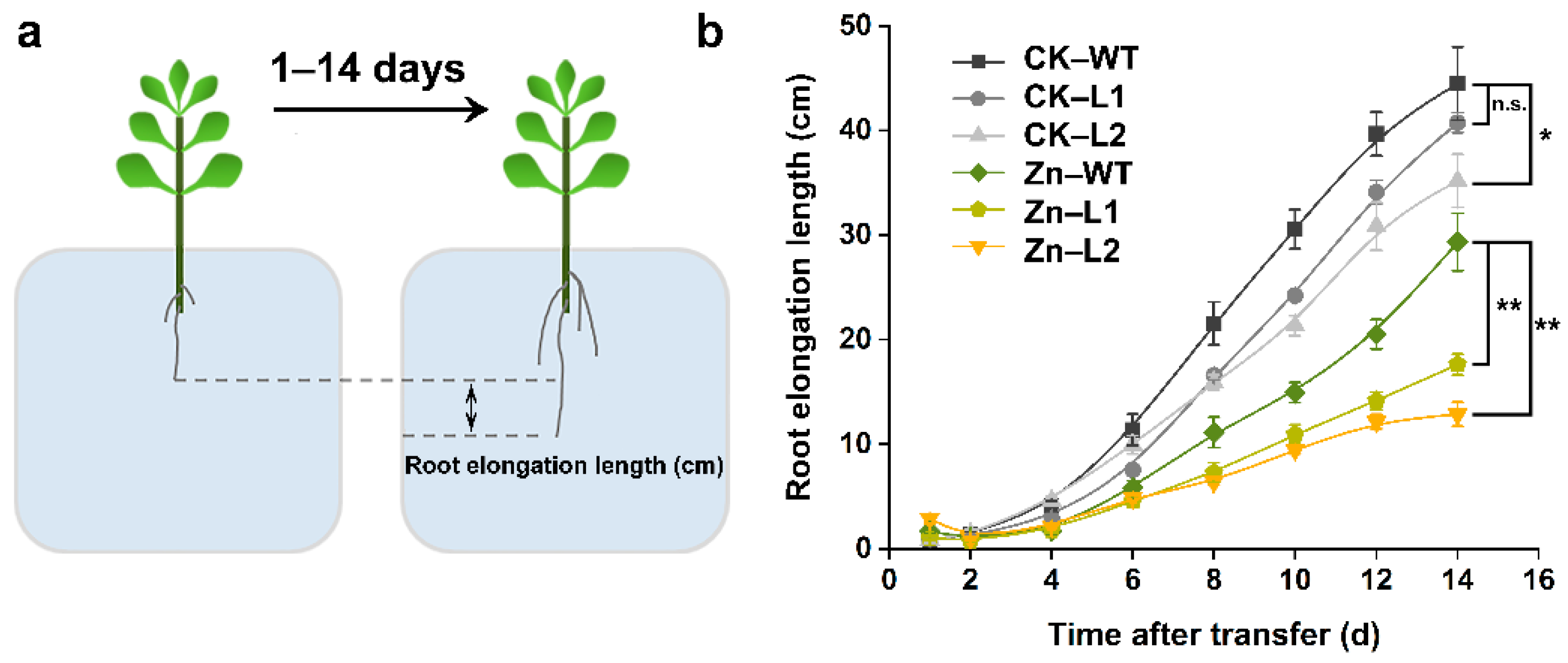
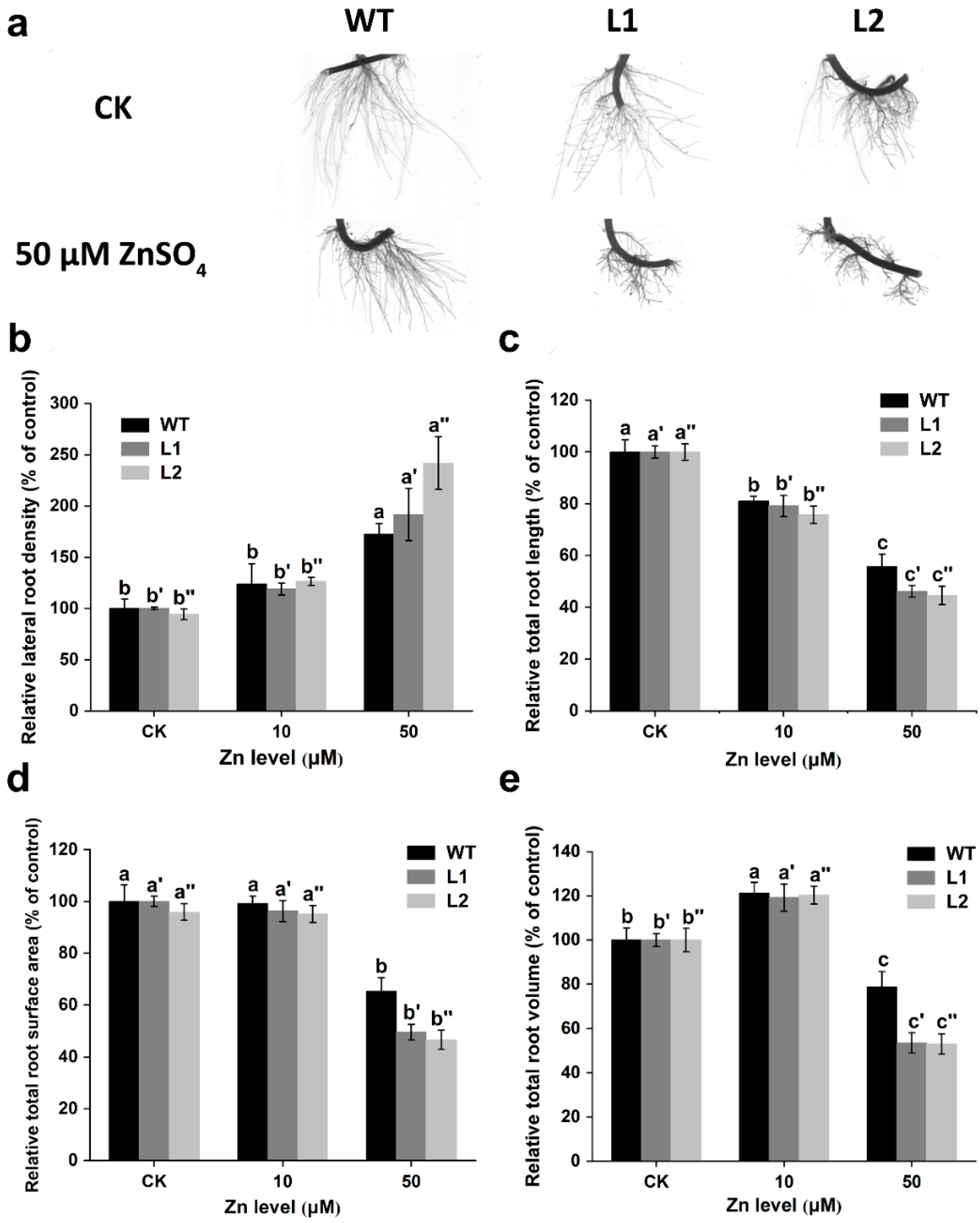
3.5. Root Morphology in the SaPCR2-Overexpressing NHE S. alfredii
4. Discussion
4.1. SaPCR2 Is Responsible for Zinc Uptake in S. alfredii
4.2. SaPCR2 Mediates Zinc Transport in the Root Elongation Zone of S. alfredii
4.3. The Overexpression of SaPCR2 Increased Zinc Concentration in the Shoots of S. alfredii
4.4. SaPCR2 Participates in Zn and Cd Transport, but May Not Participate in Mn, Cu or Fe Transport
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Duan, X.; Wang, L. Spatial distribution and source analysis of heavy metals in soils influenced by industrial enterprise distribution: Case study in Jiangsu Province. Sci. Total Environ. 2020, 710, 134953. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Zheng, N.; Tang, L.; Ji, X.; Li, Y.; Hua, X. Pollution characteristics, sources, and health risk assessment of human exposure to Cu, Zn, Cd and Pb pollution in urban street dust across China between 2009 and 2018. Environ. Int. 2019, 128, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Ma, Y.; Zhu, Y.; Tang, Z.; McGrath, S.P. Soil contamination in China: Current status and mitigation strategies. Environ. Sci. Technol. 2015, 49, 750–759. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—Iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Hansch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef]
- Lin, Y.; Aarts, M.G. The molecular mechanism of zinc and cadmium stress response in plants. Cell Mol. Life Sci. 2012, 69, 3187–3206. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, L.; Li, Y.; Li, H.; Wang, W.; Ye, B. Impacts of lead/zinc mining and smelting on the environment and human health in China. Environ. Monit. Assess. 2012, 184, 2261–2273. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals-Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Mahar, A.; Wang, P.; Ali, A.; Awasthi, M.K.; Lahori, A.H.; Wang, Q.; Li, R.; Zhang, Z. Challenges and opportunities in the phytoremediation of heavy metals contaminated soils: A review. Ecotoxicol. Environ. Saf. 2016, 126, 111–121. [Google Scholar] [CrossRef]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gurajala, H.; Wu, L.; van der Ent, A.; Qiu, R.; Baker, A.J.M.; Tang, Y.; Yang, X.; Shu, W. Hyperaccumulator Plants from China: A Synthesis of the Current State of Knowledge. Environ. Sci. Technol. 2018, 52, 11980–11994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, S.; Zeng, G.; Wu, H.; Zhang, C.; Dai, J.; Liang, J.; Yu, J.; Ren, X.; Yi, H.; Cheng, M.; et al. Biological technologies for the remediation of co-contaminated soil. Crit. Rev. Biotechnol. 2017, 37, 1062–1076. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Kang, H.; Zhang, X.; Shao, H.; Chu, L.; Ruan, C. A critical review on the bio-removal of hazardous heavy metals from contaminated soils: Issues, progress, eco-environmental concerns and opportunities. J. Hazard. Mater. 2010, 174, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yang, X.; He, Z.; Yang, J. Root Morphology and Zn2+ Uptake Kinetics of the Zn Hyperaccumulator of Sedum alfredii Hance. J. Integr. Plant Biol. 2005, 47, 927–934. [Google Scholar] [CrossRef]
- Lu, L.; Tian, S.; Yang, X.; Wang, X.; Brown, P.; Li, T.; He, Z. Enhanced root-to-shoot translocation of cadmium in the hyperaccumulating ecotype of Sedum alfredii. J. Exp. Bot. 2008, 59, 3203–3213. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, U. Metal Hyperaccumulation in Plants. Annu. Rev. Plant Biol. 2010, 61, 517–534. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C.; Schat, H. Molecular mechanisms of metal hyperaccumulation in plants. New Phytol. 2009, 181, 759–776. [Google Scholar] [CrossRef]
- Song, W.; Martinoia, E.; Lee, J.; Kim, D.; Kim, D.; Vogt, E.; Shim, D.; Choi, K.; Hwang, I.; Lee, Y. A novel family of cys-rich membrane proteins mediates cadmium resistance in Arabidopsis. Plant Physiol. 2004, 135, 1027–1039. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Choi, K.; Kim, D.; Geisler, M.; Park, J.; Vincenzetti, V.; Schellenberg, M.; Kim, S.; Lim, Y.; Noh, E.; et al. Arabidopsis PCR2 Is a Zinc Exporter Involved in Both Zinc Extrusion and Long-Distance Zinc Transport. Plant Cell 2010, 22, 2237–2252. [Google Scholar] [CrossRef] [Green Version]
- Frary, A.; Nesbitt, T.C.; Frary, A.; Grandillo, S.; van der Knaap, E.; Cong, B.; Liu, J.P.; Meller, J.; Elber, R.; Alpert, K.B.; et al. fw2.2: A quantitative trait locus key to the evolution of tomato fruit size. Science 2000, 289, 85–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.; Rupe, M.A.; Dieter, J.A.; Zou, J.; Spielbauer, D.; Duncan, K.E.; Howard, R.J.; Hou, Z.; Simmons, C.R. Cell Number Regulator1 Affects Plant and Organ Size in Maize: Implications for Crop Yield Enhancement and Heterosis. Plant Cell 2010, 22, 1057–1073. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Shohag, M.J.I.; Feng, Y.; He, Z.; Yang, X. Transcriptome Comparison Reveals the Adaptive Evolution of Two Contrasting Ecotypes of Zn/Cd Hyperaccumulator Sedum alfredii Hance. Front. Plant Sci. 2017, 8, 425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, Y.; Zhang, M.; Feng, Y.; Yang, X.; Islam, E. cDNA-AFLP analysis of inducible gene expression in zinc hyperaccumulator Sedum alfredii Hance under zinc induction. Environ. Exp. Bot. 2010, 68, 107–112. [Google Scholar] [CrossRef]
- Yang, Q.; Ma, X.; Luo, S.; Gao, J.; Yang, X.; Feng, Y. SaZIP4, an uptake transporter of Zn/Cd hyperaccumulator Sedum alfredii Hance. Environ. Exp. Bot. 2018, 155, 107–117. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Song, H.; Zhao, J.; Shabala, S.; Tian, S.; Yang, X. A novel plasma membrane-based NRAMP transporter contributes to Cd and Zn hyperaccumulation in Sedum alfredii Hance. Environ. Exp. Bot. 2020, 176, 104121. [Google Scholar] [CrossRef]
- Zhang, M.; Senoura, T.; Yang, X.; Nishizawa, N.K. Functional analysis of metal tolerance proteins isolated from Zn/Cd hyperaccumulating ecotype and non-hyperaccumulating ecotype of Sedum alfredii Hance. FEBS Lett. 2011, 585, 2604–2609. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Gao, X.; Zhao, J.; Zhang, J.; Chen, S.; Lu, L. Plant Cadmium Resistance 2 (SaPCR2) facilitates cadmium efflux in the roots of hyperaccumulator Sedum alfredii Hance. Front. Plant Sci. 2020, 11, 568887. [Google Scholar] [CrossRef]
- Yang, X.; Long, X.; Ye, H.; He, Z.; Calvert, D.V.; Stoffella, P.J. Cadmium tolerance and hyperaccumulation in a new Zn-hyperaccumulating plant species (Sedum alfredii Hance). Plant Soil 2004, 259, 181–189. [Google Scholar] [CrossRef]
- Yang, X.; Long, X.; Ni, W.; Fu, C. Sedum alfredii H: A new Zn hyperaccumulating plant first found in China. Chin. Sci. Bull. 2002, 47, 1634–1637. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Webb, S.M. The MicroAnalysis Toolkit: X-ray Fluorescence Image Processing Software. In Proceedings of the 10th International Conference on X-ray Microscopy, Univ Chicago, Chicago, IL, USA, 15–20 August 2010; pp. 196–199. [Google Scholar]
- Hacisalihoglu, G.; Hart, J.J.; Kochian, L.V. High- and low-affinity zinc transport systems and their possible role in zinc efficiency in bread wheat. Plant Physiol. 2001, 125, 456–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Q.; Jupa, R.; Luo, J.; Lux, A.; Kovac, J.; Wen, Y.; Zhou, Y.; Jan, J.; Liang, Y.; Li, T. The apoplasmic pathway via the root apex and lateral roots contributes to Cd hyperaccumulation in the hyperaccumulator Sedum alfredii. J. Exp. Bot. 2017, 68, 739–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Liao, X.; Labavitch, J.; Yang, X.; Nelson, E.; Du, Y.; Brown, P.H.; Tian, S. Speciation and localization of Zn in the hyperaccumulator Sedum alfredii by extended X-ray absorption fine structure and micro-X-ray fluorescence. Plant Physiol. Biochem. 2014, 84, 224–232. [Google Scholar] [CrossRef] [Green Version]
- Milner, M.J.; Craft, E.; Yamaji, N.; Koyama, E.; Ma, J.; Kochian, L.V. Characterization of the high affinity Zn transporter from Noccaea caerulescens, NcZNT1, and dissection of its promoter for its role in Zn uptake and hyperaccumulation. New Phytol. 2012, 195, 113–123. [Google Scholar] [CrossRef]
- Pence, N.S.; Larsen, P.B.; Ebbs, S.D.; Letham, D.L.; Lasat, M.M.; Garvin, D.F.; Eide, D.; Kochian, L.V. The molecular physiology of heavy metal transport in the Zn/Cd hyperaccumulator Thlaspi caerulescens. Proc. Natl. Acad. Sci. USA 2000, 97, 4956–4960. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef] [Green Version]
- Milner, M.J.; Seamon, J.; Craft, E.; Kochian, L.V. Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J. Exp. Bot. 2013, 64, 369–381. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Lee, H.; Jin, S.; Ko, D.; Martinoia, E.; Lee, Y.; An, G.; Ahn, S. Rice PCR1 influences grain weight and Zn accumulation in grains. Plant Cell Environ. 2015, 38, 2327–2339. [Google Scholar] [CrossRef]
- Wang, P.; Li, Z.; Wei, J.; Zhao, Z.; Sun, D.; Cui, S. A Na+/Ca2+ exchanger-like protein (AtNCL) involved in salt stress in Arabidopsis. J. Biol. Chem. 2012, 287, 44062–44070. [Google Scholar] [CrossRef] [Green Version]
- Krulwich, T.A.; Hicks, D.B.; Ito, M. Cation/proton antiporter complements of bacteria: Why so large and diverse? Mol. Microbiol. 2009, 74, 257–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padan, E.; Venturib, M.; Gerchmana, Y.; Dovera, N. Na+/H+ antiporters. Biochim. Biophys. Acta Bioenerg. 2001, 1505, 144–157. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.; Xie, R.; Wang, H.; Hu, Y.; Ge, J.; Liao, X.; Gao, X.; Brown, P.; Lin, X.; Lu, L. Calcium Deficiency Triggers Phloem Remobilization of Cadmium in a Hyperaccumulating Species. Plant Physiol. 2016, 172, 2300–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, S.; Lu, L.; Zhang, J.; Wang, K.; Brown, P.; He, Z.; Liang, J.; Yang, X. Calcium protects roots of Sedum alfredii H. against cadmium-induced oxidative stress. Chemosphere 2011, 84, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Lu, L.; Labavitch, J.; Yang, X.; He, Z.; Hu, H.; Sarangi, R.; Newville, M.; Commisso, J.; Brown, P. Cellular Sequestration of Cadmium in the Hyperaccumulator Plant Species Sedum alfredii. Plant Physiol. 2011, 157, 1914–1925. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Tian, S.; Zhang, M.; Zhang, J.; Yang, X.; Jiang, H. The role of Ca pathway in Cd uptake and translocation by the hyperaccumulator Sedum alfredii. J. Hazard. Mater. 2010, 183, 22–28. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, J.; Lin, J.; Wu, Z.; Xu, K.; Tao, J.; Lin, H.; Tian, S.; Lu, L. Role of SaPCR2 in Zn Uptake in the Root Elongation Zone of the Zn/Cd Hyperaccumulator Sedum alfredii. Life 2022, 12, 768. https://doi.org/10.3390/life12050768
Ge J, Lin J, Wu Z, Xu K, Tao J, Lin H, Tian S, Lu L. Role of SaPCR2 in Zn Uptake in the Root Elongation Zone of the Zn/Cd Hyperaccumulator Sedum alfredii. Life. 2022; 12(5):768. https://doi.org/10.3390/life12050768
Chicago/Turabian StyleGe, Jun, Jiayu Lin, Zhiying Wu, Kuan Xu, Jingyu Tao, Haizhong Lin, Shengke Tian, and Lingli Lu. 2022. "Role of SaPCR2 in Zn Uptake in the Root Elongation Zone of the Zn/Cd Hyperaccumulator Sedum alfredii" Life 12, no. 5: 768. https://doi.org/10.3390/life12050768