
Role of Uncoupling Protein 2 Gene Polymorphisms on the Risk of Ischemic Stroke in a Sardinian Population

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Ethical Approval
4.3. DNA Isolation and Genotyping
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sacco, R.L. Risk factors and outcomes for ischemic stroke. Neurology 1995, 45, S10–S14. [Google Scholar]
- Allen, C.L.; Bayraktutan, U. Risk factors for ischaemic stroke. Int. J. Stroke 2008, 3, 105–116. [Google Scholar] [CrossRef]
- Boehme, A.K.; Esenwa, C.; Elkind, M.S. Stroke Risk Factors, Genetics, and Prevention. Circ. Res. 2017, 120, 472–495. [Google Scholar] [CrossRef]
- Lan, C.; Chen, X.; Zhang, Y.; Wang, W.; Wang, W.E.; Liu, Y.; Cai, Y.; Ren, H.; Zheng, S.; Zhou, L.; et al. Curcumin prevents strokes in stroke-prone spontaneously hypertensive rats by improving vascular endothelial function. BMC Cardiovasc. Disord. 2018, 18, 43. [Google Scholar] [CrossRef] [Green Version]
- Rubattu, S.; Stanzione, R.; Bianchi, F.; Cotugno, M.; Forte, M.; Della Ragione, F.; Fioriniello, S.; D’Esposito, M.; Marchitti, S.; Madonna, M.; et al. Reduced brain UCP2 expression mediated by microRNA-503 contributes to increased stroke susceptibility in the high-salt fed stroke-prone spontaneously hypertensive rat. Cell Death Dis. 2017, 8, e2891. [Google Scholar] [CrossRef]
- Rubattu, S.; Di Castro, S.; Cotugno, M.; Bianchi, F.; Mattioli, R.; Baima, S.; Stanzione, R.; Madonna, M.; Bozzao, C.; Marchitti, S.; et al. Protective effects of Brassica oleracea sprouts extract toward renal damage in high-salt-fed SHRSP: Role of AMPK/PPARalpha/UCP2 axis. J. Hypertens 2015, 33, 1465–1479. [Google Scholar] [CrossRef]
- Rousset, S.; Alves-Guerra, M.C.; Mozo, J.; Miroux, B.; Cassard-Doulcier, A.M.; Bouillaud, F.; Ricquier, D. The biology of mitochondrial uncoupling proteins. Diabetes 2004, 53 (Suppl. 1), S130–S135. [Google Scholar] [CrossRef] [Green Version]
- Brand, M.D.; Esteves, T.C. Physiological functions of the mitochondrial uncoupling proteins UCP2 and UCP3. Cell Metab. 2005, 2, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.Y.; Baffy, G.; Perret, P.; Krauss, S.; Peroni, O.; Grujic, D.; Hagen, T.; Vidal-Puig, A.J.; Boss, O.; Kim, Y.B.; et al. Uncoupling protein-2 negatively regulates insulin secretion and is a major link between obesity, beta cell dysfunction, and type 2 diabetes. Cell 2001, 105, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Urbano, F.; Filippello, A.; Di Pino, A.; Barbagallo, D.; Di Mauro, S.; Pappalardo, A.; Rabuazzo, A.M.; Purrello, M.; Purrello, F.; Piro, S. Altered expression of uncoupling protein 2 in GLP-1-producing cells after chronic high glucose exposure: Implications for the pathogenesis of diabetes mellitus. Am. J. Physiol. Cell Physiol. 2016, 310, C558–C567. [Google Scholar] [CrossRef] [Green Version]
- Murakami, K.; Mizushige, K.; Noma, T.; Tsuji, T.; Kimura, S.; Kohno, M. Perindopril effect on uncoupling protein and energy metabolism in failing rat hearts. Hypertension 2002, 40, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Esfandiary, A.; Kutsche, H.S.; Schreckenberg, R.; Weber, M.; Pak, O.; Kojonazarov, B.; Sydykov, A.; Hirschhauser, C.; Wolf, A.; Haag, D.; et al. Protection against pressure overload-induced right heart failure by uncoupling protein 2 silencing. Cardiovasc. Res. 2019, 115, 1217–1227. [Google Scholar] [CrossRef]
- Tian, X.Y.; Ma, S.; Tse, G.; Wong, W.T.; Huang, Y. Uncoupling Protein 2 in Cardiovascular Health and Disease. Front. Physiol. 2018, 9, 1060. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Park, E.; Lee, J.; Lee, Y.; Rooney, B.V.; Park, Y. Exercise training mitigates ER stress and UCP2 deficiency-associated coronary vascular dysfunction in atherosclerosis. Sci. Rep. 2021, 11, 15449. [Google Scholar] [CrossRef]
- Forte, M.; Bianchi, F.; Cotugno, M.; Marchitti, S.; Stanzione, R.; Maglione, V.; Sciarretta, S.; Valenti, V.; Carnevale, R.; Versaci, F.; et al. An interplay between UCP2 and ROS protects cells from high-salt-induced injury through autophagy stimulation. Cell Death Dis. 2021, 12, 919. [Google Scholar] [CrossRef]
- Stanzione, R.; Forte, M.; Cotugno, M.; Bianchi, F.; Marchitti, S.; Busceti, C.L.; Fornai, F.; Rubattu, S. Uncoupling Protein 2 as a pathogenic determinant and therapeutic target in cardiovascular and metabolic diseases. Curr. Neuropharmacol. 2021, 20, 662–674. [Google Scholar] [CrossRef]
- Busceti, C.L.; Cotugno, M.; Bianchi, F.; Forte, M.; Stanzione, R.; Marchitti, S.; Battaglia, G.; Nicoletti, F.; Fornai, F.; Rubattu, S. Brain Overexpression of Uncoupling Protein-2 (UCP2) Delays Renal Damage and Stroke Occurrence in Stroke-Prone Spontaneously Hypertensive Rats. Int. J. Mol. Sci. 2020, 21, 4289. [Google Scholar] [CrossRef]
- Di Castro, S.; Scarpino, S.; Marchitti, S.; Bianchi, F.; Stanzione, R.; Cotugno, M.; Sironi, L.; Gelosa, P.; Duranti, E.; Ruco, L.; et al. Differential modulation of uncoupling protein 2 in kidneys of stroke-prone spontaneously hypertensive rats under high-salt/low-potassium diet. Hypertension 2013, 61, 534–541. [Google Scholar] [CrossRef] [Green Version]
- Qian, L.; Xu, K.; Xu, X.; Gu, R.; Liu, X.; Shan, S.; Yang, T. UCP2 −866G/A, Ala55Val and UCP3 -55C/T polymorphisms in association with obesity susceptibility—A meta-analysis study. PLoS ONE 2013, 8, e58939. [Google Scholar] [CrossRef] [Green Version]
- Dalgaard, L.T. Genetic Variance in Uncoupling Protein 2 in Relation to Obesity, Type 2 Diabetes, and Related Metabolic Traits: Focus on the Functional -866G>A Promoter Variant (rs659366). J. Obes. 2011, 2011, 340241. [Google Scholar] [CrossRef] [Green Version]
- Donadelli, M.; Dando, I.; Fiorini, C.; Palmieri, M. UCP2, a mitochondrial protein regulated at multiple levels. Cell. Mol. Life Sci. 2014, 71, 1171–1190. [Google Scholar] [CrossRef]
- Li, J.; Jiang, R.; Cong, X.; Zhao, Y. UCP2 gene polymorphisms in obesity and diabetes, and the role of UCP2 in cancer. FEBS Lett. 2019, 593, 2525–2534. [Google Scholar] [CrossRef] [Green Version]
- Surniyantoro, H.N.E.; Sadewa, A.H.; Hastuti, P. Uncoupling Protein 2 (UCP2) as Genetic Risk Factor for Obesity in Indonesia is Different in Gender Stratification. Kobe J. Med. Sci. 2018, 64, E64–E72. [Google Scholar]
- Esterbauer, H.; Schneitler, C.; Oberkofler, H.; Ebenbichler, C.; Paulweber, B.; Sandhofer, F.; Ladurner, G.; Hell, E.; Strosberg, A.D.; Patsch, J.R.; et al. A common polymorphism in the promoter of UCP2 is associated with decreased risk of obesity in middle-aged humans. Nat. Genet. 2001, 28, 178–183. [Google Scholar] [CrossRef]
- Jia, J.J.; Zhang, X.; Ge, C.R.; Jois, M. The polymorphisms of UCP2 and UCP3 genes associated with fat metabolism, obesity and diabetes. Obes. Rev. 2009, 10, 519–526. [Google Scholar] [CrossRef]
- Wang, H.; Chu, W.S.; Lu, T.; Hasstedt, S.J.; Kern, P.A.; Elbein, S.C. Uncoupling protein-2 polymorphisms in type 2 diabetes, obesity, and insulin secretion. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E1–E7. [Google Scholar] [CrossRef]
- Chai, Y.; Gu, B.; Qiu, J.R.; Yi, H.G.; Zhu, Q.; Zhang, L.; Hu, G. The uncoupling protein 2−866G > a polymorphism is associated with the risk of ischemic stroke in Chinese type 2 diabetic patients. CNS Neurosci. Ther. 2012, 18, 636–640. [Google Scholar] [CrossRef]
- Chai, Y.; Gu, B.; Qiu, J.R.; Yi, H.G.; Zhu, Q.; Zhang, L.; Hu, G. Effects of uncoupling protein 2−866G/A polymorphism on platelet reactivity and prognosis in Chinese patients with type 2 diabetes and ischemic stroke. Int. J. Neurosci. 2013, 123, 752–758. [Google Scholar] [CrossRef]
- Diaz-Maroto Cicuendez, I.; Fernandez-Diaz, E.; Garcia-Garcia, J.; Jordan, J.; Fernandez-Cadenas, I.; Montaner, J.; Serrano-Heras, G.; Segura, T. The UCP2−866G/A Polymorphism Could be Considered as a Genetic Marker of Different Functional Prognosis in Ischemic Stroke After Recanalization. Neuromolecular. Med. 2017, 19, 571–578. [Google Scholar] [CrossRef]
- Zee, R.Y.; Ridker, P.M.; Chasman, D.I. Genetic variants in eleven telomere-associated genes and the risk of incident cardio/cerebrovascular disease: The Women’s Genome Health Study. Clin. Chim. Acta 2011, 412, 199–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gioli-Pereira, L.; Santos, P.C.; Sugaya, L.S.; Ferreira, N.E.; Krieger, J.E.; Pereira, A.C.; Hueb, W.A. Association between UCP2 A55V polymorphism and risk of cardiovascular events in patients with multi-vessel coronary arterial disease. BMC Med. Genet. 2013, 14, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phulukdaree, A.; Moodley, D.; Khan, S.; Chuturgoon, A.A. Uncoupling protein 2−866G/A and uncoupling protein 3-55C/T polymorphisms in young South African Indian coronary artery disease patients. Gene 2013, 524, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.C.; Yang, L.; Liu, Y.Y.; Qin, Y.; Li, Y.P.; Zhang, L.; Yang, K.; Yang, Y. Polymorphisms in the Uncoupling Protein 2 Gene Are Associated with Diabetic Retinopathy in Han Chinese Patients with Type 2 Diabetes. Genet. Test. Mol. Biomark. 2018, 22, 637–643. [Google Scholar] [CrossRef]
- Dhamrait, S.S.; Stephens, J.W.; Cooper, J.A.; Acharya, J.; Mani, A.R.; Moore, K.; Miller, G.J.; Humphries, S.E.; Hurel, S.J.; Montgomery, H.E. Cardiovascular risk in healthy men and markers of oxidative stress in diabetic men are associated with common variation in the gene for uncoupling protein 2. Eur. Heart J. 2004, 25, 468–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleury, C.; Neverova, M.; Collins, S.; Raimbault, S.; Champigny, O.; Levi-Meyrueis, C.; Bouillaud, F.; Seldin, M.F.; Surwit, R.S.; Ricquier, D.; et al. Uncoupling protein-2: A novel gene linked to obesity and hyperinsulinemia. Nat. Genet. 1997, 15, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Arsenijevic, D.; Onuma, H.; Pecqueur, C.; Raimbault, S.; Manning, B.S.; Miroux, B.; Couplan, E.; Alves-Guerra, M.C.; Goubern, M.; Surwit, R.; et al. Disruption of the uncoupling protein-2 gene in mice reveals a role in immunity and reactive oxygen species production. Nat. Genet. 2000, 26, 435–439. [Google Scholar] [CrossRef]
- Krempler, F.; Esterbauer, H.; Weitgasser, R.; Ebenbichler, C.; Patsch, J.R.; Miller, K.; Xie, M.; Linnemayr, V.; Oberkofler, H.; Patsch, W. A functional polymorphism in the promoter of UCP2 enhances obesity risk but reduces type 2 diabetes risk in obese middle-aged humans. Diabetes 2002, 51, 3331–3335. [Google Scholar] [CrossRef]
- Cheurfa, N.; Dubois-Laforgue, D.; Ferrarezi, D.A.; Reis, A.F.; Brenner, G.M.; Bouche, C.; Le Feuvre, C.; Fumeron, F.; Timsit, J.; Marre, M.; et al. The common -866G>A variant in the promoter of UCP2 is associated with decreased risk of coronary artery disease in type 2 diabetic men. Diabetes 2008, 57, 1063–1068. [Google Scholar] [CrossRef]
- Fares, R.; Petta, S.; Lombardi, R.; Grimaudo, S.; Dongiovanni, P.; Pipitone, R.; Rametta, R.; Fracanzani, A.L.; Mozzi, E.; Craxi, A.; et al. The UCP2 -866 G>A promoter region polymorphism is associated with nonalcoholic steatohepatitis. Liver Int. 2015, 35, 1574–1580. [Google Scholar] [CrossRef]
- Xu, K.; Zhang, M.; Cui, D.; Fu, Y.; Qian, L.; Gu, R.; Wang, M.; Shen, C.; Yu, R.; Yang, T. UCP2−866G/A and Ala55Val, and UCP3-55C/T polymorphisms in association with type 2 diabetes susceptibility: A meta-analysis study. Diabetologia 2011, 54, 2315–2324. [Google Scholar] [CrossRef] [Green Version]
- Bielinski, S.J.; Pankow, J.S.; Boerwinkle, E.; Bray, M.S.; Kao, W.H.; Folsom, A.R. Lack of association between uncoupling protein-2 Ala55Val polymorphism and incident diabetes in the atherosclerosis risk in communities study. Acta Diabetol. 2008, 45, 179–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosmond, R.; Bouchard, C.; Bjorntorp, P. Lack of association between the uncoupling protein-2 Ala55Val gene polymorphism and phenotypic features of the Metabolic Syndrome. Biochim. Biophys. Acta 2002, 1588, 103–105. [Google Scholar] [CrossRef] [Green Version]
- Labayen, I.; Ortega, F.B.; Sjostrom, M.; Nilsson, T.K.; Olsson, L.A.; Ruiz, J.R. Association of common variants of UCP2 gene with low-grade inflammation in Swedish children and adolescents; the European Youth Heart Study. Pediatr. Res. 2009, 66, 350–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashemi, M.; Rezaei, H.; Kaykhaei, M.A.; Taheri, M. A 45-bp insertion/deletion polymorphism of UCP2 gene is associated with metabolic syndrome. J. Diabetes Metab. Disord. 2014, 13, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Say, Y.H. The association of insertions/deletions (INDELs) and variable number tandem repeats (VNTRs) with obesity and its related traits and complications. J. Physiol. Anthropol. 2017, 36, 25. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, G.; Sharma, R.K.; Baburaj, V.P.; Sankhwar, S.N.; Jafar, T.; Agrawal, S. Genetic risk factors for renal failure among north Indian ESRD patients. Clin. Biochem. 2008, 41, 525–531. [Google Scholar] [CrossRef]
- Heil, S.G.; Vermeulen, S.H.; Van der Rijt-Pisa, B.J.; den Heijer, M.; Blom, H.J. Role for mitochondrial uncoupling protein-2 (UCP2) in hyperhomocysteinemia and venous thrombosis risk? Clin. Chem. Lab. Med. 2008, 46, 655–659. [Google Scholar] [CrossRef]
- Mattiasson, G.; Shamloo, M.; Gido, G.; Mathi, K.; Tomasevic, G.; Yi, S.; Warden, C.H.; Castilho, R.F.; Melcher, T.; Gonzalez-Zulueta, M.; et al. Uncoupling protein-2 prevents neuronal death and diminishes brain dysfunction after stroke and brain trauma. Nat. Med. 2003, 9, 1062–1068. [Google Scholar] [CrossRef]
- Biino, G.; Casula, L.; de Terlizzi, F.; Adamo, M.; Vaccargiu, S.; Francavilla, M.; Loi, D.; Casti, A.; Atzori, M.; Cosso, M.; et al. Genetic architecture of hand quantitative ultrasound measures: A population-based study in a Sardinian genetic isolate. Bone 2010, 46, 1197–1203. [Google Scholar] [CrossRef]
- Chiang, C.W.K.; Marcus, J.H.; Sidore, C.; Biddanda, A.; Al-Asadi, H.; Zoledziewska, M.; Pitzalis, M.; Busonero, F.; Maschio, A.; Pistis, G.; et al. Genomic history of the Sardinian population. Nat. Genet. 2018, 50, 1426–1434. [Google Scholar] [CrossRef]
- Rubattu, S.; Stanzione, R.; Di Angelantonio, E.; Zanda, B.; Evangelista, A.; Tarasi, D.; Gigante, B.; Pirisi, A.; Brunetti, E.; Volpe, M. Atrial natriuretic peptide gene polymorphisms and risk of ischemic stroke in humans. Stroke 2004, 35, 814–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubattu, S.; Di Angelantonio, E.; Stanzione, R.; Zanda, B.; Evangelista, A.; Pirisi, A.; De Paolis, P.; Cota, L.; Brunetti, E.; Volpe, M. Gene polymorphisms of the renin-angiotensin-aldosterone system and the risk of ischemic stroke: A role of the A1166C/AT1 gene variant. J. Hypertens. 2004, 22, 2129–2134. [Google Scholar] [CrossRef] [PubMed]
- Rubattu, S.; Di Angelantonio, E.; Nitsch, D.; Gigante, B.; Zanda, B.; Stanzione, R.; Evangelista, A.; Pirisi, A.; Rosati, G.; Volpe, M. Polymorphisms in prothrombotic genes and their impact on ischemic stroke in a Sardinian population. Thromb. Haemost. 2005, 93, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Quarta, G.; Stanzione, R.; Evangelista, A.; Zanda, B.; Sciarretta, S.; Di Angelantonio, E.; Marchitti, S.; Di Murro, D.; Volpe, M.; Rubattu, S. A protective role of a cholesteryl ester transfer protein gene variant towards ischaemic stroke in Sardinians. J. Intern. Med. 2007, 262, 555–561. [Google Scholar] [CrossRef]
- Stanzione, R.; Di Angelantonio, E.; Evangelista, A.; Barbato, D.; Marchitti, S.; Zanda, B.; Pirisi, A.; Quarta, G.; Volpe, M.; Rubattu, S. Beta2-adrenergic receptor gene polymorphisms and risk of ischemic stroke. Am. J. Hypertens. 2007, 20, 657–662. [Google Scholar] [CrossRef] [Green Version]
- Lanni, F.; Santulli, G.; Izzo, R.; Rubattu, S.; Zanda, B.; Volpe, M.; Iaccarino, G.; Trimarco, B. The Pl(A1/A2) polymorphism of glycoprotein IIIa and cerebrovascular events in hypertension: Increased risk of ischemic stroke in high-risk patients. J. Hypertens. 2007, 25, 551–556. [Google Scholar] [CrossRef]
- Quarta, G.; Stanzione, R.; Evangelista, A.; Zanda, B.; Di Angelantonio, E.; Marchitti, S.; Di Castro, S.; Di Vavo, M.; Volpe, M.; Rubattu, S. Phosphodiesterase 4D and 5-lipoxygenase activating protein genes and risk of ischemic stroke in Sardinians. Eur. J. Hum. Genet. 2009, 17, 1448–1453. [Google Scholar] [CrossRef] [Green Version]
- Kaabi, Y.A.; Mansor, A.S.; Alfagih, A.S.; Hakami, A.M.; Summ, M.A.; Mjery, Y.A.; Alzughbi, M.N.; Habibullah, M.M. Frequency of UCP2 45-bp Ins/Del polymorphism in Saudi population from Jazan area and its association with autoimmune hypothyroidism UCP2 45-bp Ins/Del frequency in hypothyroidism. Int. J. Health Sci. 2020, 14, 11–16. [Google Scholar]


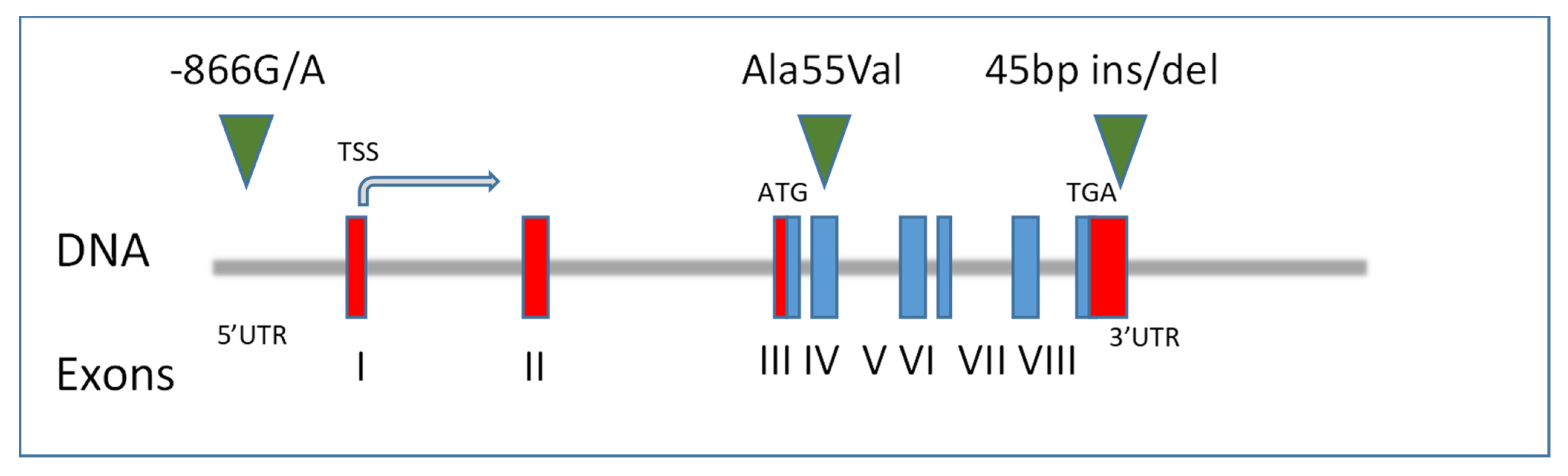{kind=link}
| Variables | Controls (N = 241) | Ischemic Strokes (N = 250) | Odds Ratio (95% CI) | p-Value |
|---|---|---|---|---|
| Median age in years a | 69.57 (SD13.9) | 74.06 (SD11.39) | 1.145 (2.24–6.73) | 0.0001 |
| Sex, Males | 138 (57.2%) | 149 (59.6%) | 1.10 (0.76–1.57) | 0.59 |
| History of smoking | 89 (36.9%) | 98 (39.2%) | 1.10 (0.76–1.8) | 0.6 |
| Alcohol | 32 (13.2%) | 30 (12.0%) | 0.89 (0.52–1.51) | 0.67 |
| Diabetes | 68 (28.2%) | 59 (23.6%) | 0.78 (0.52–1.17) | 0.24 |
| Hypercholesterolemia a | 29 (12.0%) | 58 (23.2%) | 2.20 (1.35–3.59) | 0.001 |
| Peripheral arteriopathy | 13 (5.4%) | 10 (4.0%) | 0.73 (0.31–1.69) | 0.46 |
| Hypertension a | 114 (47.3%) | 164 (65.6%) | 2.28 (1.58–3.28) | <0.0001 |
| History of MI | 34 (14.1%) | 38 (15.2%) | 1.09 (0.66–1.80) | 0.72 |
| Atrial fibrillationa | 37 (15.3%) | 68 (27.2%) | 2.06 (1.31–3.2) | 0.001 |
| Markers | CNT (N = 241) | Stroke (N = 250) | Odds Ratio (CI 95%) | p-Value |
|---|---|---|---|---|
| rs659366 | ||||
| (−866G/A) | ||||
| GG 114 (47, 3%) | GG 118 (47, 2%) | |||
| Genotype N (%) | GA 104 (43, 1%) | GA 110 (44%) | 0.80 | |
| AA 23 (9, 6%) | AA 22 (8, 8%) | 0.92 (0.48–1.75) | ||
| Allele N (%) | G 332 (68, 9%) | G 346 (69.2%) | ||
| A 150 (31.1%) | A 154 (30.8%) | 0.98 (0.752–1.291) | 0.91 | |
| rs1554995310 | ||||
| (45 bp ins/del) | ||||
| DD 134 (55, 6%) | DD 141 (56, 4%) | |||
| Genotype N (%) | DI 91 (37, 8%) | DI 97 (38, 8%) | 0.39 | |
| II 16 (6, 6%) | II 12 (4, 8%) | 0.71 (0.32–1.56) | ||
| Allele N (%) | D 359 (74.5%) | D 379 (78.6%) | ||
| I 123 (25.5%) | I 121 (21.4%) | 0.93 (0.698–1.245) | 0.93 |
| Variables | Coefficient | SE | p-Value | OR | CI 95% |
|---|---|---|---|---|---|
| rs659366 (−866G/A) | 0.0124 | 0.0376 | 0.7415 | 1.0125 | (0.9405–1.0900) |
| rs1554995310 (45 bp ins/del) | −0.049 | 0.0466 | 0.2936 | 0.9522 | (0.8691–1.0433) |
| Sex | 0.0211 | 0.2235 | 0.9247 | 1.0213 | (0.6591–1.5826) |
| Age * | 0.0233 | 0.0089 | 0.009 | 1.0236 | (1.0058–1.0416) |
| History of smoking | 0.8277 | 0.2963 | 0.0052 | 2.2879 | (1.2802–4.0891) |
| Alcohol | 0.4991 | 0.3553 | 0.1601 | 1.6473 | (0.8209–3.3054) |
| Diabetes | −0.3551 | 0.2267 | 0.1172 | 0.7011 | (0.4496–1.0933) |
| Hypercholesterolemia | 0.848 | 0.2708 | 0.0017 | 2.3351 | (1.3735–3.9698) |
| Peripheral arteriopathy | −0.2036 | 0.4755 | 0.6685 | 0.8158 | (0.3212–2.0715) |
| Hypertension | 0.6093 | 0.2031 | 0.0027 | 1.8391 | (1.2352–2.7382) |
| History of MI | −0.1287 | 0.2785 | 0.644 | 0.8792 | (0.5093–1.5177) |
| Atrial fibrillation | 0.5975 | 0.2468 | 0.0155 | 1.8176 | (1.1206–2.9481) |
| Constant | −1.8374 | 0.9315 | 0.0485 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanzione, R.; Cotugno, M.; Forte, M.; Bianchi, F.; Marchitti, S.; Palomba, N.P.; Esposito, T.; Zanda, B.; Sanna, A.; Rubattu, S. Role of Uncoupling Protein 2 Gene Polymorphisms on the Risk of Ischemic Stroke in a Sardinian Population. Life 2022, 12, 721. https://doi.org/10.3390/life12050721
Stanzione R, Cotugno M, Forte M, Bianchi F, Marchitti S, Palomba NP, Esposito T, Zanda B, Sanna A, Rubattu S. Role of Uncoupling Protein 2 Gene Polymorphisms on the Risk of Ischemic Stroke in a Sardinian Population. Life. 2022; 12(5):721. https://doi.org/10.3390/life12050721
Chicago/Turabian StyleStanzione, Rosita, Maria Cotugno, Maurizio Forte, Franca Bianchi, Simona Marchitti, Nicole Piera Palomba, Teresa Esposito, Bastianina Zanda, Alessandra Sanna, and Speranza Rubattu. 2022. "Role of Uncoupling Protein 2 Gene Polymorphisms on the Risk of Ischemic Stroke in a Sardinian Population" Life 12, no. 5: 721. https://doi.org/10.3390/life12050721