
Of Cockroaches and Symbionts: Recent Advances in the Characterization of the Relationship between Blattella germanica and Its Dual Symbiotic System


Abstract
:1. Introduction
1.1. The Impact of Symbiosis with Bacteria in Eukaryotic Evolution
1.2. Bacterial Symbiosis in Insects
1.3. Symbionts Must Be Present in Every Generation
2. The Dual Symbiotic System in Cockroaches
2.1. The Essential Role of Blattabacterium
2.2. Termites Have Lost Blattabacterium
2.3. The Gut Microbiota of Cockroaches and Its Not Fully Untangled Role
3. Why B. germanica has Preserved Both Symbiotic Systems?
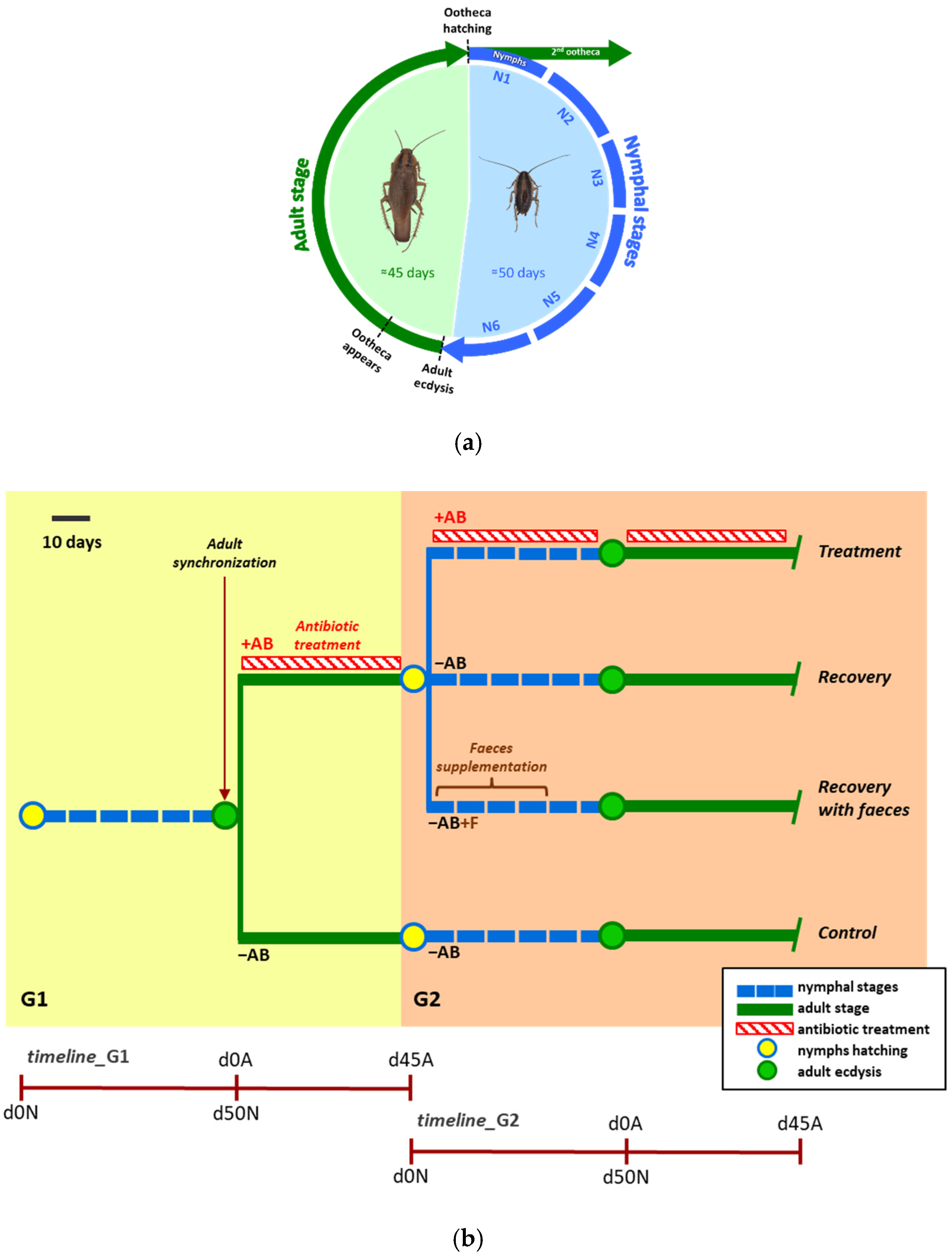
3.1. Perturbing the Gut Microbiota to Learn about It
3.2. The Resistome
3.3. Silencing Blattabacterium
4. Role of the Host Immune System in the Symbiotic Interaction
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Perreau, J.; Moran, N.A. Genetic innovations in animal–microbe symbioses. Nat. Rev. Genet. 2021, 23, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Baldo, L.; Werren, J.H. Evolutionary genetics of microbial symbiosis. Genes 2021, 12, 327. [Google Scholar] [CrossRef] [PubMed]
- Buchner, P. Endosymbiosis of Animals with Plant Microorganisms; Interscience Publishers: New York, NY, USA, 1965. [Google Scholar]
- Christian, N.; Whitaker, B.K.; Clay, K. Microbiomes: Unifying animal and plant systems through the lens of community ecology theory. Front. Microbiol. 2015, 6, 869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moya, A.; Pereto, J.; Gil, R.; Latorre, A. Learning how to live together: Genomic insights into prokaryote-animal symbioses. Nat. Rev. Genet. 2008, 9, 218–229. [Google Scholar] [CrossRef]
- Douglas, A.E. Multiorganismal insects: Diversity and function of resident microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K.; Engel, P. Mechanisms underlying gut microbiota–host interactions in insects. J. Exp. Biol. 2021, 224, jeb207696. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2020, 19, 55–71. [Google Scholar] [CrossRef]
- Sazama, E.J.; Ouellette, S.P.; Wesner, J.S. Bacterial endosymbionts are common among, but not necessarily within, insect species. Environ. Entomol. 2019, 48, 127–133. [Google Scholar] [CrossRef]
- Douglas, A.E. Mycetocyte symbiosis in insects. Biol. Rev. Camb. Philos. Soc. 1989, 64, 409–434. [Google Scholar] [CrossRef]
- Baumann, P. Biology bacteriocyte-associated endosymbionts of plant sap-sucking insects. Annu. Rev. Microbiol. 2005, 59, 155–189. [Google Scholar] [CrossRef]
- Latorre, A.; Manzano-Marín, A. Dissecting genome reduction and trait loss in insect endosymbionts. Ann. N. Y. Acad. Sci. 2017, 1389, 52–75. [Google Scholar] [CrossRef]
- Gil, R.; Latorre, A. Unity makes strength: A review on mutualistic symbiosis in representative insect clades. Life 2019, 9, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, R.; Silva, F.J.; Zientz, E.; Delmotte, F.; Gonzalez-Candelas, F.; Latorre, A.; Rausell, C.; Kamerbeek, J.; Gadau, J.; Holldobler, B.; et al. The genome sequence of Blochmannia floridanus: Comparative analysis of reduced genomes. Proc. Natl. Acad Sci. USA 2003, 100, 9388–9393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Sanchez, M.J.; Neef, A.; Peretó, J.; Patiño-Navarrete, R.; Pignatelli, M.; Latorre, A.; Moya, A. Evolutionary convergence and nitrogen metabolism in Blattabacterium strain Bge, primary endosymbiont of the cockroach Blattella germanica. PLoS Genet. 2009, 5, e1000721. [Google Scholar] [CrossRef] [Green Version]
- Patiño-Navarrete, R.; Piulachs, M.-D.; Belles, X.; Moya, A.; Latorre, A.; Peretó, J. The cockroach Blattella germanica obtains nitrogen from uric acid through a metabolic pathway shared with its bacterial endosymbiont. Biol. Lett. 2014, 10, 20140407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brune, A.; Dietrich, C. The gut microbiota of termites: Digesting the diversity in the light of ecology and evolution. Annu. Rev. Microbiol. 2015, 69, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Kundu, P.; Manna, B.; Majumder, S.; Ghosh, A. Species-wide metabolic interaction network for understanding natural lignocellulose digestion in termite gut microbiota. Sci. Rep. 2019, 9, 16329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikaelyan, A.; Meuser, K.; Brune, A. Microenvironmental heterogeneity of gut compartments drives bacterial community structure in wood- and humus-feeding higher termites. FEMS Microbiol. Ecol. 2017, 93, fiw210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, P.; Moran, N.A. The gut microbiota of insects—Diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef] [PubMed]
- Dillon, R.J.; Dillon, V.M. The gut bacteria of insects: Nonpathogenic interactions. Annu. Rev. Entomol. 2004, 49, 71–92. [Google Scholar] [CrossRef]
- Paniagua Voirol, L.R.; Frago, E.; Kaltenpoth, M.; Hilker, M.; Fatouros, N.E. Bacterial symbionts in lepidoptera: Their diversity, transmission, and impact on the host. Front. Microbiol. 2018, 9, 556. [Google Scholar] [CrossRef]
- Chen, B.; Xie, S.; Zhang, X.; Zhang, N.; Feng, H.; Sun, C.; Lu, X.; Shao, Y. Gut microbiota metabolic potential correlates with body size between mulberry-feeding lepidopteran pest species. Pest Manag. Sci. 2020, 76, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Broderick, N.A.; Lemaitre, B. Gut-associated microbes of Drosophila melanogaster. Gut Microbes 2012, 3, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Hurst, G.D.D. Extended genomes: Symbiosis and evolution. Interface Focus 2017, 7, 20170001. [Google Scholar] [CrossRef] [PubMed]
- Onchuru, T.O.; Javier Martinez, A.; Ingham, C.S.; Kaltenpoth, M. Transmission of mutualistic bacteria in social and gregarious insects. Curr. Opin. Insect Sci. 2018, 28, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Manzano-Marín, A.; Latorre, A. Snapshots of a shrinking partner: Genome reduction in Serratia symbiotica. Sci. Rep. 2016, 6, 32590. [Google Scholar] [CrossRef]
- Sacchi, L.; Grigolo, A.; Mazzini, M.; Bigliardi, E.; Baccetti, B.; Laudani, U. Symbionts in the oocytes of Blattella germanica (L.) (Dictyoptera : Blattellidae): Their mode of transmission. Int. J. Insect Morphol. Embryol. 1988, 17, 437–446. [Google Scholar] [CrossRef]
- Koga, R.; Meng, X.Y.; Tsuchida, T.; Fukatsu, T. Cellular mechanism for selective vertical transmission of an obligate insect symbiont at the bacteriocyte-embryo interface. Proc. Natl. Acad. Sci. USA 2012, 109, E1230–E1237. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [Green Version]
- Bright, M.; Bulgheresi, S. A complex journey: Transmission of microbial symbionts. Nat. Rev. Microbiol. 2010, 8, 218–230. [Google Scholar] [CrossRef] [Green Version]
- Nalepa, C.A. Origin of termite eusociality: Trophallaxis integrates the social, nutritional, and microbial environments. Ecol. Entomol. 2015, 40, 323–335. [Google Scholar] [CrossRef]
- Staudacher, H.; Kaltenpoth, M.; Breeuwer, J.A.J.; Menken, S.B.J.; Heckel, D.G.; Groot, A.T. Variability of bacterial communities in the moth Heliothis virescens indicates transient association with the host. PLoS ONE 2016, 11, e0154514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salem, H.; Florez, L.; Gerardo, N.; Kaltenpoth, M. An out-of-body experience: The extracellular dimension for the transmission of mutualistic bacteria in insects. Proc. Biol. Sci. 2015, 282, 20142957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakula, M. The persistence of a microbial flora during postembryogenesis of Drosophila melanogaster. J. Invertebr. Pathol. 1969, 14, 365–374. [Google Scholar] [CrossRef]
- Bourguignon, T.; Lo, N.; Dietrich, C.; Šobotník, J.; Sidek, S.; Roisin, Y.; Brune, A.; Evans, T.A. Rampant host switching shaped the termite gut microbiome. Curr. Biol. 2018, 28, 649–654.e2. [Google Scholar] [CrossRef] [Green Version]
- Rosas, T.; García-Ferris, C.; Domínguez-Santos, R.; Llop, P.; Latorre, A.; Moya, A. Rifampicin treatment of Blattella germanica evidences a fecal transmission route of their gut microbiota. FEMS Microbiol. Ecol. 2018, 94, fiy002. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Santos, R.; Pérez-Cobas, A.E.; Artacho, A.; Castro, J.A.; Talón, I.; Moya, A.; García-Ferris, C.; Latorre, A. Unraveling assemblage, functions and stability of the gut microbiota of Blattella germanica by antibiotic treatment. Front. Microbiol. 2020, 11, 487. [Google Scholar] [CrossRef]
- Domínguez-Santos, R.; Pérez-Cobas, A.E.; Cuti, P.; Pérez-Brocal, V.; García-Ferris, C.; Moya, A.; Latorre, A.; Gil, R. Interkingdom gut microbiome and resistome of the cockroach Blattella germanica. mSystems 2021, 6, e01213-20. [Google Scholar] [CrossRef]
- Kopanic, R.J.; Holbrook, G.L.; Sevala, V.; Schal, C. An adaptive benefit of facultative coprophagy in the German cockroach Blattella germanica. Ecol. Entomol. 2001, 26, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Jahnes, B.C.; Herrmann, M.; Sabree, Z.L. Conspecific coprophagy stimulates normal development in a germ-free model invertebrate. PeerJ 2019, 7, e6914. [Google Scholar] [CrossRef]
- Renelies-Hamilton, J.; Germer, K.; Sillam-Dussès, D.; Bodawatta, K.H.; Poulsen, M. Disentangling the relative roles of vertical transmission, subsequent colonizations, and diet on cockroach microbiome assembly. mSphere 2021, 6, e01023-20. [Google Scholar] [CrossRef] [PubMed]
- Gosalbes, M.J.; Llop, S.; Vallès, Y.; Moya, A.; Ballester, F.; Francino, M.P. Meconium microbiota types dominated by lactic acid or enteric bacteria are differentially associated with maternal eczema and respiratory problems in infants. Clin. Exp. Allergy 2013, 43, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Price, J.; Mahurkar, A.; Rahnavard, G.; Crabtree, J.; Orvis, J.; Hall, A.B.; Brady, A.; Creasy, H.H.; McCracken, C.; Giglio, M.G.; et al. Strains, functions and dynamics in the expanded Human Microbiome Project. Nature 2017, 550, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Martí, J.M.; Martínez-Martínez, D.; Rubio, T.; Gracia, C.; Peña, M.; Latorre, A.; Moya, A.; Garay, C. Health and disease imprinted in the time variability of the human microbiome. mSystems 2017, 2, e00144-16. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.J.; Rodríguez-Ruano, S.M.; Poosakkannu, A.; Batani, G.; Schmidt, J.O.; Roachell, W.; Zima, J.; Hypša, V.; Nováková, E. Ontogeny, species identity, and environment dominate microbiome dynamics in wild populations of kissing bugs (Triatominae). Microbiome 2020, 8, 146. [Google Scholar] [CrossRef]
- Wang, Y.; Rozen, D.E. Gut microbiota colonization and transmission in the burying beetle Nicrophorus vespilloides throughout development. Appl. Environ. Microbiol. 2017, 83, e03250-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourguignon, T.; Kinjo, Y.; Villa-Martín, P.; Coleman, N.V.; Tang, Q.; Arab, D.A.; Wang, Z.; Tokuda, G.; Hongoh, Y.; Ohkuma, M.; et al. Increased mutation rate is linked to genome reduction in prokaryotes. Curr. Biol. 2020, 30, 3848–3855.e4. [Google Scholar] [CrossRef]
- Kinjo, Y.; Lo, N.; Martín, P.V.; Tokuda, G.; Pigolotti, S.; Bourguignon, T. Enhanced mutation rate, relaxed selection, and the “domino effect” are associated with gene loss in Blattabacterium, a cockroach endosymbiont. Mol. Biol. Evol. 2021, 38, 3820–3831. [Google Scholar] [CrossRef]
- Vicente, C.S.L.; Mondal, S.I.; Akter, A.; Ozawa, S.; Kikuchi, T.; Hasegawa, K. Genome analysis of new Blattabacterium spp., obligatory endosymbionts of Periplaneta fuliginosa and P. japonica. PLoS ONE 2018, 13, e0200512. [Google Scholar] [CrossRef]
- Jennings, E.C.; Korthauer, M.W.; Benoit, J.B. Genome sequence of a Blattabacterium strain isolated from the viviparous cockroach, Diploptera punctata. Microbiol. Resour. Announc. 2020, 9, e00229-20. [Google Scholar] [CrossRef] [PubMed]
- Tinker, K.A.; Ottesen, E.A. Phylosymbiosis across deeply diverging lineages of omnivorous cockroaches (order Blattodea). Appl. Environ. Microbiol. 2020, 86, e02513-19. [Google Scholar] [CrossRef] [PubMed]
- Lampert, N.; Mikaelyan, A.; Brune, A. Diet is not the primary driver of bacterial community structure in the gut of litter-feeding cockroaches. BMC Microbiol. 2019, 19, 238. [Google Scholar] [CrossRef] [PubMed]
- Jennings, E.C.; Korthauer, M.W.; Hamilton, T.L.; Benoit, J.B. Matrotrophic viviparity constrains microbiome acquisition during gestation in a live-bearing cockroach, Diploptera punctata. Ecol. Evol. 2019, 9, 10601–10614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, C.; Köhler, T.; Brune, A. The Cockroach origin of the termite gut microbiota: Patterns in bacterial community structure reflect major evolutionary events. Appl. Environ. Microbiol. 2014, 80, 2261–2269. [Google Scholar] [CrossRef] [Green Version]
- Gontang, E.A.; Aylward, F.O.; Carlos, C.; del Rio, T.G.; Chovatia, M.; Fern, A.; Lo, C.-C.; Malfatti, S.A.; Tringe, S.G.; Currie, C.R.; et al. Major changes in microbial diversity and community composition across gut sections of a juvenile Panchlora cockroach. PLoS ONE 2017, 12, e0177189. [Google Scholar] [CrossRef]
- Bauer, E.; Lampert, N.; Mikaelyan, A.; Köhler, T.; Maekawa, K.; Brune, A. Physicochemical conditions, metabolites and community structure of the bacterial microbiota in the gut of wood-feeding cockroaches (Blaberidae: Panesthiinae). FEMS Microbiol. Ecol. 2015, 91, fiu028. [Google Scholar] [CrossRef] [Green Version]
- Tinker, K.A.; Ottesen, E.A. The core gut microbiome of the American cockroach, Periplaneta americana, is stable and resilient to dietary shifts. Appl. Environ. Microbiol. 2016, 82, 6603–6610. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kim, J.Y.; Yi, M.; Lee, I.-Y.; Lee, W.-J.; Moon, H.S.; Yong, D.; Yong, T.-S. Comparative microbiome analysis of three species of laboratory-reared Periplaneta cockroaches. Korean J. Parasitol. 2020, 58, 537–542. [Google Scholar] [CrossRef]
- Tinker, K.A.; Ottesen, E.A. Differences in gut microbiome composition between sympatric wild and allopatric laboratory populations of omnivorous cockroaches. Front. Microbiol. 2021, 12, 703785. [Google Scholar] [CrossRef]
- Schauer, C.; Thompson, C.L.; Brune, A. The bacterial community in the gut of the cockroach Shelfordella lateralis reflects the close evolutionary relatedness of cockroaches and termites. Appl. Environ. Microbiol. 2012, 78, 2758–2767. [Google Scholar] [CrossRef] [Green Version]
- Schauer, C.; Thompson, C.; Brune, A. Pyrotag sequencing of the gut microbiota of the cockroach Shelfordella lateralis reveals a highly dynamic core but only limited effects of diet on community structure. PLoS ONE 2014, 9, e85861. [Google Scholar] [CrossRef] [PubMed]
- Berlanga, M.; Llorens, C.; Comas, J.; Guerrero, R. Gut bacterial community of the xylophagous cockroaches Cryptocercus punctulatus and Parasphaeria boleiriana. PLoS ONE 2016, 11, e0152400. [Google Scholar] [CrossRef] [Green Version]
- Carrasco, P.; Pérez-Cobas, A.E.; Van De Pol, C.; Baixeras, J.; Moya, A.; Latorre, A. Succession of the gut microbiota in the cockroach Blattella germanica. Int. Microbiol. 2014, 17, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cobas, A.E.; Maiques, E.; Angelova, A.; Carrasco, P.; Moya, A.; Latorre, A. Diet shapes the gut microbiota of the omnivorous cockroach Blattella germanica. FEMS Microbiol. Ecol. 2015, 91, fiv022. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Yang, R. Life history and functional capacity of the microbiome are altered in beta-cypermethrin-resistant cockroaches. Int. J. Parasitol. 2019, 49, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Sabree, Z.L.; Huang, C.Y.; Arakawa, G.; Tokuda, G.; Lo, N.; Watanabe, H.; Moran, N.A. Genome shrinkage and loss of nutrient-providing potential in the obligate symbiont of the primitive termite Mastotermes darwiniensis. Appl. Environ. Microbiol. 2012, 78, 204–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.S.; Park, P.; Takeda, M. Roles of fat body trophocytes, mycetocytes and urocytes in the American cockroach, Periplaneta americana under starvation conditions: An ultrastructural study. Arthropod Struct. Dev. 2013, 42, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Patiño-Navarrete, R.; Moya, A.; Latorre, A.; Peretó, J. Comparative genomics of Blattabacterium cuenoti: The frozen legacy of an ancient endosymbiont genome. Genome Biol. Evol. 2013, 5, 351–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabree, Z.L.; Kambhampati, S.; Moran, N.A. Nitrogen recycling and nutritional provisioning by Blattabacterium, the cockroach endosymbiont. Proc. Natl. Acad. Sci. USA 2009, 106, 19521–19526. [Google Scholar] [CrossRef] [Green Version]
- González-Domenech, C.; Belda, E.; Patiño-Navarrete, R.; Moya, A.; Peretó, J.; Latorre, A. Metabolic stasis in an ancient symbiosis: Genome-scale metabolic networks from two Blattabacterium cuenoti strains, primary endosymbionts of cockroaches. BMC Microbiol. 2012, 12 (Suppl. 1), S5. [Google Scholar] [CrossRef] [Green Version]
- Inward, D.; Beccaloni, G.; Eggleton, P. Death of an order: A comprehensive molecular phylogenetic study confirms that termites are eusocial cockroaches. Biol. Lett. 2007, 3, 331–335. [Google Scholar] [CrossRef]
- Bourguignon, T.; Lo, N.; Cameron, S.L.; Šobotník, J.; Hayashi, Y.; Shigenobu, S.; Watanabe, D.; Roisin, Y.; Miura, T.; Evans, T.A. The evolutionary history of termites as inferred from 66 mitochondrial genomes. Mol. Biol. Evol. 2015, 32, 406–421. [Google Scholar] [CrossRef] [Green Version]
- Neef, A.; Latorre, A.; Peretó, J.; Silva, F.J.; Pignatelli, M.; Moya, A. Genome economization in the endosymbiont of the wood roach Cryptocercus punctulatus due to drastic loss of amino acid synthesis capabilities. Genome Biol. Evol. 2011, 3, 1437–1448. [Google Scholar] [CrossRef] [Green Version]
- Lo, N.; Tokuda, G.; Watanabe, H.; Rose, H.; Slaytor, M.; Maekawa, K.; Bandi, C.; Noda, H. Evidence from multiple gene sequences indicates that termites evolved from wood-feeding cockroaches. Curr. Biol. 2000, 10, 801–804. [Google Scholar] [CrossRef] [Green Version]
- Bandi, C.; Sironi, M.; Damiani, G.; Magrassi, L.; Nalepa, C.A.; Laudani, U.; Sacchi, L. The establishment of intracellular symbiosis in an ancestor of cockroaches and termites. Proc. R. Soc. Lond. B Biol. Sci. 1995, 259, 293–299. [Google Scholar] [CrossRef]
- Lo, N.; Bandi, C.; Watanabe, H.; Nalepa, C.; Beninati, T. Evidence for cocladogenesis between diverse dictyopteran lineages and their intracellular endosymbionts. Mol. Biol. Evol. 2003, 20, 907–913. [Google Scholar] [CrossRef] [Green Version]
- Kinjo, Y.; Bourguignon, T.; Tong, K.J.; Kuwahara, H.; Lim, S.J.; Yoon, K.B.; Shigenobu, S.; Park, Y.C.; Nalepa, C.A.; Hongoh, Y.; et al. Parallel and gradual genome erosion in the Blattabacterium endosymbionts of Mastotermes darwiniensis and Cryptocercus wood roaches. Genome Biol. Evol. 2018, 10, 1622–1630. [Google Scholar] [CrossRef] [Green Version]
- Chouvenc, T.; Šobotník, J.; Engel, M.S.; Bourguignon, T. Termite evolution: Mutualistic associations, key innovations, and the rise of Termitidae. Cell. Mol. Life Sci. 2021, 78, 2749–2769. [Google Scholar] [CrossRef]
- Nalepa, C.A. Origin of mutualism between termites and flagellated gut protists: Transition from horizontal to vertical transmission. Front. Ecol. Evol. 2020, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Tokuda, G.; Elbourne, L.D.H.H.; Kinjo, Y.; Saitoh, S.; Sabree, Z.; Hojo, M.; Yamada, A.; Hayashi, Y.; Shigenobu, S.; Bandi, C.; et al. Maintenance of essential amino acid synthesis pathways in the Blattabacterium cuenoti symbiont of a wood-feeding cockroach. Biol. Lett. 2013, 9, 20121153. [Google Scholar] [CrossRef] [Green Version]
- Hervé, V.; Liu, P.; Dietrich, C.; Sillam-Dussès, D.; Stiblik, P.; Šobotník, J.; Brune, A. Phylogenomic analysis of 589 metagenome-assembled genomes encompassing all major prokaryotic lineages from the gut of higher termites. PeerJ. 2020, 8, e8614. [Google Scholar] [CrossRef] [Green Version]
- Marynowska, M.; Goux, X.; Sillam-Dussès, D.; Rouland-Lefèvre, C.; Halder, R.; Wilmes, P.; Gawron, P.; Roisin, Y.; Delfosse, P.; Calusinska, M. Compositional and functional characterisation of biomass-degrading microbial communities in guts of plant fibre- and soil-feeding higher termites. Microbiome 2020, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- Wada-Katsumata, A.; Zurek, L.; Nalyanya, G.; Roelofs, W.L.; Zhang, A.; Schal, C. Gut bacteria mediate aggregation in the German cockroach. Proc. Natl. Acad. Sci. USA 2015, 112, 201504031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, R.; Potts, R.; Pietri, J.E. The persistence of Escherichia coli infection in German cockroaches (Blattodea: Blattellidae) varies between host developmental stages and is influenced by the gut microbiota. J. Med. Entomol. 2020, 57, 1964–1971. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of Prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef]
- Kakumanu, M.L.; Maritz, J.M.; Carlton, J.M.; Schal, C. Overlapping community compositions of gut and fecal microbiomes in lab-reared and field-collected German cockroaches. Appl. Environ. Microbiol. 2018, 84, e01037-18. [Google Scholar] [CrossRef] [Green Version]
- Tegtmeier, D.; Thompson, C.L.; Schauer, C.; Brune, A. Oxygen affects gut bacterial colonization and metabolic activities in a gnotobiotic cockroach model. Appl. Environ. Microbiol. 2016, 82, 1080–1089. [Google Scholar] [CrossRef] [Green Version]
- Mikaelyan, A.; Thompson, C.L.; Hofer, M.J.; Brune, A. Deterministic assembly of complex bacterial communities in guts of germ-free cockroaches. Appl. Environ. Microbiol. 2016, 82, 1256–1263. [Google Scholar] [CrossRef] [Green Version]
- Vera-Ponce de León, A.; Jahnes, B.C.; Otero-Bravo, A.; Sabree, Z.L. Microbiota perturbation or elimination can inhibit normal development and elicit a starvation-like response in an omnivorous model invertebrate. mSystems 2021, 6, e00802-21. [Google Scholar] [CrossRef]
- D’Costa, V.M.; McGrann, K.M.; Hughes, D.W.; Wright, G.D. Sampling the antibiotic resistome. Science 2006, 311, 374–377. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.D.; Wright, G.D. Antibacterial drug discovery in the resistance era. Nature 2016, 529, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Cha, C.J. Antibiotic resistome from the One-Health perspective: Understanding and controlling antimicrobial resistance transmission. Exp. Mol. Med. 2021, 53, 301–309. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, Y.; Gao, G.F. Combining metagenomics and metatranscriptomics to study human, animal and environmental resistomes. Med. Microecol. 2020, 3, 100014. [Google Scholar] [CrossRef]
- Zhuang, M.; Achmon, Y.; Cao, Y.; Liang, X.; Chen, L.; Wang, H.; Siame, B.A.; Leung, K.Y. Distribution of antibiotic resistance genes in the environment. Environ. Pollut. 2021, 285, 117402. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, D.; Walsh, F. Antibiotic resistance genes across a wide variety of metagenomes. FEMS Microbiol. Ecol. 2016, 92, fiv168. [Google Scholar] [CrossRef]
- Hyde, J.; Gorham, C.; Brackney, D.E.; Steven, B. Antibiotic resistant bacteria and commensal fungi are common and conserved in the mosquito microbiome. PLoS ONE 2019, 14, e0218907. [Google Scholar] [CrossRef] [Green Version]
- Akinjogunla, O.J.; Odeyemib, A.T.; Udoinyangc, E.P. Cockroaches (Periplaneta americana and Blattella germanica): Reservoirs of multi drug resistant (MDR) bacteria in Uyo, Akwa Ibom State. Sci. J. Biol. Sci. 2012, 1, 19–30. [Google Scholar] [CrossRef]
- Pai, H.-H. Multidrug resistant bacteria isolated from cockroaches in long-term care facilities and nursing homes. Acta Trop. 2013, 125, 18–22. [Google Scholar] [CrossRef]
- Pérez-Cobas, A.E.; Moya, A.; Gosalbes, M.J.; Latorre, A. Colonization resistance of the gut microbiota against Clostridium difficile. Antibiotics 2015, 4, 337–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langdon, A.; Crook, N.; Dantas, G. The effects of antibiotics on the microbiome throughout development and alternative approaches for therapeutic modulation. Genome Med. 2016, 8, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babakhani, S.; Oloomi, M. Transposons: The agents of antibiotic resistance in bacteria. J. Basic Microbiol. 2018, 58, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Henry, S.M. The significance of microorganisms in the nutrition of insects. Trans. N. Y. Acad. Sci. 1962, 24, 676–683. [Google Scholar] [CrossRef]
- Brooks, M.A. Comments on the classification of intracellular symbiotes of cockroaches and a description of the species. J. Invertebr. Pathol. 1970, 16, 249–258. [Google Scholar] [CrossRef]
- Muñoz-Benavent, M.; Latorre, A.; Alemany-Cosme, E.; Marín-Miret, J.; Domínguez-Santos, R.; Silva, F.J.; Gil, R.; García-Ferris, C. Gut microbiota cannot compensate the impact of (quasi)aposymbiosis in Blattella germanica. Biology 2021, 10, 1013. [Google Scholar] [CrossRef]
- Marra, A.; Hanson, M.A.; Kondo, S.; Erkosar, B.; Lemaitre, B. Drosophila antimicrobial peptides and lysozymes regulate gut microbiota composition and abundance. mBio 2021, 12, e0082421. [Google Scholar] [CrossRef]
- Maire, J.; Vincent-Monégat, C.; Masson, F.; Zaidman-Rémy, A.; Heddi, A. An IMD-like pathway mediates both endosymbiont control and host immunity in the cereal weevil Sitophilus spp. Microbiome 2018, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Mergaert, P. Role of antimicrobial peptides in controlling symbiotic bacterial populations. Nat. Prod. Rep. 2018, 35, 336–356. [Google Scholar] [CrossRef]
- Bosch, T.C.G. Cnidarian-microbe interactions and the origin of innate immunity in metazoans. Annu. Rev. Microbiol. 2013, 67, 499–518. [Google Scholar] [CrossRef] [Green Version]
- Van De Velde, W.; Zehirov, G.; Szatmari, A.; Debreczeny, M.; Ishihara, H.; Kevei, Z.; Farkas, A.; Mikulass, K.; Nagy, A.; Tiricz, H.; et al. Plant peptides govern terminal differentiation of bacteria in symbiosis. Science 2010, 327, 1122–1126. [Google Scholar] [CrossRef] [PubMed]
- Login, F.H.; Balmand, S.; Vallier, A.; Vincent-Monégat, C.; Vigneron, A.; Weiss-Gayet, M.; Rochat, D.; Heddi, A. Antimicrobial peptides keep insect endosymbionts under control. Science 2011, 334, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Mergaert, P.; Kikuchi, Y.; Shigenobu, S.; Nowack, E.C.M. Metabolic integration of bacterial endosymbionts through antimicrobial peptides. Trends Microbiol. 2017, 25, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Carro, L.; Pujic, P.; Alloisio, N.; Fournier, P.; Boubakri, H.; Hay, A.E.; Poly, F.; François, P.; Hocher, V.; Mergaert, P.; et al. Alnus peptides modify membrane porosity and induce the release of nitrogen-rich metabolites from nitrogen-fixing Frankia. ISME J. 2015, 9, 1723–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masson, F.; Zaidman-Rémy, A.; Heddi, A. Antimicrobial peptides and cell processes tracking endosymbiont dynamics. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150298. [Google Scholar] [CrossRef] [Green Version]
- Harrison, M.C.; Jongepier, E.; Robertson, H.M.; Arning, N.; Bitard-Feildel, T.; Chao, H.; Childers, C.P.; Dinh, H.; Doddapaneni, H.; Dugan, S.; et al. Hemimetabolous genomes reveal molecular basis of termite eusociality. Nat. Ecol. Evol. 2018, 2, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhu, S.; Jia, Q.; Yuan, D.; Ren, C.; Li, K.; Liu, S.; Cui, Y.; Zhao, H.; Cao, Y.; et al. The genomic and functional landscapes of developmental plasticity in the American cockroach. Nat. Commun. 2018, 9, 1008. [Google Scholar] [CrossRef]
- Silva, F.J.; Muñoz-Benavent, M.; García-Ferris, C.; Latorre, A. Blattella germanica displays a large arsenal of antimicrobial peptide genes. Sci. Rep. 2020, 10, 21058. [Google Scholar] [CrossRef]
- Shigenobu, S.; Stern, D.L. Aphids evolved novel secreted proteins for symbiosis with bacterial endosymbiont. Proc. Biol. Sci. 2013, 280, 20121952. [Google Scholar] [CrossRef] [Green Version]
- Uchi, N.; Fukudome, M.; Nozaki, N.; Suzuki, M.; Osuki, K.I.; Shigenobu, S.; Uchiumi, T. Antimicrobial activities of cysteine-rich peptides specific to bacteriocytes of the pea aphid Acyrthosiphon pisum. Microbes Environ. 2019, 34, 155. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; Gupta, V.K.; Jiang, Y.; Yang, B.; Gong, L.; Zhu, H. Cross-kingdom small RNAs among animals, plants and microbes. Cells 2019, 8, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriano-Gutierrez, S.; Bongrand, C.; Essock-Burns, T.; Wu, L.; McFall-Ngai, M.J.; Ruby, E.G. The noncoding small RNA SsrA is released by Vibrio fischeri and modulates critical host responses. PLoS Biol. 2020, 18, e3000934. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Host Scientific Name | Family | Reference |
|---|---|---|
| Blaberus craniifer | Blaberidae | [52] |
| Byrsotria rothi | Blaberidae | [53] |
| Diploptera punctata | Blaberidae | [52,54,55] |
| Elliptorhina chopardi | Blaberidae | [55] |
| Eublaberus posticus | Blaberidae | [55] |
| Gromphadorhina portentosa | Blaberidae | [52] |
| Lucihormetica verrucosa | Blaberidae | [52] |
| Nauphoeta cinerea | Blaberidae | [52] |
| Opisthoplatia orientalis | Blaberidae | [55] |
| Oxyhaloa deusta | Blaberidae | [52] |
| Panchlora sp. | Blaberidae | [56] |
| Panchlora viridis | Blaberidae | [52] |
| Panesthia angustipennis | Blaberidae | [55,57] |
| Pycnoscelus surinamensis | Blaberidae | [52] |
| Salganea esakii | Blaberidae | [55] |
| Schultesia lampyridiformis | Blaberidae | [52,55] |
| Blatta orientalis | Blattidae | [55] |
| Eurycotis floridana | Blattidae | [55] |
| Periplaneta americana | Blattidae | [52,58,59,60] |
| Periplaneta fuliginosa | Blattidae | [52,59,60] |
| Periplaneta japonica | Blattidae | [59] |
| Shelfordella lateralis | Blattidae | [42,61,62] |
| Ergaula pilosa | Corydiidae | [52] |
| Ergaula capucina | Corydiidae | [52,55] |
| Polyphaga aegyptiaca | Corydiidae | [52] |
| Therea olegrandjeani | Corydiidae | [52] |
| Cryptocercus punctulatus | Cryptocercidae | [55,63] |
| Blattella germanica | Ectobiidae | [37], [38,39], [64,65], [66] |
| Parcoblatta fulvescens | Ectobiidae | [52] |
| Symploce macroptera | Ectobiidae | [55] |
| Symploce pallens | Ectobiidae | [52] |
| Rhyparobia maderae | Nauphoetidae | [55] |
| Mastotermes darwiniensis | Mastotermitidae | [55,67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latorre, A.; Domínguez-Santos, R.; García-Ferris, C.; Gil, R. Of Cockroaches and Symbionts: Recent Advances in the Characterization of the Relationship between Blattella germanica and Its Dual Symbiotic System. Life 2022, 12, 290. https://doi.org/10.3390/life12020290
Latorre A, Domínguez-Santos R, García-Ferris C, Gil R. Of Cockroaches and Symbionts: Recent Advances in the Characterization of the Relationship between Blattella germanica and Its Dual Symbiotic System. Life. 2022; 12(2):290. https://doi.org/10.3390/life12020290
Chicago/Turabian StyleLatorre, Amparo, Rebeca Domínguez-Santos, Carlos García-Ferris, and Rosario Gil. 2022. "Of Cockroaches and Symbionts: Recent Advances in the Characterization of the Relationship between Blattella germanica and Its Dual Symbiotic System" Life 12, no. 2: 290. https://doi.org/10.3390/life12020290